Circular RNAs: Biogenesis, Mechanism, and Function in Human Cancers

Abstract
:1. Introduction
2. The Biogenesis of circRNAs in Cancers
2.1. Introns Flank circRNAs
2.2. Regulation of circRNA Biogenesis by RNA-Binding Proteins (RBPs)
2.3. Impacts of Genomic Alterations on the Formation of circRNAs
3. Functional Mechanisms of circRNAs
3.1. Interfering with Parental Gene Regulation
3.2. Acting as miRNA Sponges
3.3. Binding to Proteins
3.4. Translating Proteins or Peptide
4. Perspectives
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| circRNA | Circular RNA |
| lncRNA | Long noncoding RNA |
| ciRNAs | Circular intronic RNAs |
| EIciRNAs | Exonic-intronic circRNAs |
| TricRNA | tRNA intronic circular RNA |
| RBPs | RNA-binding proteins |
| siRNA | Small interfering RNA |
| hESCs | Human embryonic stem cells |
| TCGA | The Cancer Genome Atlas |
| PMID | PubMed Unique Identifier |
| SNPs | Single nucleotide polymorphisms |
References
- Hsu, M.T.; Coca-Prados, M. Electron microscopic evidence for the circular form of RNA in the cytoplasm of eukaryotic cells. Nature 1979, 280, 339–340. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. Rna 2013, 19, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Bao, C.; Guo, W.; Li, S.; Chen, J.; Chen, B.; Luo, Y.; Lyu, D.; Li, Y.; Shi, G.; et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat. Commun. 2016, 7, 11215. [Google Scholar] [CrossRef]
- Maass, P.G.; Glazar, P.; Memczak, S.; Dittmar, G.; Hollfinger, I.; Schreyer, L.; Sauer, A.V.; Toka, O.; Aiuti, A.; Luft, F.C.; et al. A map of human circular RNAs in clinically relevant tissues. J. Mol. Med. 2017, 95, 1179–1189. [Google Scholar] [CrossRef] [PubMed]
- Piwecka, M.; Glazar, P.; Hernandez-Miranda, L.R.; Memczak, S.; Wolf, S.A.; Rybak-Wolf, A.; Filipchyk, A.; Klironomos, F.; Cerda Jara, C.A.; Fenske, P.; et al. Loss of a mammalian circular RNA locus causes miRNA deregulation and affects brain function. Science 2017, 357. [Google Scholar] [CrossRef]
- Cocquerelle, C.; Mascrez, B.; Hetuin, D.; Bailleul, B. Mis-splicing yields circular RNA molecules. Faseb J. 1993, 7, 155–160. [Google Scholar] [CrossRef]
- Vo, J.N.; Cieslik, M.; Zhang, Y.; Shukla, S.; Xiao, L.; Zhang, Y.; Wu, Y.M.; Dhanasekaran, S.M.; Engelke, C.G.; Cao, X.; et al. The Landscape of Circular RNA in Cancer. Cell 2019, 176, 869–881. [Google Scholar] [CrossRef]
- Zaphiropoulos, P.G. Circular RNAs from transcripts of the rat cytochrome P450 2C24 gene: correlation with exon skipping. Proc. Natl. Acad. Sci. USA 1996, 93, 6536–6541. [Google Scholar] [CrossRef]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef]
- Conn, S.J.; Pillman, K.A.; Toubia, J.; Conn, V.M.; Salmanidis, M.; Phillips, C.A.; Roslan, S.; Schreiber, A.W.; Gregory, P.A.; Goodall, G.J. The RNA binding protein quaking regulates formation of circRNAs. Cell 2015, 160, 1125–1134. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef]
- Abelson, J.; Trotta, C.R.; Li, H. tRNA splicing. J. Biol. Chem. 1998, 273, 12685–12688. [Google Scholar] [CrossRef]
- Salgia, S.R.; Singh, S.K.; Gurha, P.; Gupta, R. Two reactions of Haloferax volcanii RNA splicing enzymes: joining of exons and circularization of introns. Rna 2003, 9, 319–330. [Google Scholar] [CrossRef]
- Lu, Z.; Filonov, G.S.; Noto, J.J.; Schmidt, C.A.; Hatkevich, T.L.; Wen, Y.; Jaffrey, S.R.; Matera, A.G. Metazoan tRNA introns generate stable circular RNAs in vivo. Rna 2015, 21, 1554–1565. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Wilusz, J.E. Short intronic repeat sequences facilitate circular RNA production. Genes Dev. 2014, 28, 2233–2247. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.; Memczak, S.; Wyler, E.; Torti, F.; Porath, H.T.; Orejuela, M.R.; Piechotta, M.; Levanon, E.Y.; Landthaler, M.; Dieterich, C.; et al. Analysis of intron sequences reveals hallmarks of circular RNA biogenesis in animals. Cell Rep. 2015, 10, 170–177. [Google Scholar] [CrossRef]
- Han, L.; Diao, L.; Yu, S.; Xu, X.; Li, J.; Zhang, R.; Yang, Y.; Werner, H.M.J.; Eterovic, A.K.; Yuan, Y.; et al. The Genomic Landscape and Clinical Relevance of A-to-I RNA Editing in Human Cancers. Cancer Cell 2015, 28, 515–528. [Google Scholar] [CrossRef] [Green Version]
- Licatalosi, D.D.; Darnell, R.B. RNA processing and its regulation: global insights into biological networks. Nat. Rev. Genet. 2010, 11, 75–87. [Google Scholar] [CrossRef]
- Gerstberger, S.; Hafner, M.; Tuschl, T. A census of human RNA-binding proteins. Nat. Rev. Genet. 2014, 15, 829–845. [Google Scholar] [CrossRef]
- Neelamraju, Y.; Hashemikhabir, S.; Janga, S.C. The human RBPome: from genes and proteins to human disease. J. Proteom. 2015, 127, 61–70. [Google Scholar] [CrossRef]
- Wang, Z.L.; Li, B.; Luo, Y.X.; Lin, Q.; Liu, S.R.; Zhang, X.Q.; Zhou, H.; Yang, J.H.; Qu, L.H. Comprehensive Genomic Characterization of RNA-Binding Proteins across Human Cancers. Cell Rep. 2018, 22, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, C.X.; Xue, W.; Zhang, Y.; Jiang, S.; Yin, Q.F.; Wei, J.; Yao, R.W.; Yang, L.; Chen, L.L. Coordinated circRNA Biogenesis and Function with NF90/NF110 in Viral Infection. Mol. Cell 2017, 67, 214–227. [Google Scholar] [CrossRef]
- Cieply, B.; Park, J.W.; Nakauka-Ddamba, A.; Bebee, T.W.; Guo, Y.; Shang, X.; Lengner, C.J.; Xing, Y.; Carstens, R.P. Multiphasic and Dynamic Changes in Alternative Splicing during Induction of Pluripotency Are Coordinated by Numerous RNA-Binding Proteins. Cell Rep. 2016, 15, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.Y.; Li, T.C.; Wu, Y.Y.; Yeh, C.H.; Chiang, W.; Chuang, C.Y.; Kuo, H.C. The circular RNA circBIRC6 participates in the molecular circuitry controlling human pluripotency. Nat. Commun. 2017, 8, 1149. [Google Scholar] [CrossRef]
- Bass, B.L. RNA editing by adenosine deaminases that act on RNA. Annu. Rev. Biochem. 2002, 71, 817–846. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Guarnerio, J.; Bezzi, M.; Jeong, J.C.; Paffenholz, S.V.; Berry, K.; Naldini, M.M.; Lo-Coco, F.; Tay, Y.; Beck, A.H.; Pandolfi, P.P. Oncogenic Role of Fusion-circRNAs Derived from Cancer-Associated Chromosomal Translocations. Cell 2016, 165, 289–302. [Google Scholar] [CrossRef]
- Chen, N.; Zhao, G.; Yan, X.; Lv, Z.; Yin, H.; Zhang, S.; Song, W.; Li, X.; Li, L.; Du, Z.; et al. A novel FLI1 exonic circular RNA promotes metastasis in breast cancer by coordinately regulating TET1 and DNMT1. Genome Biol. 2018, 19, 218. [Google Scholar] [CrossRef]
- Wu, N.; Yuan, Z.; Du, K.Y.; Fang, L.; Lyu, J.; Zhang, C.; He, A.; Eshaghi, E.; Zeng, K.; Ma, J.; et al. Translation of yes-associated protein (YAP) was antagonized by its circular RNA via suppressing the assembly of the translation initiation machinery. Cell Death Differ. 2019. [Google Scholar] [CrossRef]
- Xu, J.Z.; Shao, C.C.; Wang, X.J.; Zhao, X.; Chen, J.Q.; Ouyang, Y.X.; Feng, J.; Zhang, F.; Huang, W.H.; Ying, Q.; et al. circTADA2As suppress breast cancer progression and metastasis via targeting miR-203a-3p/SOCS3 axis. Cell Death Dis. 2019, 10, 175. [Google Scholar] [CrossRef]
- Yang, R.; Xing, L.; Zheng, X.; Sun, Y.; Wang, X.; Chen, J. The circRNA circAGFG1 acts as a sponge of miR-195-5p to promote triple-negative breast cancer progression through regulating CCNE1 expression. Mol. Cancer 2019, 18, 4. [Google Scholar] [CrossRef]
- Zeng, K.; Chen, X.; Xu, M.; Liu, X.; Hu, X.; Xu, T.; Sun, H.; Pan, Y.; He, B.; Wang, S. CircHIPK3 promotes colorectal cancer growth and metastasis by sponging miR-7. Cell Death Dis. 2018, 9, 417. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, F.; Xiao, X.; Xie, F.; Tao, D.; Huang, C.; Liu, D.; Wang, M.; Wang, L.; Zeng, F.; et al. CircHIPK3 sponges miR-558 to suppress heparanase expression in bladder cancer cells. Embo Rep. 2017, 18, 1646–1659. [Google Scholar] [CrossRef]
- Fang, L.; Du, W.W.; Awan, F.M.; Dong, J.; Yang, B.B. The circular RNA circ-Ccnb1 dissociates Ccnb1/Cdk1 complex suppressing cell invasion and tumorigenesis. Cancer Lett. 2019, 459, 216–226. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Y.; Wu, S.; Zhou, Z.; Ding, X.; Shi, R.; Thorne, R.F.; Zhang, X.D.; Hu, W.; Wu, M. CircACC1 Regulates Assembly and Activation of AMPK Complex under Metabolic Stress. Cell Metab. 2019, 30, 157–173. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, F.; Fang, E.; Xiao, W.; Mei, H.; Li, H.; Li, D.; Song, H.; Wang, J.; Hong, M.; et al. Circular RNA circAGO2 drives cancer progression through facilitating HuR-repressed functions of AGO2-miRNA complexes. Cell Death Differ. 2019, 26, 1346–1364. [Google Scholar] [CrossRef]
- Du, W.W.; Yang, W.; Liu, E.; Yang, Z.; Dhaliwal, P.; Yang, B.B. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res. 2016, 44, 2846–2858. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.C.; Wong, C.W.; Liang, P.P.; Shi, M.; Cao, Y.; Rao, S.T.; Tsui, S.K.; Waye, M.M.; Zhang, Q.; Fu, W.M.; et al. Translation of the circular RNA circbeta-catenin promotes liver cancer cell growth through activation of the Wnt pathway. Genome Biol. 2019, 20, 84. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, K.; Xu, X.; Yang, Y.; Yan, S.; Wei, P.; Liu, H.; Xu, J.; Xiao, F.; Zhou, H.; et al. A peptide encoded by circular form of LINC-PINT suppresses oncogenic transcriptional elongation in glioblastoma. Nat. Commun. 2018, 9, 4475. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z. Efficient backsplicing produces translatable circular mRNAs. Rna 2015, 21, 172–179. [Google Scholar] [CrossRef]
- Abe, N.; Matsumoto, K.; Nishihara, M.; Nakano, Y.; Shibata, A.; Maruyama, H.; Shuto, S.; Matsuda, A.; Yoshida, M.; Ito, Y.; et al. Rolling Circle Translation of Circular RNA in Living Human Cells. Sci. Rep. 2015, 5, 16435. [Google Scholar] [CrossRef]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E.; et al. Translation of CircRNAs. Mol. Cell 2017, 66, 9–21. [Google Scholar] [CrossRef]
- Smid, M.; Wilting, S.M.; Uhr, K.; Rodriguez-Gonzalez, F.G.; de Weerd, V.; Prager-Van der Smissen, W.J.C.; van der Vlugt-Daane, M.; van Galen, A.; Nik-Zainal, S.; Butler, A.; et al. The circular RNome of primary breast cancer. Genome Res. 2019, 29, 356–366. [Google Scholar] [CrossRef]
- Sudmant, P.H.; Rausch, T.; Gardner, E.J.; Handsaker, R.E.; Abyzov, A.; Huddleston, J.; Zhang, Y.; Ye, K.; Jun, G.; Fritz, M.H.; et al. An integrated map of structural variation in 2,504 human genomes. Nature 2015, 526, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ran, Y.; Tao, C.; Li, S.; Chen, J.; Yang, E. Detection of circular RNA expression and related quantitative trait loci in the human dorsolateral prefrontal cortex. Genome Biol. 2019, 20, 99. [Google Scholar] [CrossRef]
- Zhang, F.; Zhao, X.; Dong, H.; Xu, J. circRNA expression analysis in lung adenocarcinoma: comparison of paired fresh frozen and formalin-fixed paraffin-embedded specimens. Biochem. Biophys. Res. Commun. 2018, 500, 738–743. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: a promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef] [Green Version]
- Bahn, J.H.; Zhang, Q.; Li, F.; Chan, T.M.; Lin, X.; Kim, Y.; Wong, D.T.; Xiao, X. The landscape of microRNA, Piwi-interacting RNA, and circular RNA in human saliva. Clin. Chem. 2015. [Google Scholar] [CrossRef]
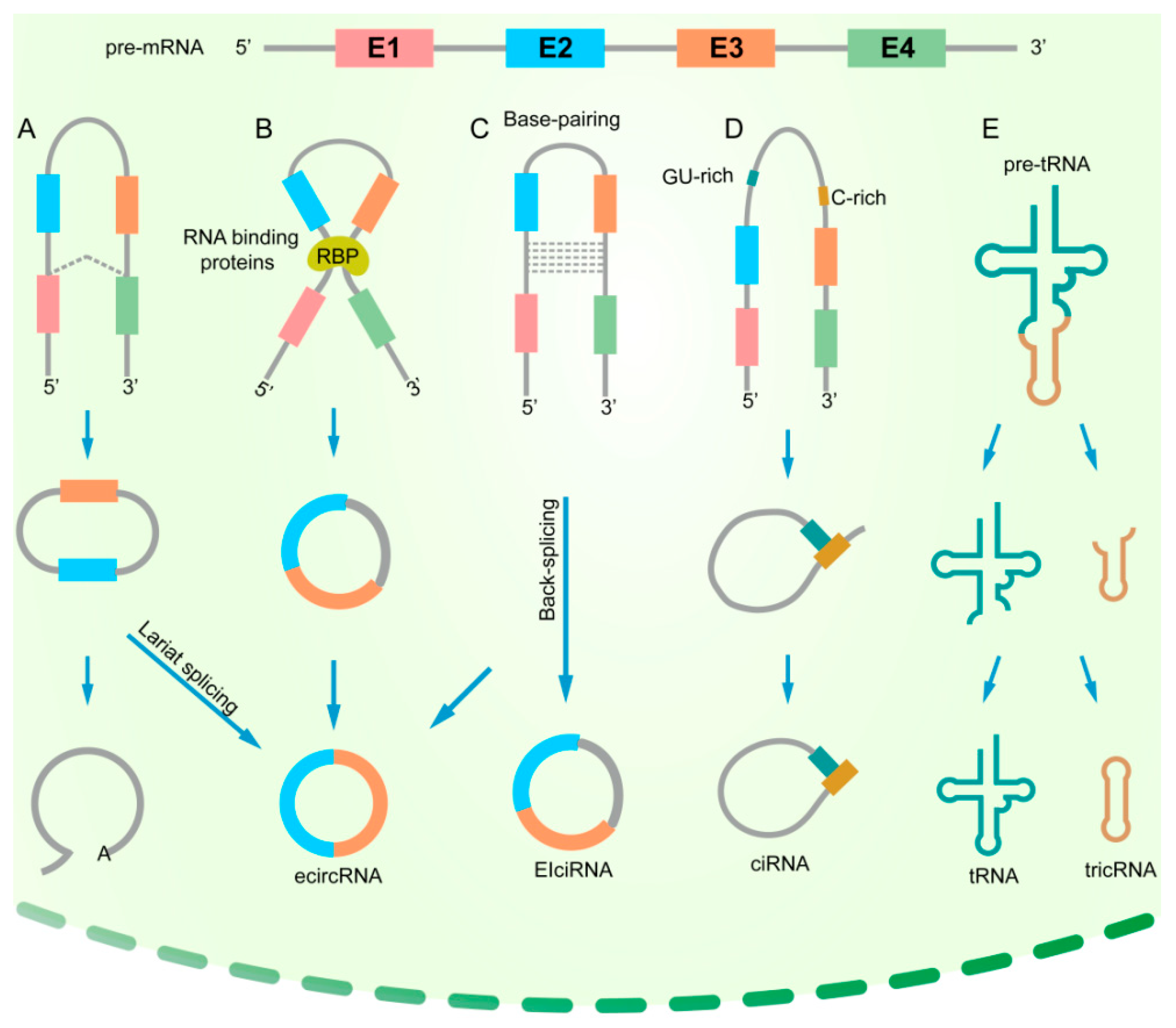


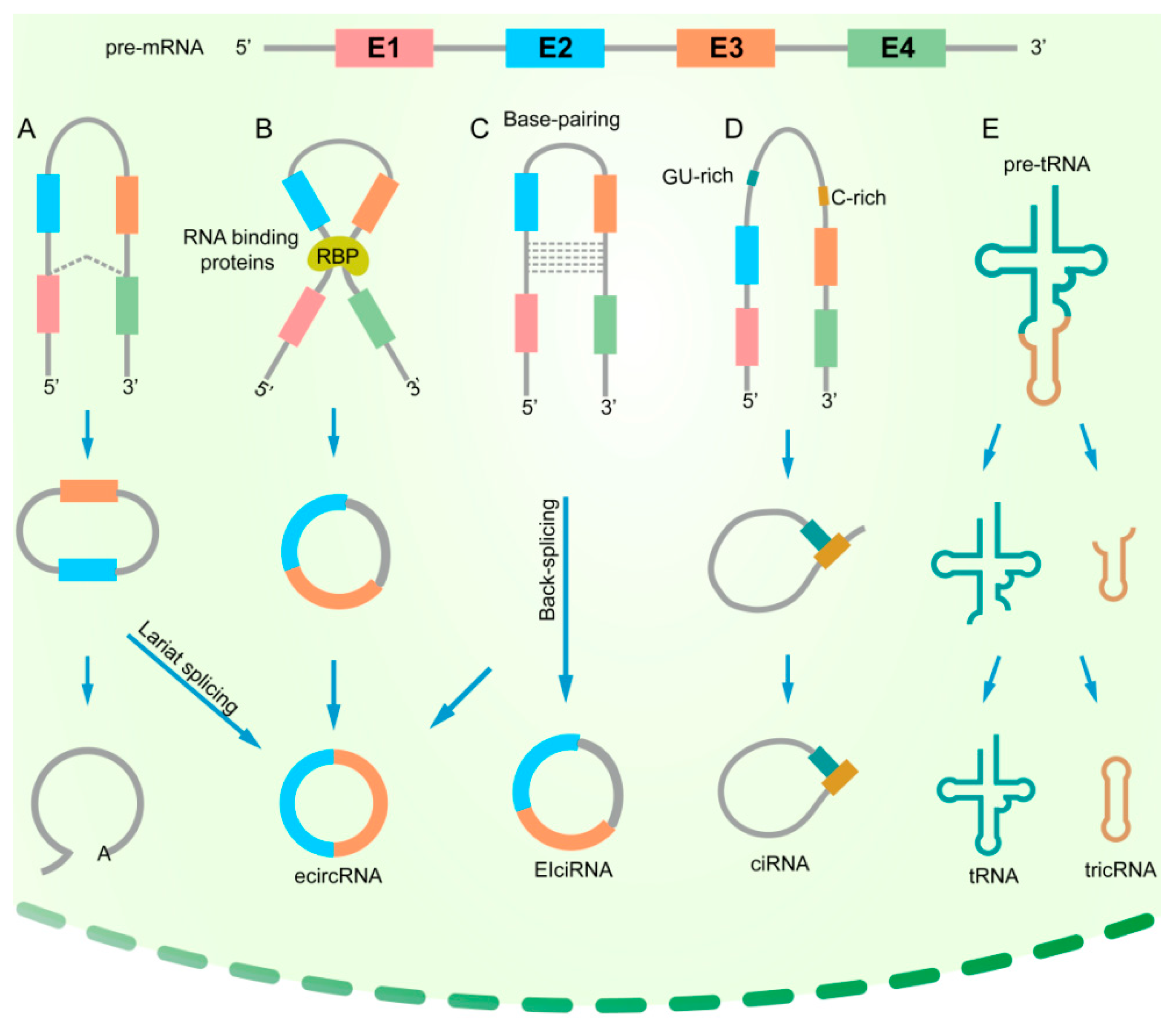{kind=link}
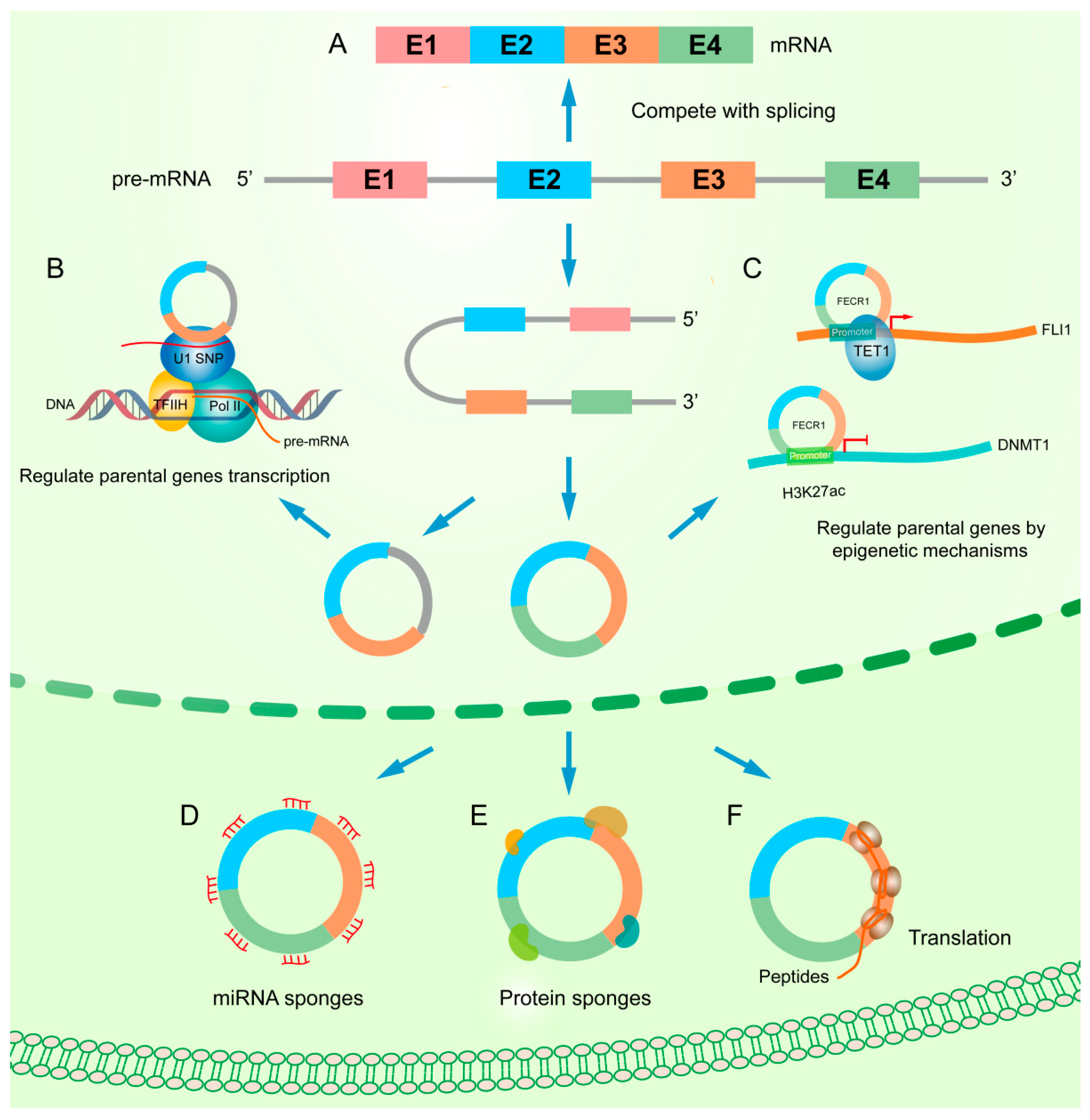{kind=link}
| Gene | Effect on the Formation of circRNA | Mechanisms | PMID |
|---|---|---|---|
| QKI | Promote | QKI enhances circRNA formation by binding to its consensus target single-stranded RNA (ssRNA) motif in introns that flank circRNA-forming exons. | 25768908 |
| TNRC6A | Promote | The RNA-binding protein TNRC6A binds to the flanking intron sequence of circHOMER1 and regulates the formation of circHOMER1. In the absence of TNRC6A, circHOMER1 cannot be effectively looped. | 29726904 |
| NF90/NF110 | Promote | NF90/NF110 stimulates circRNA formation by promoting the stability of intron complementary sequences | 28625552 |
| HNRNPL | Promote | Circular RNA formation regulated by HNRNPL back splicing. | 28611215 |
| MBL/MBNL1 | Promote | MBL/MBNL1 strongly and specifically binds with circMbl flanking introns, which contain conserved muscleblind binding sites and strongly affects circMbl biosynthesis | 25242144 |
| ESRP1 | Promote | The splicing factor ESRP1, which is controlled by the core pluripotency-associated factors, OCT4 and NANOG, can lead to the promotion of circBIRC6 generation. | 29074849 |
| FUS | Inhibit | FUS was found to regulate circRNA generation by binding introns that flank back-splicing junctions | 28358055 |
| DHX9 | Inhibit | DHX9 can bind to IRAlus and possesses RNA helicase activity. It was speculated that DHX9 may inhibit circRNA expression by unwinding RNA pairs that flank circularized exons. | 28355180 |
| ADAR1 | Inhibit | ADAR1 binds double-stranded RNA to mediate adenosine-to-inosine (A-to-I) RNA editing to inhibit circRNA formation. | 25558066 |
| Function | CircRNA | Cancer Type | Expression | Targeting miRNA or Genes | Mechanisms | PMID |
|---|---|---|---|---|---|---|
| Acting as miRNA sponge | circAGFG1 | TNBC | Up | miR-195-5p/CCNE1 | CircAGFG1 can promote TNBC cell proliferation, mobility, and invasion as well as tumorigenesis and metastasis in vivo by acting as a ceRNA (competing endogenous RNA) of miR-195-5p to relieve the repressive effect of miR-195-5p on its target cyclin E1 (CCNE1). | 30621700 |
| circHIPK3 | CRC | Up | miR-7/FAK, IGF1R, YY1, EGFR | circHIPK3 promotes proliferation/migration | 29549306 | |
| circCDR1 | ESCC | Up | miR-7/HOXB13 | CirsCDR1 functions as the sponge of miR-7 and reactivates its downstream HOXB13-mediated NF-κB/p65 pathway. | 30082829 | |
| circHIPK3 | BCa | Down | miR-558/HPSE | circHIPK3 targets miR-558 to suppress the expression of HPSE and inhibits migration, invasion, and angiogenesis of bladder cancer cells in vitro and suppresses bladder cancer growth and metastasis in vivo. | 28794202 | |
| circTRIM33-12 | HCC | Down | miR-191/ TET1 | CircTRIM33-12 upregulate TET1 expression by sponging miR-191, resulting in significantly reduced 5-hydroxymethylcytosine (5hmC) levels in HCC cells. | 31153371 | |
| circTADA2As | TNBC | Down | miR-203a-3p/SOCS3 | circTADA2As suppresses cell proliferation, migration, invasion, and clonogenicity and possesses a tumor-suppressor capability. | 30787278 | |
| circLARP4 | GC | Down | miR-424-5p/ LATS1 | circLARP4 is mainly localized in the cytoplasm and inhibits biological behaviors of GC cells by sponging miR-424. | 28893265 | |
| circATP2B1 | ccRCC | Down | miR-204-3p/FN1 | CircATP2B1 is suppressed by ERβ, and then reduces miR-204-3p, which increases fibronectin 1 expression and enhances ccRCC cell invasion. | 29490945 | |
| circITCH | BCa | Down | miR-17, miR-224/p21, PTEN | circITCH promotes the aggressive biological behaviors of BCa via up-regulating the expression of p21 and PTEN through ‘sponging’ miR-17 and miR-224 | 29386015 | |
| circMTO1 | HCC | Down | miR-9/ p21 | circMTO1 can down-regulate p21 by acting as the sponge of oncogenic miR-9 to suppress hepatocellular carcinoma progression. | 28520103 | |
| circDB | HCC | Up | miR-34a/USP7 | CircDB promotes tumor growth and reduces DNA damage by suppressing miR-34a and activating the USP7/Cyclin A2 signaling pathway | 30546088 | |
| Binding to proteins | CircACC1 | CRC | Up | AMPK | CircACC1 functions to stabilize and promote the enzymatic activity of the AMPK holoenzyme by forming a ternary complex with the regulatory β and γ subunits. | 31155494 |
| circDNMT1 | BRCA | Up | P53, AUF1 | Ectopically expressed circDnmt1 promotes the nuclear translocation of both p53 and AUF1, p53 nuclear translocation induces cellular autophagy, while AUF1 nuclear translocation reduces Dnmt1 mRNA instability, resulting in increased Dnmt1 translation. | 29973691 | |
| circAGO2 | GC | Up | HuR | circAGO2 binds with HuR protein to promote its activation and enrichment on the 3′-untranslated region of target genes, which reduces AGO2 binding and repression of AGO2/miRNA-mediated gene silencing | 30341421 | |
| circPABPN1 | Hela cell | Up | HuR | The binding of CircPABPN1 to HuR inhibits HuR binding to PABPN1 mRNA and reduces PABPN1 translation. | 28080204 | |
| circFOXO3 | BRCA | Down | circFOXO3, p53 | CircFoxo3 promotes MDM2-induced p53 ubiquitination and subsequent degradation by binding to Foxo3 protein and p53, resulting in cell apoptosis. | 27886165 | |
| circCCNB1 | BRCA | Down | Ccnb1/Cdk1 | CircCcnb1 can interact with both Ccnb1 and Cdk1 proteins. Ectopic delivery of circCcnb1 inhibits tumor growth and extends mouse viability. | 31199987 | |
| Translating proteins or peptide | circCTNNB1 | HCC | Up | 370-amino acid β-catenin isoform | CircCTNNB1 produces a novel, 370 amino acid β-catenin isoform that uses the start codon as the linear β-catenin mRNA transcript, and translation is terminated at a new stop codon created by circularization. | 31027518 |
| CircPINT | GBM | Up | PINT87aa | lncRNA-PINT can be translated into a small peptide to suppress glioblastoma cell proliferation | 30367041 | |
| CircE7 | Derived from human papillomavirus and presented in CESC and HNSC | Up | E7 protein | Specific disruption of circE7 in CaSki cervical carcinoma cells reduces E7 protein levels and inhibits cancer cell growth both in vitro and in tumor xenografts. | 31127091 | |
| circSHPRH | GBM | Down | SHPRH-146aa | SHPRH-146aa is a tumor suppressor in human glioblastoma, which is translated by circ-SHPRH. | 29343848 | |
| circFBXW7 | GBM | Down | FBXW7-185aa | The spanning junction open reading frame in circ-FBXW7 driven by internal ribosome entry site encodes a novel 21 kDa protein (FBXW7-185aa). Upregulation of FBXW7-185aa in cancer cells inhibits proliferation and cell cycle acceleration. | 28903484 | |
| Regulating parental gene expression at multiple levels | circFECR1 | BRCA | Up | TET1, DNMT1. | CircFECR1 regulates DNA methylation and demethylation enzymes to control breast cancer tumor growth. | 30537986 |
| circYAP | BRCA | Up | eIF4G, PABP | CircYap can bind with Yap mRNA and the translation initiation associated proteins, eIF4G and PABP, which functionally leads to the suppression of Yap translation initiation. | 31092884 | |
| circEIF3J, circPAIP2 | Hela, HEK293 | Up | U1 snRNA | EIciRNAs predominantly localizes in the nucleus, interacts with U1 snRNP, and promotes transcription of their parental genes. | 25664725 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Cai, Y.; Xu, J. Circular RNAs: Biogenesis, Mechanism, and Function in Human Cancers. Int. J. Mol. Sci. 2019, 20, 3926. https://doi.org/10.3390/ijms20163926
Zhao X, Cai Y, Xu J. Circular RNAs: Biogenesis, Mechanism, and Function in Human Cancers. International Journal of Molecular Sciences. 2019; 20(16):3926. https://doi.org/10.3390/ijms20163926
Chicago/Turabian StyleZhao, Xing, Yujie Cai, and Jianzhen Xu. 2019. "Circular RNAs: Biogenesis, Mechanism, and Function in Human Cancers" International Journal of Molecular Sciences 20, no. 16: 3926. https://doi.org/10.3390/ijms20163926
APA StyleZhao, X., Cai, Y., & Xu, J. (2019). Circular RNAs: Biogenesis, Mechanism, and Function in Human Cancers. International Journal of Molecular Sciences, 20(16), 3926. https://doi.org/10.3390/ijms20163926