Plastid Genomes of Carnivorous Plants Drosera rotundifolia and Nepenthes × ventrata Reveal Evolutionary Patterns Resembling Those Observed in Parasitic Plants

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Plastome Size and Gene Content in D. rotundifolia
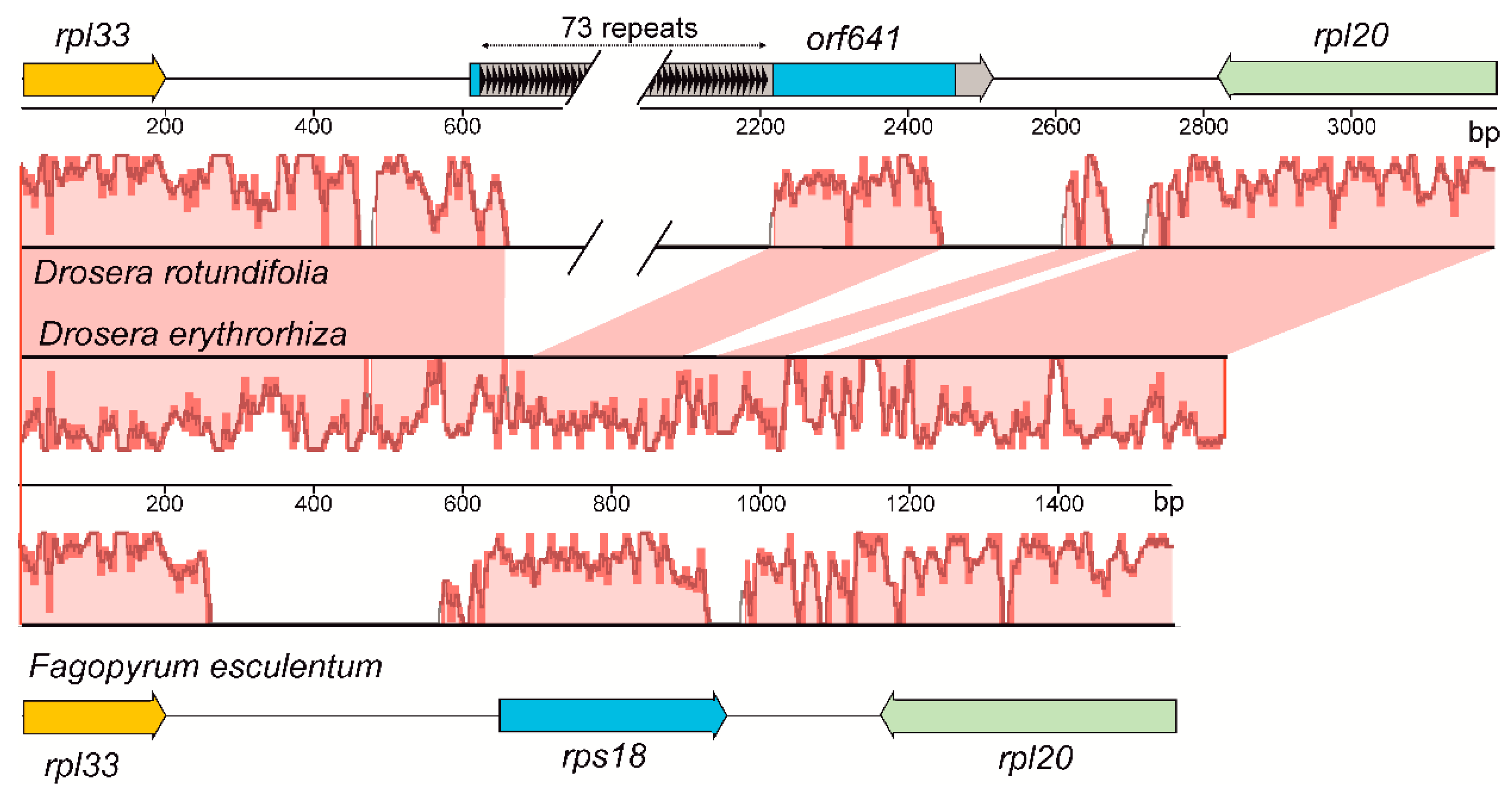
2.2. Structural Rearrangements and Duplications in the D. rotundifolia Plastome
2.3. The N. × ventrata Plastome Retains Conserved Structure and Gene Content
2.4. Identification and Prediction of RNA Editing Sites
3. Discussion
3.1. Evolution of the rps18 Gene in D. rotundifolia Plastome
3.2. Gene Loss and Genome Rearrangements in the Plastids in Three Families of Carnivorous Plants
3.3. Reduction of RNA Editing Sites in Plastomes of Some Carnivorous Plants
3.4. Convergent Plastid Genome Evolution in Carnivorous and Parasitic Plants
4. Materials and Methods
4.1. Plant Material and DNA Isolation
4.2. Sequencing and Assembly of the Plastid Genome of D. rotundifolia
4.3. Sequencing and Assembly of the Plastid Genome of N. × ventrata
4.4. Plastid Genome Annotation and Analysis Tools
4.5. RNA Editing Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| LSC | Large single-copy |
| SSC | Short single-copy |
| IR | Inverted repeat |
References
- Juniper, B.E. The path to plant carnivory. In Insects and the Plant Surface; Juniper, B.E., Southwood, T.R.E., Eds.; Edward Arnold: London, UK, 1986; pp. 195–218. [Google Scholar]
- Adamec, L. Mineral nutrition of carnivorous plants: A review. Bot. Rev. 1997, 63, 273–299. [Google Scholar] [CrossRef]
- Fleischmann, A.; Schlauer, J.; Smith, S.A.; Givnish, T.J. Evolution of carnivory in angiosperms. In Carnivorous Plants: Physiology, Ecology, and Evolution; Adamec, L., Ellison, A., Eds.; Oxford University Press: London, UK, 2017; pp. 22–42. [Google Scholar]
- Albert, V.A.; Williams, S.E.; Chase, M.W. Carnivorous plants: Phylogeny and structural evolution. Science 1992, 257, 1491–1495. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.G.; Almenara, D.P.; Winter, C.E.; Fritsch, P.W.; Lambers, H.; Oliveira, R.S. Underground leaves of Philcoxia trap and digest nematodes. Proc. Natl. Acad. Sci. USA 2012, 109, 1154–1158. [Google Scholar] [CrossRef] [PubMed]
- Givnish, T.J. New evidence on the origin of carnivorous plants. Proc. Natl. Acad. Sci. USA 2015, 112, 10–11. [Google Scholar] [CrossRef] [PubMed]
- Westwood, J.H.; Yoder, J.I.; Timko, M.P.; dePamphilis, C.W. The evolution of parasitism in plants. Trends Plant. Sci. 2010, 15, 227–235. [Google Scholar] [CrossRef] [PubMed]
- dePamphilis, C.W.; Palmer, J.D. Loss of photosynthetic and chlororespiratory genes from the plastid genome of a parasitic flowering plant. Nature 1990, 348, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Wakasugi, T.; Tsudzuki, T.; Sugiura, M. The genomics of land plant chloroplasts: Gene content and alteration of genomic information by RNA editing. Photosynth. Res. 2001, 70, 107–118. [Google Scholar] [CrossRef]
- Wicke, S.; Schneeweiss, G.M.; dePamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant. Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef]
- Wolfe, K.H.; Morden, C.W.; Palmer, J.D. Function and evolution of a minimal plastid genome from a nonphotosynthetic parasitic plant. Proc. Natl. Acad. Sci. USA 1992, 89, 10648–10652. [Google Scholar] [CrossRef]
- Barrett, C.F.; Freudenstein, J.V.; Li, J.; Mayfield-Jones, D.R.; Perez, L.; Pires, J.C.; Santos, C. Investigating the path of plastid genome degradation in an early-transitional clade of heterotrophic orchids, and implications for heterotrophic angiosperms. Mol. Biol. Evol. 2014, 31, 3095–3112. [Google Scholar] [CrossRef]
- Wicke, S.; Naumann, J. Molecular evolution of plastid genomes in parasitic flowering plants. Adv. Bot. Res. 2018, 85, 315–347. [Google Scholar]
- Barrett, C.F.; Davis, J.I. The plastid genome of the mycoheterotrophic Corallorhiza striata (Orchidaceae) is in the relatively early stages of degradation. Am. J. Bot. 2012, 99, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Funk, H.T.; Berg, S.; Krupinska, K.; Maier, U.G.; Krause, K. Complete DNA sequences of the plastid genomes of two parasitic flowering plant species, Cuscuta reflexa and Cuscuta gronovii. BMC Plant. Biol. 2007, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Delannoy, E.; Fujii, S.; des Francs-Small, C.C.; Brundrett, M.; Small, I. Rampant gene loss in the underground orchid Rhizanthella gardneri highlights evolutionary constraints on plastid genomes. Mol. Biol. Evol. 2011, 28, 2077–2086. [Google Scholar] [CrossRef] [PubMed]
- Wicke, S.; Müller, K.F.; de Pamphilis, C.W.; Quandt, D.; Wickett, N.J.; Zhang, Y.; Renner, S.S.; Schneeweiss, G.M. Mechanisms of functional and physical genome reduction in photosynthetic and nonphotosynthetic parasitic plants of the broomrape family. Plant Cell 2013, 25, 3711–3725. [Google Scholar] [CrossRef] [PubMed]
- Bellot, S.; Renner, S.S. The plastomes of two species in the endoparasite genus Pilostyles (Apodanthaceae) each retain just five or six possibly functional genes. Genome Biol. Evol. 2015, 8, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Lam, V.K.; Soto Gomez, M.; Graham, S.W. The highly reduced plastome of mycoheterotrophic Sciaphila (Triuridaceae) is colinear with its green relatives and is under strong purifying selection. Genome Biol. Evol. 2015, 7, 2220–2236. [Google Scholar] [CrossRef] [PubMed]
- Ravin, N.V.; Gruzdev, E.V.; Beletsky, A.V.; Mazur, A.M.; Prokhortchouk, E.B.; Filyushin, M.A.; Kochieva, E.Z.; Kadnikov, V.V.; Mardanov, A.V.; Skryabin, K.G. The loss of photosynthetic pathways in the plastid and nuclear genomes of the non-photosynthetic mycoheterotrophic eudicot Monotropa hypopitys. BMC Plant Biol. 2016, 16(3), 238. [Google Scholar] [CrossRef]
- Molina, J.; Hazzouri, K.M.; Nickrent, D.; Geisler, M.; Meyer, R.S.; Pentony, M.M.; Flowers, J.M.; Pelser, P.; Barcelona, J.; Inovejas, S.A.; et al. Possible loss of the chloroplast genome in the parasitic flowering plant Rafflesia lagascae (Rafflesiaceae). Mol. Biol. Evol. 2014, 31, 793–803. [Google Scholar] [CrossRef]
- Wicke, S.; Schäferhoff, B.; dePamphilis, C.W.; Müller, K.F. Disproportional plastome-wide increase of substitution rates and relaxed purifying selection in genes of carnivorous Lentibulariaceae. Mol. Biol. Evol. 2014, 31, 529–545. [Google Scholar] [CrossRef]
- Silva, S.R.; Diaz, Y.C.; Penha, H.A.; Pinheiro, D.G.; Fernandes, C.C.; Miranda, V.F.; Michael, T.P.; Varani, A.M. The chloroplast genome of Utricularia reniformis sheds light on the evolution of the ndh gene complex of terrestrial carnivorous plants from the Lentibulariaceae family. PLoS ONE 2016, 11, e0165176. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.R.; Michael, T.P.; Meer, E.J.; Pinheiro, D.G.; Varani, A.M.; Miranda, V.F.O. Comparative genomic analysis of Genlisea (corkscrew plants-Lentibulariaceae) chloroplast genomes reveals an increasing loss of the ndh genes. PLoS ONE 2018, 13, e0190321. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, A.; Cross, A.T.; Gibson, R.; Gonella, P.M.; Dixon, K.W. Systematics and evolution of Droseraceae. In Carnivorous Plants: Physiology, Ecology, and Evolution; Adamec, L., Ellison, A., Eds.; Oxford University Press: London, UK, 2017; pp. 45–57. [Google Scholar]
- Meimberg, H.; Heubl, G. Introduction of a nuclear marker for phylogenetic analysis of Nepenthaceae. Plant Biol. 2006, 8, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Mithöfer, A. Carnivorous pitcher plants: Insights in an old topic. Phytochemistry 2011, 72, 1678–1682. [Google Scholar] [CrossRef] [PubMed]
- Fleming, R. Hybrid Nepenthes. Carniv. Plant Newslett. 1979, 8, 10–12. [Google Scholar]
- Hatano, N.; Hamada, T. Proteome analysis of pitcher fluid of the carnivorous plant Nepenthes alata. J. Proteome Res. 2008, 7, 809–816. [Google Scholar] [CrossRef]
- Wan Zakaria, W.N.; Loke, K.K.; Goh, H.H.; Mohd Noor, N. RNA-seq analysis for plant carnivory gene discovery in Nepenthes × ventrata. Genom. Data 2016, 7, 18–19. [Google Scholar] [CrossRef]
- Nevill, P.G.; Howell, K.A.; Cross, A.T.; Williams, A.V.; Zhong, X.; Tonti-Filippini, J.; Boykin, L.M.; Dixon, K.W.; Small, I. Plastome-wide rearrangements and gene losses in carnivorous Droseraceae. Genome Biol. Evol. 2019, 11, 472–485. [Google Scholar] [CrossRef]
- Zhu, Z.-X.; Wang, J.-H.; Chen, C.-R.; Zhao, K.-K.; Wang, H.-F. Complete plastome sequence of Nepenthes mirabilis (Nepenthaceae): A ‘vulnerable’ herb in China. Mitochondrial DNA B 2018, 3, 732–733. [Google Scholar] [CrossRef]
- Yao, G.; Jin, J.J.; Li, H.T.; Yang, J.B.; Mandala, V.S.; Croley, M.; Mostow, R.; Douglas, N.A.; Chase, M.W.; Christenhusz, M.J.M.; et al. Plastid phylogenomic insights into the evolution of Caryophyllales. Mol. Phylogenet. Evol. 2019, 134, 74–86. [Google Scholar] [CrossRef]
- Logacheva, M.D.; Samigullin, T.H.; Dhingra, A.; Penin, A.A. Comparative chloroplast genomics and phylogenetics of Fagopyrum esculentum ssp. ancestrale -a wild ancestor of cultivated buckwheat. BMC Plant. Biol. 2008, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Kode, V.; Mudd, E.A.; Iamtham, S.; Day, A. The tobacco plastid accD gene is essential and is required for leaf development. Plant J. 2005, 44, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Krause, K. Plastid genomes of parasitic plants: A trail of reductions and losses. In Organelle genetics; Bullerwell, C.E., Ed.; Springer: Berlin, Germany, 2012; pp. 79–103. [Google Scholar]
- Sloan, D.B.; Triant, D.A.; Forrester, N.J.; Bergner, L.M.; Wu, M.; Taylor, D.R. A recurring syndrome of accelerated plastid genome evolution in the angiosperm tribe Sileneae (Caryophyllaceae). Mol. Phylogenet. Evol. 2014, 72, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Zoschke, R.; Nakamura, M.; Liere, K.; Sugiura, M.; Börner, T.; Schmitz-Linneweber, C. An organellar maturase associates with multiple group II introns. Proc. Natl. Acad. Sci. USA 2010, 107, 3245–3250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffy, A.M.; Kelchner, S.A.; Wolf, P.G. Conservation of selection on matK following an ancient loss of its flanking intron. Gene 2009, 438, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeal, J.R.; Kuehl, J.V.; Boore, J.L.; Leebens-Mack, J.; dePamphilis, C.W. Parallel loss of plastid introns and their maturase in the genus Cuscuta. PLoS ONE 2009, 4, e5982. [Google Scholar] [CrossRef] [PubMed]
- Chumley, T.W.; Palmer, J.D.; Mower, J.P.; Fourcade, H.M.; Calie, P.J.; Boore, J.L.; Jansen, R.K. The complete chloroplast genome sequence of Pelargonium x hortorum: Organization and evolution of the largest and most highly rearranged chloroplast genome of land plants. Mol. Biol. Evol. 2006, 23, 2175–2190. [Google Scholar] [CrossRef]
- Cai, Z.; Guisinger, M.; Kim, H.G.; Ruck, E.; Blazier, J.C.; McMurtry, V.; Kuehl, J.V.; Boore, J.; Jansen, R.K. Extensive reorganization of the plastid genome of Trifolium subterraneum (Fabaceae) is associated with numerous repeated sequences and novel DNA insertions. J. Mol. Evol. 2008, 67, 696–704. [Google Scholar] [CrossRef]
- Levinson, G.; Gutman, G.A. Slipped-strand mispairing: A major mechanism for DNA sequence evolution. Mol. Biol. Evol. 1987, 4, 203–221. [Google Scholar]
- Shinozaki, K.; Ohme, M.; Tanaka, M.; Wakasugi, T.; Hayashida, N.; Matsubayashi, T.; Zaita, N.; Chunwongse, J.; Obokata, J.; Yamaguchi-Shinozaki, K.; et al. The complete nucleotide sequence of the tobacco chloroplast genome: Its gene organization and expression. EMBO J. 1986, 5, 2043–2049. [Google Scholar] [CrossRef]
- Bock, R. Sense from nonsense: How the genetic information of chloroplasts is altered by RNA editing. Biochimie 2000, 82, 549–557. [Google Scholar] [CrossRef]
- Zeng, W.H.; Liao, S.C.; Chang, C.C. Identification of RNA editing sites in chloroplast transcripts of Phalaenopsis aphrodite and comparative analysis with those of other seed plants. Plant Cell Physiol. 2007, 48, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Puerta, M.V. Involvement of plastid, mitochondrial and nuclear genomes in plant-to-plant horizontal gene transfer. Acta Soc. Bot. Pol. 2014, 83, 317–323. [Google Scholar] [CrossRef]
- Davis, C.C.; Xi, Z. Horizontal gene transfer in parasitic plants. Curr. Opin. Plant. Biol. 2015, 26, 14–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millen, R.S.; Olmstead, R.G.; Adams, K.L.; Palmer, J.D.; Lao, N.T.; Heggie, L.; Kavanagh, T.A.; Hibberd, J.M.; Gray, J.C.; Morden, C.W.; et al. Many parallel losses of infA from chloroplast DNA during angiosperm evolution with multiple independent transfers to the nucleus. Plant Cell 2001, 13, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Gantt, J.S.; Baldauf, S.L.; Calie, P.J.; Weeden, N.F.; Palmer, J.D. Transfer of rpl22 to the nucleus greatly preceded its loss from the chloroplast and involved the gain of an intron. EMBO J. 1991, 10, 3073–3078. [Google Scholar] [CrossRef]
- Jansen, R.K.; Cai, Z.; Raubeson, L.A.; Daniell, H.; Depamphilis, C.W.; Leebens-Mack, J.; Müller, K.F.; Guisinger-Bellian, M.; Haberle, R.C.; Hansen, A.K.; et al. Analysis of 81 genes from 64 plastid genomes resolves relationships in angiosperms and identifies genome-scale evolutionary patterns. Proc. Natl. Acad. Sci. USA 2007, 104, 19369–19374. [Google Scholar] [CrossRef] [Green Version]
- Ueda, M.; Fujimoto, M.; Arimura, S.; Murata, J.; Tsutsumi, N.; Kadowaki, K. Loss of the rpl32 gene from the chloroplast genome and subsequent acquisition of a preexisting transit peptide within the nuclear gene in Populus. Gene 2007, 402, 51–56. [Google Scholar] [CrossRef]
- Ueda, M.; Nishikawa, T.; Fujimoto, M.; Takanashi, H.; Arimura, S.; Tsutsumi, N.; Kadowaki, K. Substitution of the gene for chloroplast RPS16 was assisted by generation of a dual targeting signal. Mol. Biol. Evol. 2008, 25, 1566–1575. [Google Scholar] [CrossRef]
- McNeal, J.R.; Kuehl, J.V.; Boore, J.L.; de Pamphilis, C.W. Complete plastid genome sequences suggest strong selection for retention of photosynthetic genes in the parasitic plant genus Cuscuta. BMC Plant Biol. 2007, 7, 57. [Google Scholar] [CrossRef]
- Kim, H.T.; Kim, J.S.; Moore, M.J.; Neubig, K.M.; Williams, N.H.; Whitten, W.M.; Kim, J.H. Seven new complete plastome sequences reveal rampant independent loss of the ndh gene family across orchids and associated instability of the inverted repeat/small single-copy region boundaries. PLoS ONE 2015, 10, e0142215. [Google Scholar] [CrossRef] [PubMed]
- Wakasugi, T.; Tsudzuki, J.; Ito, S.; Nakashima, K.; Tsudzuki, T.; Sugiura, M. Loss of all ndh genes as determined by sequencing the entire chloroplast genome of the black pine Pinus thunbergii. Proc. Natl. Acad. Sci. USA 1994, 91, 9794–9798. [Google Scholar] [CrossRef] [PubMed]
- Rumeau, D.; Peltier, G.; Cournac, L. Chlororespiration and cyclic electron flow around PSI during photosynthesis and plant stress response. Plant Cell Environ. 2007, 30, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Horváth, E.M.; Peter, S.O.; Joët, T.; Rumeau, D.; Cournac, L.; Horváth, G.V.; Kavanagh, T.A.; Schäfer, C.; Peltier, G.; Medgyesy, P. Targeted inactivation of the plastid ndhB gene in tobacco results in an enhanced sensitivity of photosynthesis to moderate stomatal closure. Plant Physiol. 2000, 123, 1337–1350. [Google Scholar] [CrossRef] [PubMed]
- Ruhlman, T.A.; Chang, W.J.; Chen, J.J.; Huang, Y.T.; Chan, M.T.; Zhang, J.; Liao, D.C.; Blazier, J.C.; Jin, X.; Shih, M.C.; et al. NDH expression marks major transitions in plant evolution and reveals coordinate intracellular gene loss. BMC Plant Biol. 2015, 15, 100. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Laclette, E.; Lyons, E.; Hernández-Guzmán, G.; Pérez-Torres, C.A.; Carretero-Paulet, L.; Chang, T.H.; Lan, T.; Welch, A.J.; Juárez, M.J.; Simpson, J.; et al. Architecture and evolution of a minute plant genome. Nature 2013, 498, 94–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drescher, A.; Ruf, S.; Calsa, T., Jr.; Carrer, H.; Bock, R. The two largest chloroplast genome-encoded open reading frames of higher plants are essential genes. Plant J. 2000, 22, 97–104. [Google Scholar] [CrossRef]
- Kikuchi, S.; Bédard, J.; Hirano, M.; Hirabayashi, Y.; Oishi, M.; Imai, M.; Takase, M.; Ide, T.; Nakai, M. Uncovering the protein translocon at the chloroplast inner envelope membrane. Science. 2013, 339, 571–574. [Google Scholar] [CrossRef]
- Bölter, B.; Soll, J. Ycf1/Tic214 is not essential for the accumulation of plastid proteins. Mol. Plant. 2017, 10, 219–221. [Google Scholar] [CrossRef]
- Petersen, G.; Cuenca, A.; Seberg, O. Plastome evolution in hemiparasitic mistletoes. Genome Biol. Evol. 2015, 7, 2520–2532. [Google Scholar] [CrossRef]
- Rischer, H.; Hamm, A.; Bringmann, G. Nepenthes insignis uses a C2-portion of the carbon skeleton of L-alanine acquired via its carnivorous organs, to build up the allelochemical plumbagin. Phytochem. 2002, 59, 603–609. [Google Scholar] [CrossRef]
- Pavlovič, A.; Krausko, M.; Libiaková, M.; Adamec, L. Feeding on prey increases photosynthetic efficiency in the carnivorous sundew Drosera capensis. Ann. Bot. 2014, 113, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Gruzdev, E.V.; Mardanov, A.V.; Beletsky, A.V.; Ravin, N.V.; Skryabin, K.G. The complete mitochondrial genome of the carnivorous flowering plant Nepenthes x ventrata. Mitochondrial DNA B 2018, 3, 1259–1260. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics. 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics. 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Garrison, E.; Marth, G. Haplotype-based variant detection from short-read sequencing. arXiv 2012, arXiv:1207.3907. Available online: https://arxiv.org/abs/1207.3907 (accessed on 21 August 2019).
- Cingolani, P.; Platts, A.; Wang le, L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Mower, J.P. PREP-Mt: Predictive RNA editor for plant mitochondrial genes. BMC Bioinform. 2005, 6, 96. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Function | Genes |
|---|---|
| Photosystem I | psaA, psaB, psaC, psaI, psaJ, ycf3i, ycf4 |
| Photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ |
| Cytochrome b6/f complex | petA, petBi, petDi, petG, petL, petN, ccsA |
| ATP synthase | atpA, atpB, atpE, atpFi, atpH, atpI |
| RNA polymerase | rpoA, rpoB, rpoC1i, rpoC2 |
| Ribosomal proteins (large subunit) | rpl2, rpl14, rpl16i, rpl20, rpl22, rpl23, rpl32, rpl33, rpl36 |
| Ribosomal proteins (small subunit) | rps2, rps3, rps4, rps7, rps8, rps11, rps12i, rps14 (x2), rps15, rps16, rps19 |
| Other protein-coding genes | rbcL, infA, matK, cemA, clpP, accD, ycf68 |
| rRNAs | rrn16, rrn23, rrn4.5, rrn5 |
| tRNAs | trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnG-GCC, trnH-GUG, trnI-CAU, trnI-GAUi, trnK-UUU, trnL-CAA, trnL-UAAi, trnL-UAG, trnM-CAU (x2), trnfM-CAU, trnN-GUU, trnP-UGG (x2), trnQ-UUG, trnR-ACG, trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GAC, trnW-CCA, trnY-GUA |
| Pseudogenes | ψrps18(orf641), ψycf15 |
| Pseudogenes present along with an intact copy | ψinfA, ψpsaB, ψpsaJ, ψpsbI, ψpsbJ, ψrpl2 |
| Species | Plastome Size (bp) | Repetitive Sequences (bp) | Repetitive Sequences (%) | Tandem Repeats (bp) |
|---|---|---|---|---|
| Caryophyllales; Droseraceae | ||||
| Drosera rotundifolia | 192,912 | 32,380 | 23.13% | 736 |
| Drosera erythrorhiza | 134,391 | 4708 | 4.84% | 164 |
| Drosera regia | 136,810 | 1883 | 1.66% | 175 |
| Dionaea muscipula | 117,589 | 5599 | 4.90% | 460 |
| Aldrovanda vesiculosa | 141,568 | 2514 | 2.20% | 326 |
| Caryophyllales; Nepenthaceae | ||||
| Nepenthes × ventrata | 156,637 | 664 | 0.51% | 82 |
| Nepenthes mirabilis | 156,381 | 5426 | 4.20% | 237 |
| Lamiales; Lentibulariaceae | ||||
| Pinguicula ehlersiae | 147,147 | 893 | 0.74% | 151 |
| Utricularia macrorhiza | 153,228 | 632 | 0.50% | 476 |
| Utricularia reniformis | 139,725 | 682 | 0.59% | 60 |
| Utricularia gibba | 152,113 | 780 | 0.63% | 225 |
| Genlisea margaretae | 141,255 | 500 | 0.43% | 440 |
| Genlisea aurea | 140,010 | 586 | 0.51% | 251 |
| Genlisea filiformis | 140,308 | 622 | 0.54% | 204 |
| Genlisea pygmaea | 140,466 | 467 | 0.40% | 227 |
| Genlisea repens | 140,432 | 467 | 0.40% | 225 |
| Genlisea tuberosa | 140,677 | 563 | 0.49% | 208 |
| Genlisea violacea | 143,416 | 769 | 0.65% | 60 |
| Caryophyllales; Caryophyllaceae | ||||
| Silene noctiflora | 151,639 | 2852 | 2.34% | 225 |
| Silene chalcedonica | 148,081 | 4566 | 3.67% | 135 |
| Silene conica | 147,208 | 1785 | 1.48% | 40 |
| Silene conoidea | 147,896 | 1615 | 1.33% | 151 |
| Silene paradoxa | 151,632 | 2023 | 1.60% | 256 |
| Silene latifolia | 151,736 | 1189 | 0.94% | 20 |
| Silene vulgaris | 151,583 | 1121 | 0.89% | 181 |
| Agrostemma githago | 151,733 | 1297 | 1.03% | 105 |
| Gene | Amino Acid Position | Codon * | Amino Acid Change | Editing Frequency | PREP-Cp Editing Score |
|---|---|---|---|---|---|
| atpF | 31 | CcA | P=>L | 98% | 0.86 |
| rps2 | 54 | AcA | T=>I | 72% | 0.71 |
| rps2 | 92 | TcA | S=>L | 93% | 1.00 |
| rps14 | 50 | CcA | P=>L | 67% | 1.00 |
| rpl22 | 73 | TcA | S=>L | 70% | ** |
| rpl23 | 24 | TcT | S=>F | 81% | 0.71 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gruzdev, E.V.; Kadnikov, V.V.; Beletsky, A.V.; Kochieva, E.Z.; Mardanov, A.V.; Skryabin, K.G.; Ravin, N.V. Plastid Genomes of Carnivorous Plants Drosera rotundifolia and Nepenthes × ventrata Reveal Evolutionary Patterns Resembling Those Observed in Parasitic Plants. Int. J. Mol. Sci. 2019, 20, 4107. https://doi.org/10.3390/ijms20174107
Gruzdev EV, Kadnikov VV, Beletsky AV, Kochieva EZ, Mardanov AV, Skryabin KG, Ravin NV. Plastid Genomes of Carnivorous Plants Drosera rotundifolia and Nepenthes × ventrata Reveal Evolutionary Patterns Resembling Those Observed in Parasitic Plants. International Journal of Molecular Sciences. 2019; 20(17):4107. https://doi.org/10.3390/ijms20174107
Chicago/Turabian StyleGruzdev, Eugeny V., Vitaly V. Kadnikov, Alexey V. Beletsky, Elena Z. Kochieva, Andrey V. Mardanov, Konstantin G. Skryabin, and Nikolai V. Ravin. 2019. "Plastid Genomes of Carnivorous Plants Drosera rotundifolia and Nepenthes × ventrata Reveal Evolutionary Patterns Resembling Those Observed in Parasitic Plants" International Journal of Molecular Sciences 20, no. 17: 4107. https://doi.org/10.3390/ijms20174107
APA StyleGruzdev, E. V., Kadnikov, V. V., Beletsky, A. V., Kochieva, E. Z., Mardanov, A. V., Skryabin, K. G., & Ravin, N. V. (2019). Plastid Genomes of Carnivorous Plants Drosera rotundifolia and Nepenthes × ventrata Reveal Evolutionary Patterns Resembling Those Observed in Parasitic Plants. International Journal of Molecular Sciences, 20(17), 4107. https://doi.org/10.3390/ijms20174107