LOXL3 Function Beyond Amino Oxidase and Role in Pathologies, Including Cancer


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
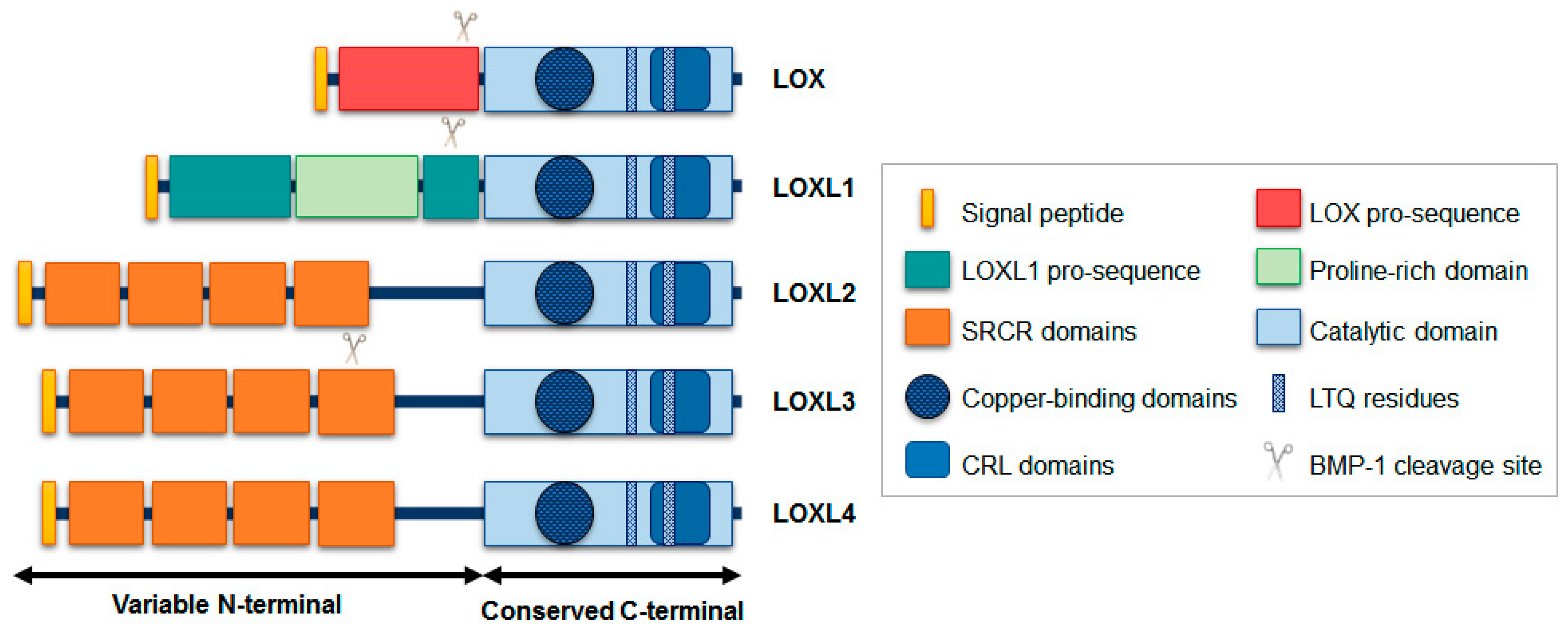
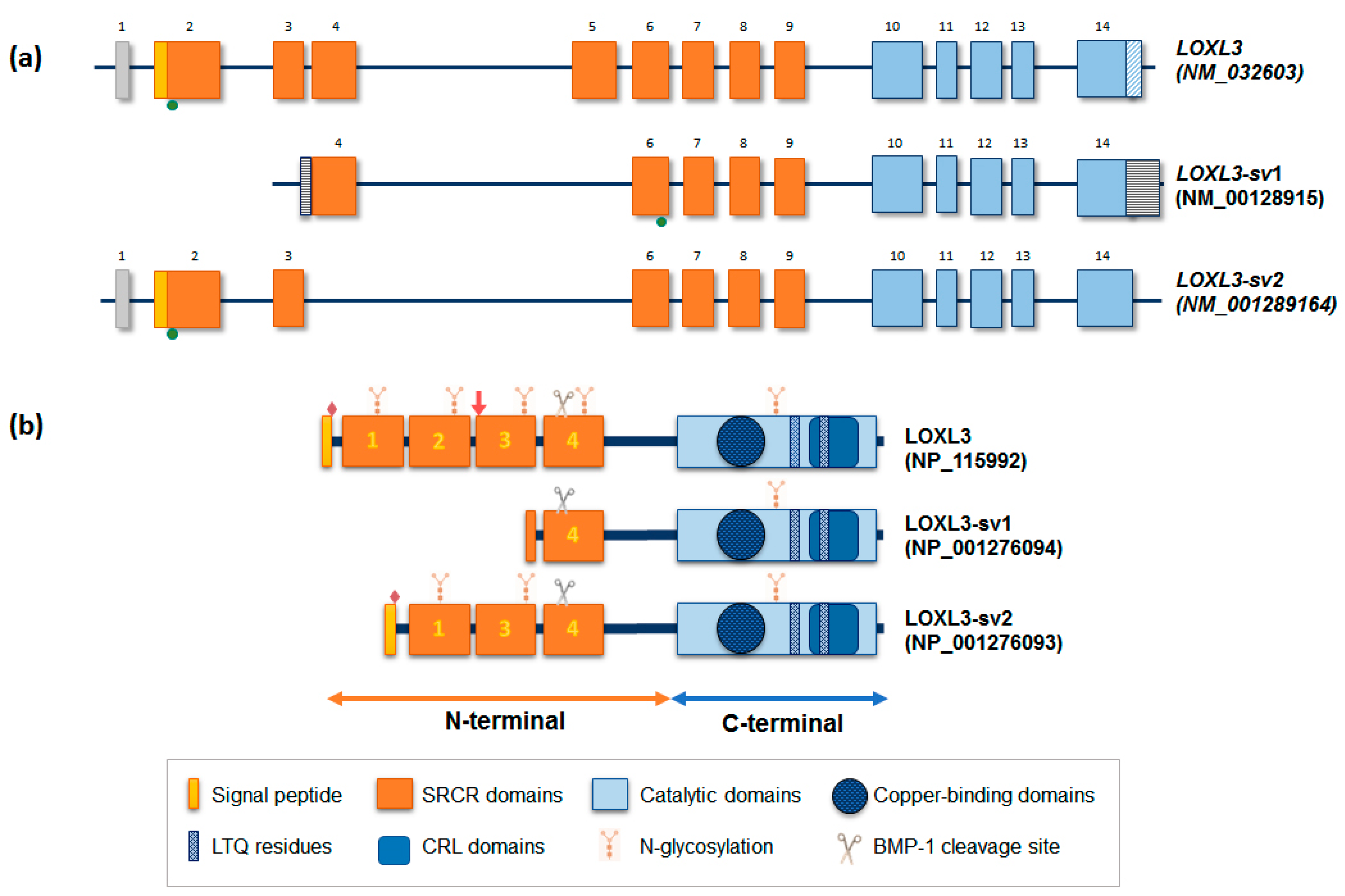
2. LOXL3: Gene and Protein Structure
3. Localization
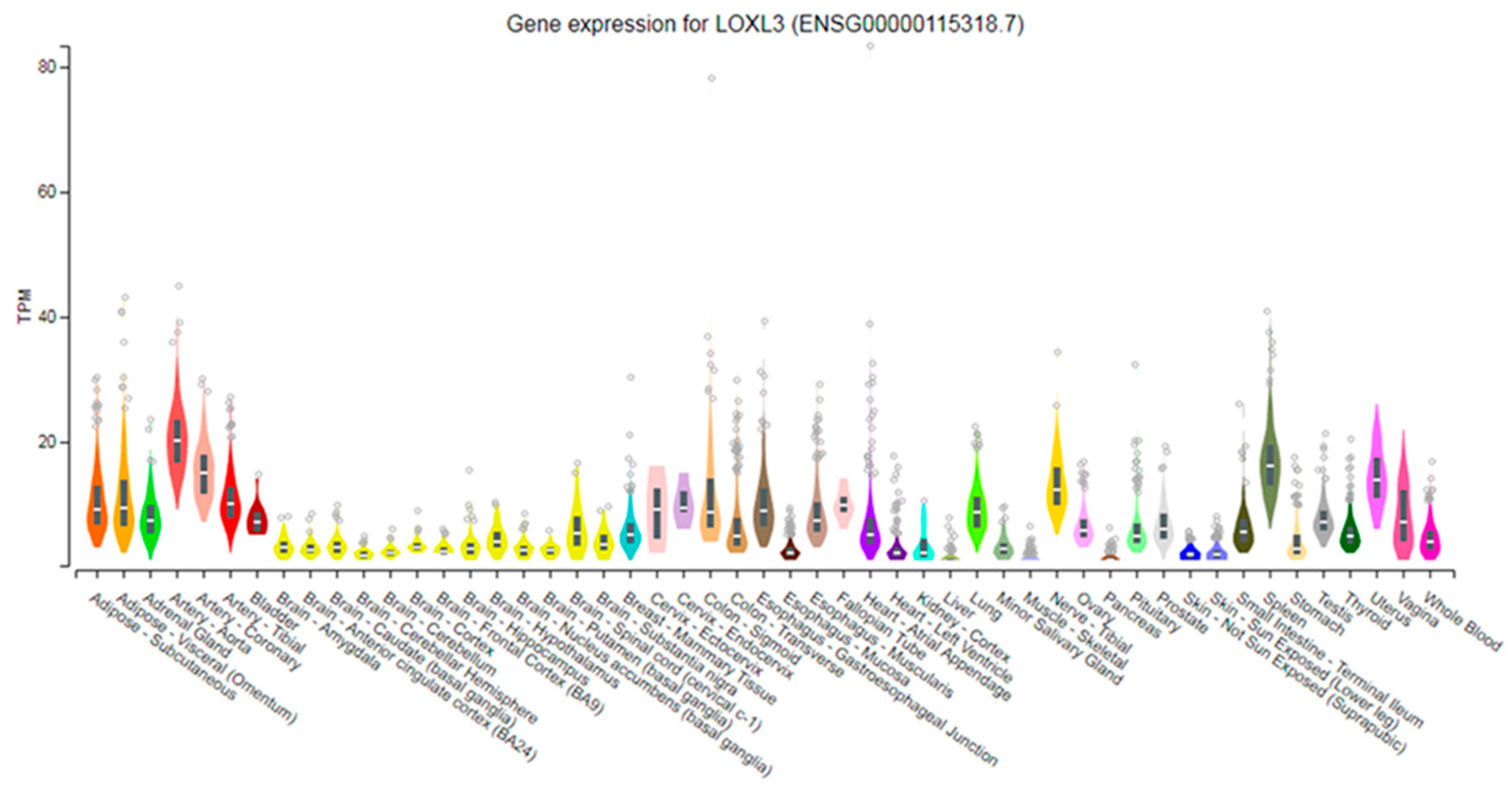
3.1. Tissue-Specific Expression of LOXL3
3.2. Subcellular Localization
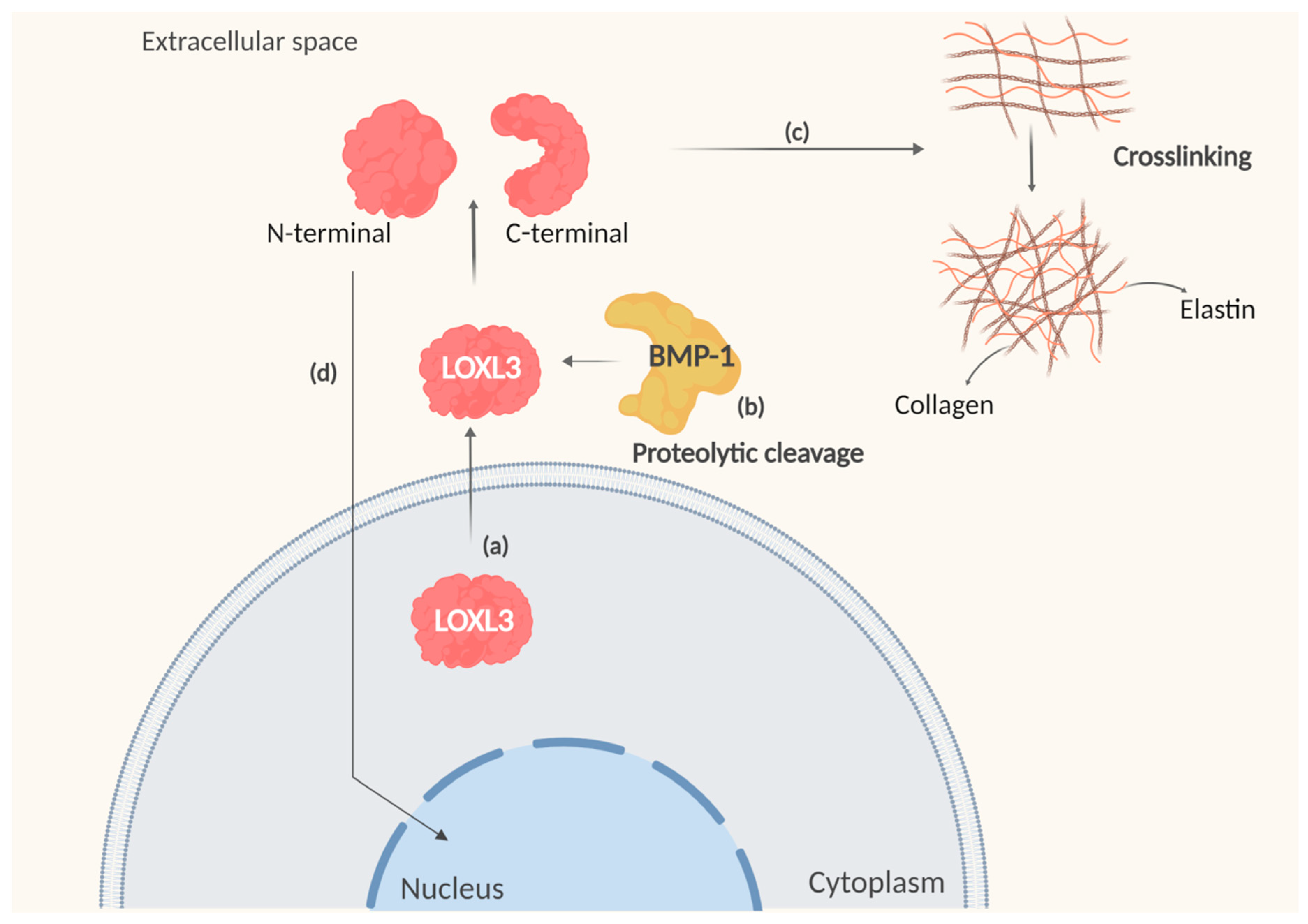
3.3. Amine Oxidase Activity of LOXL3
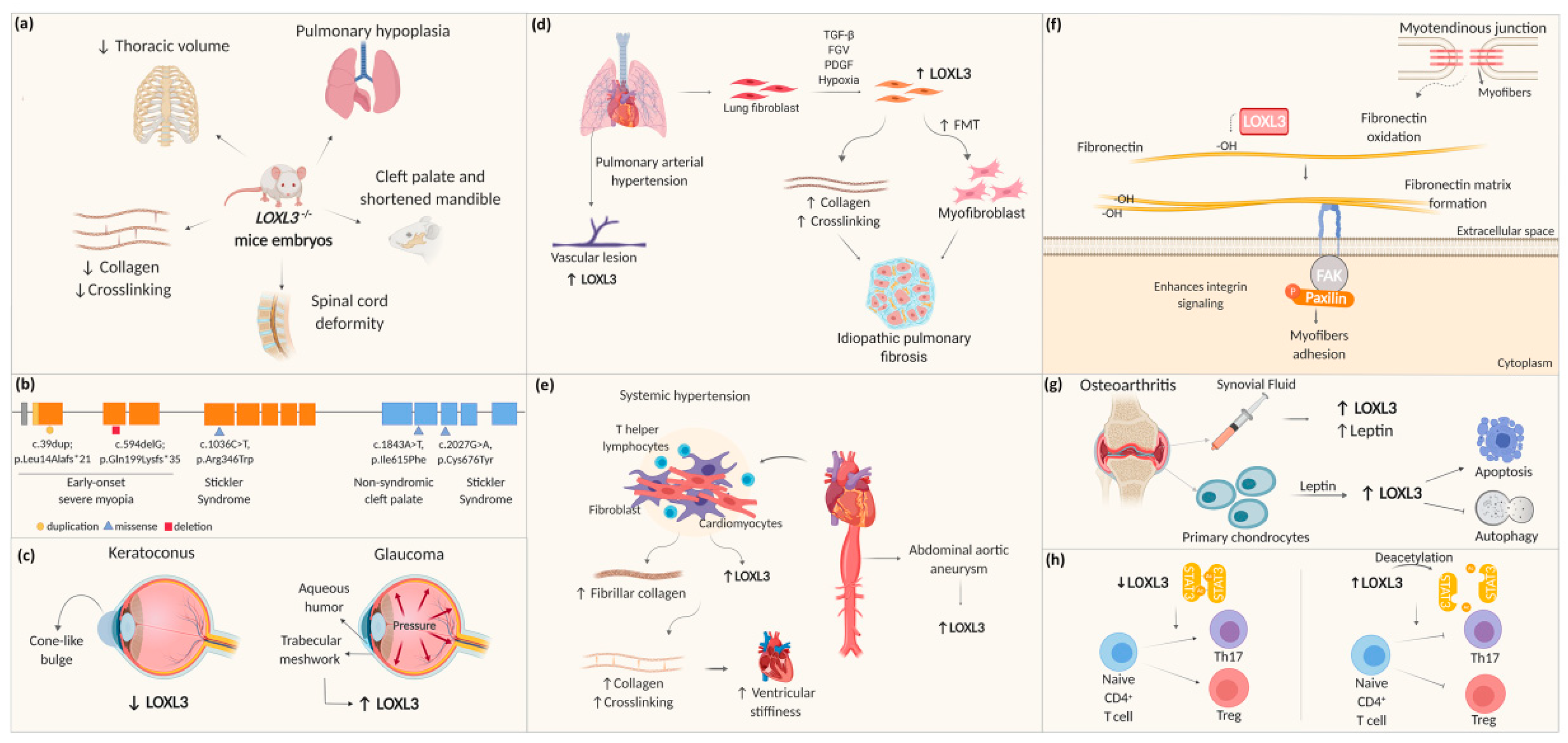
4. LOXL3 Roles in Development and Diseases
4.1. LOXL3 in Craniofacial-Ocular System
4.2. LOXL3 in Pulmonary and Cardiovascular Systems
4.3. LOXL3 in Myotendinous and Osteoarticular System
4.4. LOXL3 and Immune System
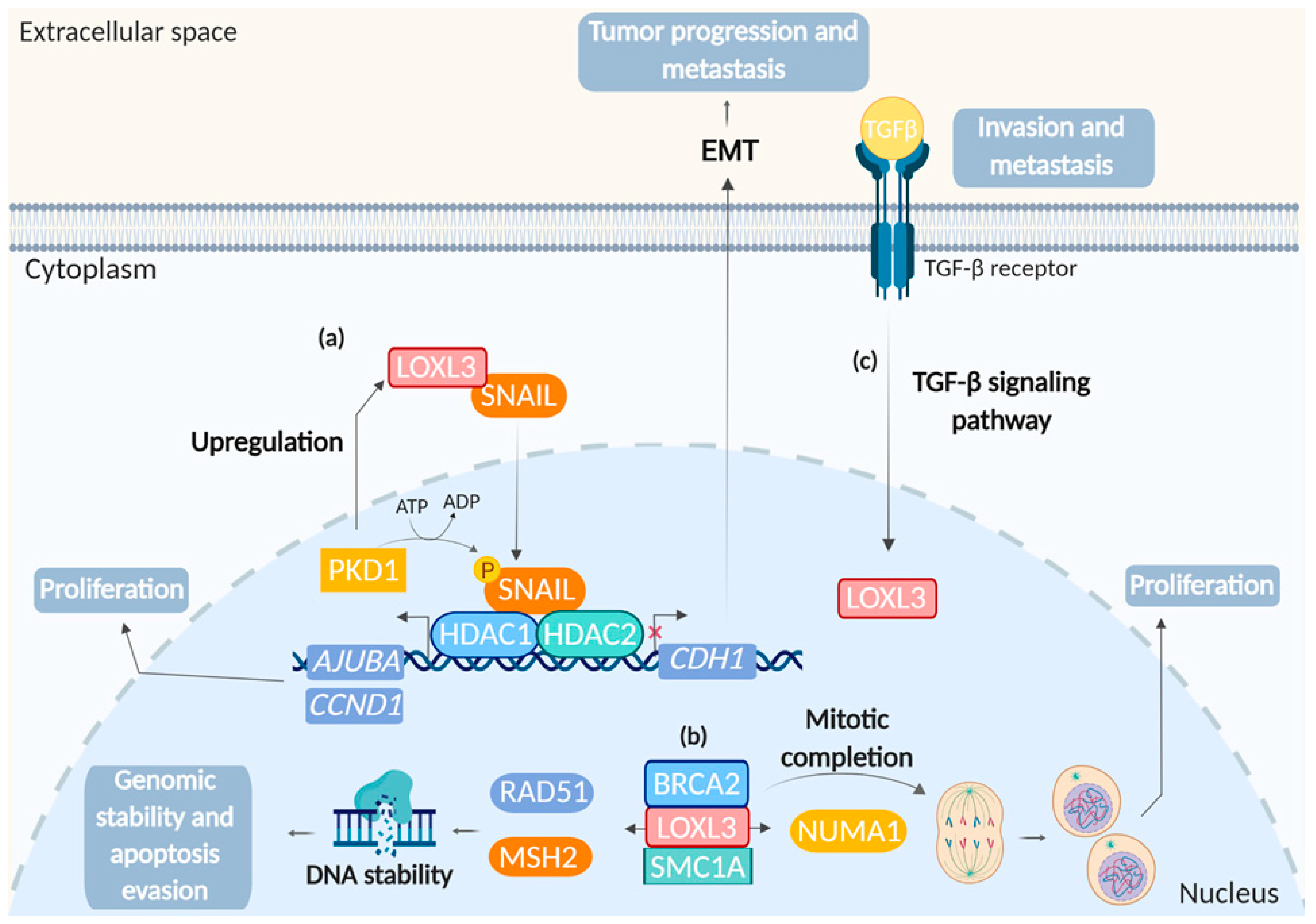
4.5. LOXL3 and Cancer
5. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AJUBA | Ajuba LIM protein |
| AP-1 | Activating protein-1 |
| AP-2 | Activating protein-2 |
| BMP-1 | Bone morphogenetic protein 1 |
| BRAF | B-Raf proto-oncogene |
| BRCA2 | Breast cancer 2 |
| CAAT | CCAAT sequence |
| CCND1 | Cyclin D1 gene |
| CDH1 | Cadherin-1 gene |
| COL2A1 | Collagen type II alpha 1 chain gene |
| COL11A1 | Collagen type XI alpha 1 chain gene |
| CRE | cAMP response element |
| c-Rel | Proto-oncogene c-Rel |
| CRL | Cytokine receptor-like |
| ECM | Extracellular matrix |
| FGF | Fibroblast growth factor 1 |
| EMT | Epithelial-mesenchymal transition |
| EST | Expressed sequence tag |
| FMT | Fibroblast-to-myofibroblast transition |
| GAGA | GAGA sequence |
| GATA | GATA sequence |
| GTEx | Genotype Tissue Expression |
| HDAC | Histone deacetylase |
| hTERT | Human telomerase reverse transcriptase |
| IPF | Idiopathic pulmonary fibrosis |
| IRF | Interferon regulatory factors |
| JNK | c-Jun N-terminal kinases |
| LOX | Lysyl oxidase |
| LOXL1 | Lysyl oxidase-like 1 |
| LOXL2 | Lysyl oxidase-like 2 |
| LOXL3 | Lysyl oxidase-like 3 |
| LOXL4 | Lysyl oxidase-like 4 |
| LTQ | Lysyl tyrosyl quinone |
| MIBP | Muscle integrin binding protein |
| MSH2 | MutS homolog 2 |
| NF-κB | Nuclear factor kappa B |
| NRSF | Neural restrictive silencer factor |
| NUDR | Nuclear DEAF-1-related |
| NUMA1 | Nuclear mitotic apparatus protein 1 |
| PAX | Paired box |
| PDGF | Platelet-derived growth factor |
| PKD1 | Protein kinase D1 |
| Pit1 | Pituitary transcript factor 1 |
| RAD51 | DNA repair protein RAD51 homolog 1 |
| RFX1 | Regulatory factor X1 |
| SRCR | Scavenger receptor cysteine-rich |
| SNAIL | Snail homolog 1 |
| SMC1A | Structural maintenance of chromosomes 1A |
| SMC3 | Structural maintenance of chromosomes 3 |
| SRF | Serum response element |
| SS | Stickler Syndrome |
| STAT3 | Signal transducer and activator of transcription 3 |
| TATA | TATAAA sequence |
| TGF | Transforming growth factor |
| TPM | Transcripts per million |
| UTR | Untranslated region |
| YY1 | Yin yang 1 |
| β-APN | β-aminopropionitrile |
References
- Barker, H.E.; Cox, T.R.; Erler, J.T. The rationale for targeting the LOX family in cancer. Nat. Rev. Cancer 2012, 12, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, T.; Eustace, A.; West, C. Lysyl oxidase: From basic science to future cancer treatment. Cell Struct. Funct. 2012, 37, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Mayorca-Guiliani, A.; Erler, J.T. The potential for targeting extracellular LOX proteins in human malignancy. Onco Targets Ther. 2013, 6, 1729–1735. [Google Scholar]
- Asuncion, L.; Fogelgren, B.; Fong, K.S.K.; Fong, S.F.T.; Kim, Y.; Csiszar, K. A novel human lysyl oxidase-like gene (LOXL4) on chromosome 10q24 has an altered scavenger receptor cysteine rich domain. Matrix Biol. 2001, 20, 487–491. [Google Scholar] [CrossRef]
- Mäki, J.M.; Kivirikko, K.I. Cloning and characterization of a fourth human lysyl oxidase isoenzyme. Biochem. J. 2001, 355, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Jourdan-Le, C.S.; Tomsche, A.; Ujfalusi, A.; Jia, L.; Csiszar, K. Central nervous system, uterus, heart, and leukocyte expression of the LOXL3 gene, encoding a novel lysyl oxidase-like protein. Genom 2001, 74, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Kim, Y. A tissue-specific variant of the human lysyl oxidase-like protein 3 (LOXL3) functions as an amine oxidase with substrate specificity. J. Biol. Chem. 2006, 281, 37282–37290. [Google Scholar] [CrossRef]
- Huang, Y.; Dai, J.; Tang, R.; Zhao, W.; Zhou, Z.; Wang, W.; Ying, K.; Xie, Y.; Mao, Y. Cloning and characterization of a human lysyl oxidase-like 3 gene (hLOXL3). Matrix Biol. 2001, 20, 153–157. [Google Scholar] [CrossRef]
- Uzel, M.I.; Scott, I.C.; Babakhanlou-Chase, H.; Palamakumbura, A.H.; Pappano, W.N.; Hong, H.H.; Greenspan, D.S.; Trackman, P.C. Multiple bone morphogenetic protein 1-related mammalian metalloproteinases process pro-lysyl oxidase at the correct physiological site and control lysyl oxidase activation in mouse embryo fibroblast cultures. J. Biol. Chem. 2001, 276, 22537–22543. [Google Scholar] [CrossRef]
- Panchenko, M.V.; Stetler-Stevenson, W.G.; Trubetskoy, O.V.; Gacheru, S.N.; Kagan, H.M. Metalloproteinase activity secreted by fibrogenic cells in the processing of prolysyl oxidase. Potential role of procollagen C-proteinase. J. Biol. Chem. 1996, 271, 7113–7119. [Google Scholar] [CrossRef]
- Ma, L.; Huang, C.; Wang, X.J.; Xin, D.E.; Wang, L.S.; Zou, Q.C.; Zhang, Y.S.; Tan, M.D.; Wang, Y.M.; Zhao, T.C.; et al. Lysyl Oxidase 3 is a Dual-Specificity Enzyme Involved in STAT3 Deacetylation and Deacetylimination Modulation. Mol. Cell 2017, 65, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Jeong, C.; Kim, Y. LOXL3-sv2, a novel variant of human lysyl oxidase-like 3 (LOXL3), functions as an amine oxidase. Int. J. Mol. Med. 2017, 39, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Consortium, G. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar]
- Mele, M.; Ferreira, P.G.; Reverter, F.; DeLuca, D.S.; Monlong, J.; Sammeth, M.; Young, T.R.; Goldmann, J.M.; Pervouchine, D.D.; Sullivan, T.J.; et al. The human transcriptome across tissues and individuals. Science 2015, 348, 660–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battle, A.; Brown, C.D.; Engelhardt, B.E.; Montgomery, S.B. Genetic effects on gene expression across human tissues. Nature 2017, 550, 204–213. [Google Scholar] [PubMed]
- Dudakova, L.; Sasaki, T.; Liskova, P.; Palos, M.; Jirsova, K. The presence of lysyl oxidase-like enzymes in human control and keratoconic corneas. Histol. Histopathol. 2016, 31, 63–71. [Google Scholar] [PubMed]
- Kraft-Sheleg, O.; Zaffryar-Eilot, S.; Genin, O.; Yaseen, W.; Soueid-Baumgarten, S.; Kessler, O.; Smolkin, T.; Akiri, G.; Neufeld, G.; Cinnamon, Y.; et al. Localized LoxL3-Dependent Fibronectin Oxidation Regulates Myofiber Stretch and Integrin-Mediated Adhesion. Dev. Cell 2016, 36, 550–561. [Google Scholar] [CrossRef] [Green Version]
- Peinado, H.; del Carmen Iglesias-de la Cruz, M.; Olmeda, D.; Csiszar, K.; Fong, K.S.; Vega, S.; Nieto, M.A.; Cano, A.; Portillo, F. A molecular role for lysyl oxidase-like 2 enzyme in snail regulation and tumor progression. EMBO J. 2005, 24, 3446–3458. [Google Scholar] [CrossRef]
- Santamaria, P.G.; Floristan, A.; Fontanals-Cirera, B.; Vazquez-Naharro, A.; Santos, V.; Morales, S.; Yuste, L.; Peinado, H.; Garcia-Gomez, A.; Portillo, F.; et al. Lysyl oxidase-like 3 is required for melanoma cell survival by maintaining genomic stability. Cell Death Differ. 2018, 25, 935–950. [Google Scholar] [CrossRef]
- Kasashima, H.; Yashiro, M.; Okuno, T.; Miki, Y.; Kitayama, K.; Masuda, G.; Kinoshita, H.; Morisaki, T.; Fukuoka, T.; Hasegawa, T.; et al. Significance of the Lysyl Oxidase Members Lysyl Oxidase Like 1, 3, and 4 in Gastric Cancer. Digestion 2018, 98, 238–248. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, B.; Hua, X.; Zhou, P.; Guo, L.; Peng, Y.; Qiu, K. Effect of newly identified hTERT-interacting proteins on telomerase activity. Acta. Biochim. Biophys. Sin. 2013, 45, 674–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Boxtel, A.L.; Gansner, J.M.; Hakvoort, H.W.; Snell, H.; Legler, J.; Gitlin, J.D. Lysyl oxidase-like 3b is critical for cartilage maturation during zebrafish craniofacial development. Matrix Biol. 2011, 30, 178–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Yang, R.; Liu, Z.; Hou, C.; Zong, W.; Zhang, A.; Sun, X.; Gao, J. Loss of lysyl oxidase-like 3 causes cleft palate and spinal deformity in mice. Hum. Mol. Genet. 2015, 24, 6174–6185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzahrani, F.; Al Hazzaa, S.A.; Tayeb, H.; Alkuraya, F.S. LOXL3, encoding lysyl oxidase-like 3, is mutated in a family with autosomal recessive Stickler syndrome. Hum. Genet. 2015, 134, 451–453. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.K.; Alkaabi, M.K.; ElBarky, A.M.; El-Hattab, A.W. LOXL3 novel mutation causing a rare form of autosomal recessive Stickler syndrome. Clin. Genet. 2019, 95, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.F.J.; Little, J.; Mossey, P.A.; Steegers-Theunissen, R.P.M.; Bonsi, M.; Bassi Andreasi, R.; Rubini, M. Association between a common missense variant in LOXL3 gene and the risk of non-syndromic cleft palate. Congenit. Anom. 2018, 58, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gao, B.; Xiao, X.; Li, S.; Jia, X.; Sun, W.; Guo, X.; Zhang, Q. Exome sequencing identified null mutations in LOXL3 associated with early-onset high myopia. Mol. Vis. 2016, 22, 161–167. [Google Scholar]
- Sethi, A.; Mao, W.; Wordinger, R.J.; Clark, A.F. Transforming growth factor-beta induces extracellular matrix protein cross-linking lysyl oxidase (LOX) genes in human trabecular meshwork cells. Invest. Ophthalmol. Vis. Sci. 2011, 52, 5240–5250. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Z.; Zhang, T.; Lin, Z.; Li, Z.; Zhang, A.; Sun, X.; Gao, J. Loss of Lysyl Oxidase-like 3 Attenuates Embryonic Lung Development in Mice. Sci. Rep. 2016, 6, 33856. [Google Scholar] [CrossRef]
- Jones, M.G.; Andriotis, O.G.; Roberts, J.J.; Lunn, K.; Tear, V.J.; Cao, L.; Ask, K.; Smart, D.E.; Bonfanti, A.; Johnson, P.; et al. Nanoscale dysregulation of collagen structure-function disrupts mechano-homeostasis and mediates pulmonary fibrosis. Elife 2018, 7, e36354. [Google Scholar] [CrossRef]
- Aumiller, V.; Strobel, B.; Romeike, M.; Schuler, M.; Stierstorfer, B.E.; Kreuz, S. Comparative analysis of lysyl oxidase (like) family members in pulmonary fibrosis. Sci. Rep. 2017, 7, 149. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, S.; Li, W. LOX/LOXL in pulmonary fibrosis: Potential therapeutic targets. J. Drug Target 2019, 27, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Philp, C.J.; Siebeke, I.; Clements, D.; Miller, S.; Habgood, A.; John, A.E.; Navaratnam, V.; Hubbard, R.B.; Jenkins, G.; Johnson, S.R. Extracellular Matrix Cross-Linking Enhances Fibroblast Growth and Protects against Matrix Proteolysis in Lung Fibrosis. Am. J. Respir. Cell Mol. Biol. 2018, 58, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Nave, A.H.; Mizikova, I.; Niess, G.; Steenbock, H.; Reichenberger, F.; Talavera, M.L.; Veit, F.; Herold, S.; Mayer, K.; Vadasz, I.; et al. Lysyl oxidases play a causal role in vascular remodeling in clinical and experimental pulmonary arterial hypertension. Arter. Thromb. Vasc. Biol. 2014, 34, 1446–1458. [Google Scholar] [CrossRef] [PubMed]
- Remus, E.W.; O’Donnell, R.E., Jr.; Rafferty, K.; Weiss, D.; Joseph, G.; Csiszar, K.; Fong, S.F.; Taylor, W.R. The role of lysyl oxidase family members in the stabilization of abdominal aortic aneurysms. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Horak, K.; Larson, D.F. Role of T lymphocytes in hypertension-induced cardiac extracellular matrix remodeling. Hypertension 2006, 48, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Vazquez, R.; Zabadi, S.; Watson, R.R.; Larson, D.F. T-lymphocytes mediate left ventricular fibrillar collagen cross-linking and diastolic dysfunction in mice. Matrix Biol. 2010, 29, 511–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Konomi, K.; Yamasaki, S.; Aratani, S.; Tsuchimochi, K.; Yokouchi, M.; Masuko-Hongo, K.; Yagishita, N.; Nakamura, H.; Komiya, S.; et al. Comparative analysis of gene expression profiles in intact and damaged regions of human osteoarthritic cartilage. Arthritis Rheum 2006, 54, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.M.; Du, S.H.; Huang, L.G.; Li, J.H.; Xiao, L.; Tong, P. Leptin promotes apoptosis and inhibits autophagy of chondrocytes through upregulating lysyl oxidase-like 3 during osteoarthritis pathogenesis. Osteoarthr. Cartil. 2016, 24, 1246–1253. [Google Scholar] [CrossRef] [Green Version]
- Eiseler, T.; Köhler, C.; Nimmagadda, S.C.; Jamali, A.; Funk, N.; Joodi, G.; Storz, P.; Seufferlein, T. Protein kinase D1 mediates anchorage-dependent and –independent growth of tumor cells via the zinc finger transcription factor Snail1. J. Biol. Chem. 2012, 287, 32367–32380. [Google Scholar] [CrossRef]
- Sebban, S.; Davidson, B.; Reich, R. Lysyl oxidase-like 4 is alternatively spliced in an anatomic site-specific manner in tumors involving the serosal cavities. Virchows Arch. 2009, 454, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.J.; Park, S.H.; Mun, S.H.; Kwak, S.G.; Lee, S.J.; Oh, H.K. Association between lysyl oxidase and fibrotic focus in relation with inflammation in breast cancer. Oncol. Lett. 2018, 15, 2431–2440. [Google Scholar] [CrossRef] [PubMed]
- Tadmor, T.; Bejar, J.; Attias, D.; Mischenko, E.; Sabo, E.; Neufeld, G.; Vadasz, Z. The expression of lysyl-oxidase gene family members in myeloproliferative neoplasms. Am. J. Hematol. 2013, 88, 355–358. [Google Scholar] [CrossRef] [PubMed]
- Dufresne, J.; Bowden, P.; Thavarajah, T.; Florentinus-Mefailoski, A.; Chen, Z.Z.; Tucholska, M.; Norzin, T.; Ho, M.T.; Phan, M.; Mohamed, N.; et al. The plasma peptides of ovarian cancer. Clin. Proteom. 2018, 15, 41. [Google Scholar] [CrossRef] [PubMed]
- Barbazan, J.; Muinelo-Romay, L.; Vieito, M.; Candamio, S.; Diaz-Lopez, A.; Cano, A.; Gomez-Tato, A.; de Cal, M.D.C.; Abal, M.; Lopez-Lopez, R.A. A multimarker panel for circulating tumor cells detection predicts patient outcome and therapy response in metastatic colorectal cancer. Int. J. Cancer 2014, 135, 2633–2643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Insua, Y.V.; de la Camara, J.; Vazquez, E.B.; Fernandez, A.; Rivera, F.V.; Silva, M.J.V.; Barbazan, J.; Muinelo-Romay, L.; Folgar, S.C.; Abalo, A.; et al. Predicting Outcome and Therapy Response in mCRC Patients Using an Indirect Method for CTCs Detection by a Multigene Expression Panel: A Multicentric Prospective Validation Study. Int. J. Mol. Sci. 2017, 18, 1265. [Google Scholar] [CrossRef] [PubMed]
- Hajdu, I.; Kardos, J.; Major, B.; Fabo, G.; Lorincz, Z.; Cseh, S.; Dorman, G. Inhibition of the LOX enzyme family members with old and new ligands. Selectivity analysis revisited. Bioorg. Med. Chem. Lett. 2018, 28, 3113–3118. [Google Scholar] [CrossRef] [PubMed]
- Schilter, H.; Findlay, A.D.; Perryman, L.; Yow, T.T.; Moses, J.; Zahoor, A.; Turner, C.I.; Deodhar, M.; Foot, J.S.; Zhou, W.; et al. The lysyl oxidase like 2/3 enzymatic inhibitor, PXS-5153A, reduces crosslinks and ameliorates fibrosis. J. Cell. Mol. Med. 2019, 23, 1759–1770. [Google Scholar] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laurentino, T.d.S.; Soares, R.d.S.; Marie, S.K.N.; Oba-Shinjo, S.M. LOXL3 Function Beyond Amino Oxidase and Role in Pathologies, Including Cancer. Int. J. Mol. Sci. 2019, 20, 3587. https://doi.org/10.3390/ijms20143587
Laurentino TdS, Soares RdS, Marie SKN, Oba-Shinjo SM. LOXL3 Function Beyond Amino Oxidase and Role in Pathologies, Including Cancer. International Journal of Molecular Sciences. 2019; 20(14):3587. https://doi.org/10.3390/ijms20143587
Chicago/Turabian StyleLaurentino, Talita de S., Roseli da S. Soares, Suely K. N. Marie, and Sueli M. Oba-Shinjo. 2019. "LOXL3 Function Beyond Amino Oxidase and Role in Pathologies, Including Cancer" International Journal of Molecular Sciences 20, no. 14: 3587. https://doi.org/10.3390/ijms20143587
APA StyleLaurentino, T. d. S., Soares, R. d. S., Marie, S. K. N., & Oba-Shinjo, S. M. (2019). LOXL3 Function Beyond Amino Oxidase and Role in Pathologies, Including Cancer. International Journal of Molecular Sciences, 20(14), 3587. https://doi.org/10.3390/ijms20143587