Molecular-Level Interactions between Engineered Materials and Cells

 , , and
, , and 
Abstract
:1. Introduction
2. Engineered Materials
2.1. Physical/Mechanical Properties of Materials—Stiffness and Viscosity
2.2. Physical/Mechanical Properties of Materials—Geometrical Factors Including Shape and Morphology
2.3. Physical/Mechanical Properties of Materials—Electrical and Magnetic Potentials
2.4. Chemical/Biological Functionalization of Materials
3. Cellular Sensing of Engineered Materials
3.1. Sensing Receptors on the Cell Surface—Integrins
3.2. Sensing Receptors on the Cell Surface—Mechanosensitive Channels and GPCRs
4. Cytoplasmic Mechanotransduction
5. Nuclear Mechanotransduction
6. Changes in Physical and Chemical States of Nuclear Genomes
7. Changes in Gene Expression
8. Conclusions
Funding
Conflicts of Interest
References
- Park, I.H.; Lerou, P.H.; Zhao, R.; Huo, H.; Daley, G.Q. Generation of human-induced pluripotent stem cells. Nat. Protoc. 2008, 3, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Conaway, L.; Rutherford Bethard, J.; Al-Ayoubi, A.M.; Thompson Bradley, A.; Zheng, H.; Weed, S.A.; Eblen, S.T. Phosphorylation of the alternative mRNA splicing factor 45 (SPF45) by Clk1 regulates its splice site utilization, cell migration and invasion. Nucleic Acids Res. 2013, 41, 4949–4962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.; Zhang, H.; Chen, Z.; Yang, Z.; Shi, D.; Liu, T.; Chen, W.; Yao, F.; Su, X.; Deng, W.; et al. TFAP2B overexpression contributes to tumor growth and progression of thyroid cancer through the COX-2 signaling pathway. Cell Death Dis. 2019, 10, 397. [Google Scholar] [CrossRef] [PubMed]
- Cullot, G.; Boutin, J.; Toutain, J.; Prat, F.; Pennamen, P.; Rooryck, C.; Teichmann, M.; Rousseau, E.; Lamrissi-Garcia, I.; Guyonnet-Duperat, V.; et al. CRISPR-Cas9 genome editing induces megabase-scale chromosomal truncations. Nat. Commun. 2019, 10, 1136. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.H.; Liu, G.Y.; Wang, R.; Moon, S.H.; Gross, R.W.; Patti, G.J. Identifying off-target effects of etomoxir reveals that carnitine palmitoyltransferase I is essential for cancer cell proliferation independent of beta-oxidation. PLoS Biol. 2018, 16, e2003782. [Google Scholar] [CrossRef]
- Hacein-Bey-Abina, S.; Von Kalle, C.; Schmidt, M.; McCormack, M.P.; Wulffraat, N.; Leboulch, P.; Lim, A.; Osborne, C.S.; Pawliuk, R.; Morillon, E.; et al. LMO2-associated clonal T cell proliferation in two patients after gene therapy for SCID-X1. Science 2003, 302, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, D.H.; Kim, H.N.; Wang, C.J.; Kwak, M.K.; Hur, E.; Suh, K.Y.; An, S.S.; Levchenko, A. Directed migration of cancer cells guided by the graded texture of the underlying matrix. Nat. Mater. 2016, 15, 792–801. [Google Scholar] [CrossRef] [Green Version]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef]
- Jang, Y.H.; Park, Y.S.; Nam, J.S.; Yang, Y.; Lee, J.E.; Lee, K.H.; Kang, M.; Chialastri, A.; Noh, H.; Park, J.; et al. Nanotopography-based engineering of retroviral DNA integration patterns. Nanoscale 2019, 11, 5693–5704. [Google Scholar] [CrossRef]
- Zhang, C.; Xie, B.; Zou, Y.; Zhu, D.; Lei, L.; Zhao, D.; Nie, H. Zero-dimensional, one-dimensional, two-dimensional and three-dimensional biomaterials for cell fate regulation. Adv. Drug Deliv. Rev. 2018, 132, 33–56. [Google Scholar] [CrossRef]
- Xi, W.; Saw, T.B.; Delacour, D.; Lim, C.T.; Ladoux, B. Material approaches to active tissue mechanics. Nat. Rev. Mater. 2019, 4, 23–44. [Google Scholar] [CrossRef]
- Muhamed, I.; Chowdhury, F.; Maruthamuthu, V. Biophysical Tools to Study Cellular Mechanotransduction. Bioengineering 2017, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.; Kumar, S.; Schaffer, D. Novel biomaterials to study neural stem cell mechanobiology and improve cell-replacement therapies. Curr. Opin. Biomed. Eng. 2017, 4, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Vining, K.H.; Mooney, D.J. Mechanical forces direct stem cell behaviour in development and regeneration. Nat. Rev. Mol. Cell Biol. 2017, 18, 728–742. [Google Scholar] [CrossRef] [PubMed]
- Di Cio, S.; Gautrot, J.E. Cell sensing of physical properties at the nanoscale: Mechanisms and control of cell adhesion and phenotype. Acta Biomater. 2016, 30, 26–48. [Google Scholar] [CrossRef] [PubMed]
- Charras, G.; Yap, A.S. Tensile Forces and Mechanotransduction at Cell-Cell Junctions. Curr. Biol. 2018, 28, R445–R457. [Google Scholar] [CrossRef] [PubMed]
- Ingber, D.E. Mechanosensation through integrins: Cells act locally but think globally. Proc. Natl. Acad. Sci. USA 2003, 100, 1472–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Q.; Yates, K.; Vogt, C.; Qian, Z.; Frost, M.C.; Zhao, F. Increasing mechanical strength of gelatin hydrogels by divalent metal ion removal. Sci. Rep. 2014, 4, 4706. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Oh, S.; Chae, S.; Lee, J.W.; Choi, K.H.; Lee, K.E.; Chang, J.; Shi, L.; Choi, J.Y.; Lee, J.H. Exceptional Mechanical Properties of Phase-Separation-Free Mo3Se3(-)-Chain-Reinforced Hydrogel Prepared by Polymer Wrapping Process. Nano Lett. 2019, 19, 5717–5724. [Google Scholar] [CrossRef]
- Stutchbury, B.; Atherton, P.; Tsang, R.; Wang, D.Y.; Ballestrem, C. Distinct focal adhesion protein modules control different aspects of mechanotransduction. J. Cell Sci. 2017, 130, 1612–1624. [Google Scholar] [CrossRef] [Green Version]
- Patel, N.R.; Whitehead, A.K.; Newman, J.J.; Caldorera-Moore, M.E. Poly (ethylene glycol) Hydrogels with Tailorable Surface and Mechanical Properties for Tissue Engineering Applications. ACS Biomater. Sci. Eng. 2017, 3, 1494–1498. [Google Scholar] [CrossRef]
- Whitehead, A.K.; Barnett, H.H.; Caldorera-Moore, M.E.; Newman, J.J. Poly (ethylene glycol) hydrogel elasticity influences human mesenchymal stem cell behavior. Regen. Biomater. 2018, 5, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Hendrikson, W.J.; Rouwkema, J.; van Blitterswijk, C.A.; Moroni, L. Influence of PCL molecular weight on mesenchymal stromal cell differentiation. RSC Adv. 2015, 5, 54510–54516. [Google Scholar] [CrossRef] [Green Version]
- Palchesko, R.N.; Zhang, L.; Sun, Y.; Feinberg, A.W. Development of polydimethylsiloxane substrates with tunable elastic modulus to study cell mechanobiology in muscle and nerve. PLoS ONE 2012, 7, e51499. [Google Scholar] [CrossRef] [PubMed]
- Pedraza, E.; Brady, A.C.; Fraker, C.A.; Stabler, C.L. Synthesis of macroporous poly (dimethylsiloxane) scaffolds for tissue engineering applications. J. Biomater. Sci. Polym. Ed. 2013, 24, 1041–1056. [Google Scholar] [CrossRef]
- Varshney, N.; Sahi, A.K.; Vajanthri, K.Y.; Poddar, S.; Balavigneswaran, C.K.; Prabhakar, A.; Rao, V.; Mahto, S.K. Culturing melanocytes and fibroblasts within three-dimensional macroporous PDMS scaffolds: Towards skin dressing material. Cytotechnology 2019, 71, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, K.; Gu, X.; Leong, K.W. Biophysical Regulation of Cell Behavior-Cross Talk between Substrate Stiffness and Nanotopography. Engineering 2017, 3, 36–54. [Google Scholar] [CrossRef]
- Lanniel, M.; Huq, E.; Allen, S.; Buttery, L.; Williams, P.M.; Alexander, M.R. Substrate induced differentiation of human mesenchymal stem cells on hydrogels with modified surface chemistry and controlled modulus. Soft Matter 2011, 7, 6501–6514. [Google Scholar] [CrossRef]
- Moeller, J.; Denisin, A.K.; Sim, J.Y.; Wilson, R.E.; Ribeiro, A.J.S.; Pruitt, B.L. Controlling cell shape on hydrogels using lift-off protein patterning. PLoS ONE 2018, 13, e0189901. [Google Scholar] [CrossRef]
- Wei, S.C.; Fattet, L.; Tsai, J.H.; Guo, Y.R.; Pai, V.H.; Majeski, H.E.; Chen, A.C.; Sah, R.L.; Taylor, S.S.; Engler, A.J.; et al. Matrix stiffness drives epithelial mesenchymal transition and tumour metastasis through a TWIST1-G3BP2 mechanotransduction pathway. Nat. Cell Biol. 2015, 17, 678. [Google Scholar] [CrossRef]
- Bennett, M.; Cantini, M.; Reboud, J.; Cooper, J.M.; Roca-Cusachs, P.; Salmeron-Sanchez, M. Molecular clutch drives cell response to surface viscosity. Proc. Natl. Acad. Sci. USA 2018, 115, 1192–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutolf, M.P.; Hubbell, J.A. Synthetic biomaterials as instructive extracellular microenvironments for morphogenesis in tissue engineering. Nat. Biotechnol. 2005, 23, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Cipitria, A.; Salmeron-Sanchez, M. Mechanotransduction and Growth Factor Signalling to Engineer Cellular Microenvironments. Adv. Healthc. Mater. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Chae, S.; Oh, S.; Kim, S.H.; Choi, K.H.; Meeseepong, M.; Chang, J.; Kim, N.; Kim, Y.H.; Lee, N.E.; et al. Single-Chain Atomic Crystals as Extracellular Matrix-Mimicking Material with Exceptional Biocompatibility and Bioactivity. Nano Lett. 2018, 18, 7619–7627. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, J.J.; Zhang, J.; Wang, X.; Kawazoe, N.; Chen, G. Gold nanoparticle size and shape influence on osteogenesis of mesenchymal stem cells. Nanoscale 2016, 8, 7992–8007. [Google Scholar] [CrossRef] [PubMed]
- Dam, D.H.; Lee, J.H.; Sisco, P.N.; Co, D.T.; Zhang, M.; Wasielewski, M.R.; Odom, T.W. Direct observation of nanoparticle-cancer cell nucleus interactions. ACS Nano 2012, 6, 3318–3326. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.M.; Jean, R.P.; Tan, J.L.; Liu, W.F.; Sniadecki, N.J.; Spector, A.A.; Chen, C.S. Emergent patterns of growth controlled by multicellular form and mechanics. Proc. Natl. Acad. Sci. USA 2005, 102, 11594–11599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altomare, L.; Gadegaard, N.; Visai, L.; Tanzi, M.C.; Fare, S. Biodegradable microgrooved polymeric surfaces obtained by photolithography for skeletal muscle cell orientation and myotube development. Acta Biomater. 2010, 6, 1948–1957. [Google Scholar] [CrossRef]
- Kang, D.H.; Kim, D.; Wang, S.; Song, D.; Yoon, M.H. Water-insoluble, nanocrystalline, and hydrogel fibrillar scaffolds for biomedical applications. Polym. J. 2018, 50, 637–647. [Google Scholar] [CrossRef]
- Sarker, B.; Walter, C.; Pathak, A. Direct Micropatterning of Extracellular Matrix Proteins on Functionalized Polyacrylamide Hydrogels Shows Geometric Regulation of Cell-Cell Junctions. Acs Biomater. Sci. Eng. 2018, 4, 2340–2349. [Google Scholar] [CrossRef]
- Li, J.J.; Kawazoe, N.; Chen, G. Gold nanoparticles with different charge and moiety induce differential cell response on mesenchymal stem cell osteogenesis. Biomaterials 2015, 54, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Gautrot, J.E.; Malmstrom, J.; Sundh, M.; Margadant, C.; Sonnenberg, A.; Sutherland, D.S. The nanoscale geometrical maturation of focal adhesions controls stem cell differentiation and mechanotransduction. Nano Lett. 2014, 14, 3945–3952. [Google Scholar] [CrossRef] [PubMed]
- Werner, M.; Blanquer, S.B.; Haimi, S.P.; Korus, G.; Dunlop, J.W.; Duda, G.N.; Grijpma, D.W.; Petersen, A. Surface Curvature Differentially Regulates Stem Cell Migration and Differentiation via Altered Attachment Morphology and Nuclear Deformation. Adv. Sci. 2017, 4, 1600347. [Google Scholar] [CrossRef] [PubMed]
- Di Cio, S.; Boggild, T.M.L.; Connelly, J.; Sutherland, D.S.; Gautrot, J.E. Differential integrin expression regulates cell sensing of the matrix nanoscale geometry. Acta Biomater. 2017, 50, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Poudineh, M.; Wang, Z.; Labib, M.; Ahmadi, M.; Zhang, L.; Das, J.; Ahmed, S.; Angers, S.; Kelley, S.O. Three-Dimensional Nanostructured Architectures Enable Efficient Neural Differentiation of Mesenchymal Stem Cells via Mechanotransduction. Nano Lett. 2018, 18, 7188–7193. [Google Scholar] [CrossRef] [PubMed]
- Rostam, H.M.; Singh, S.; Vrana, N.E.; Alexander, M.R.; Ghaemmaghami, A.M. Impact of surface chemistry and topography on the function of antigen presenting cells. Biomater. Sci. 2015, 3, 424–441. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Peng, R.; Ding, J. Cell-material interactions revealed via material techniques of surface patterning. Adv. Mater. 2013, 25, 5257–5286. [Google Scholar] [CrossRef] [PubMed]
- Parandakh, A.; Anbarlou, A.; Tafazzoli-Shadpour, M.; Ardeshirylajimi, A.; Khani, M.M. Substrate topography interacts with substrate stiffness and culture time to regulate mechanical properties and smooth muscle differentiation of mesenchymal stem cells. Colloids Surf. B 2019, 173, 194–201. [Google Scholar] [CrossRef]
- Wang, K.; Bruce, A.; Mezan, R.; Kadiyala, A.; Wang, L.; Dawson, J.; Rojanasakul, Y.; Yang, Y. Nanotopographical Modulation of Cell Function through Nuclear Deformation. ACS Appl. Mater. Interfaces 2016, 8, 5082–5092. [Google Scholar] [CrossRef] [Green Version]
- Pitrez, P.R.; Estronca, L.; Vazao, H.; Egesipe, A.L.; Le Corf, A.; Navarro, C.; Levy, N.; De Sandre-Giovannoli, A.; Nissan, X.; Ferreira, L. Substrate Topography Modulates Cell Aging on a Progeria Cell Model. ACS Biomater. Sci. Eng. 2018, 4, 1498–1504. [Google Scholar] [CrossRef]
- Dobbenga, S.; Fratila-Apachitei, L.E.; Zadpoor, A.A. Nanopattern-induced osteogenic differentiation of stem cells—A systematic review. Acta Biomater. 2016, 46, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.S.; Kim, D.S.; Suhito, I.R.; Lee, W.; Song, I.; Kim, T.H. Two-dimensional material-based bionano platforms to control mesenchymal stem cell differentiation. Biomater. Res. 2018, 22, 10. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Li, S.; Le, W. Nanomaterials modulate stem cell differentiation: Biological interaction and underlying mechanisms. J. Nanobiotechnol. 2017, 15, 75. [Google Scholar] [CrossRef] [PubMed]
- Sjostrom, T.; Dalby, M.J.; Hart, A.; Tare, R.; Oreffo, R.O.; Su, B. Fabrication of pillar-like titania nanostructures on titanium and their interactions with human skeletal stem cells. Acta Biomater. 2009, 5, 1433–1441. [Google Scholar] [CrossRef] [PubMed]
- Damodaran, V.B.; Bhatnagar, D.; Leszczak, V.; Popat, K.C. Titania nanostructures: A biomedical perspective. RSC Adv. 2015, 5, 37149–37171. [Google Scholar] [CrossRef]
- Ferra-Canellas, M.D.M.; Llopis-Grimalt, M.A.; Monjo, M.; Ramis, J.M. Tuning Nanopore Diameter of Titanium Surfaces to Improve Human Gingival Fibroblast Response. Int. J. Mol. Sci. 2018, 19, 2881. [Google Scholar] [CrossRef]
- Kulkarni, M.; Mazare, A.; Gongadze, E.; Perutkova, S.; Kralj-Iglic, V.; Milosev, I.; Schmuki, P.; Iglic, A.; Mozetic, M. Titanium nanostructures for biomedical applications. Nanotechnology 2015, 26, 062002. [Google Scholar] [CrossRef]
- Li, W.; Yan, Z.; Ren, J.; Qu, X. Manipulating cell fate: Dynamic control of cell behaviors on functional platforms. Chem. Soc. Rev. 2018, 47, 8639–8684. [Google Scholar] [CrossRef]
- Wei, Y.; Mo, X.; Zhang, P.; Li, Y.; Liao, J.; Li, Y.; Zhang, J.; Ning, C.; Wang, S.; Deng, X.; et al. Directing Stem Cell Differentiation via Electrochemical Reversible Switching between Nanotubes and Nanotips of Polypyrrole Array. ACS Nano 2017, 11, 5915–5924. [Google Scholar] [CrossRef]
- Wong, D.S.; Li, J.; Yan, X.; Wang, B.; Li, R.; Zhang, L.; Bian, L. Magnetically Tuning Tether Mobility of Integrin Ligand Regulates Adhesion, Spreading, and Differentiation of Stem Cells. Nano Lett. 2017, 17, 1685–1695. [Google Scholar] [CrossRef]
- Wu, C.; Shen, Y.; Chen, M.; Wang, K.; Li, Y.; Cheng, Y. Recent Advances in Magnetic-Nanomaterial-Based Mechanotransduction for Cell Fate Regulation. Adv. Mater. 2018, 30, e1705673. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Sun, J.; Zhao, L.; Zhang, F.; Liang, X.J.; Guo, Y.; Weir, M.D.; Reynolds, M.A.; Gu, N.; Xu, H.H.K. Magnetic field and nano-scaffolds with stem cells to enhance bone regeneration. Biomaterials 2018, 183, 151–170. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zheng, X.; Ahmed, W.; Yao, Y.; Bai, J.; Chen, Y.; Gao, C. Design and Applications of Cell-Selective Surfaces and Interfaces. Biomacromolecules 2018, 19, 1746–1763. [Google Scholar] [CrossRef] [PubMed]
- Engin, A.B.; Nikitovic, D.; Neagu, M.; Henrich-Noack, P.; Docea, A.O.; Shtilman, M.I.; Golokhvast, K.; Tsatsakis, A.M. Mechanistic understanding of nanoparticles’ interactions with extracellular matrix: The cell and immune system. Part. Fibre Toxicol. 2017, 14, 22. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Magli, A.; Anugrah, Y.; Koester, S.J.; Perlingeiro, R.C.R.; Shen, W. Nanotopography-responsive myotube alignment and orientation as a sensitive phenotypic biomarker for Duchenne Muscular Dystrophy. Biomaterials 2018, 183, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Koo, S.; Muhammad, R.; Peh, G.S.; Mehta, J.S.; Yim, E.K. Micro-and nanotopography with extracellular matrix coating modulate human corneal endothelial cell behavior. Acta Biomater. 2014, 10, 1975–1984. [Google Scholar] [CrossRef] [PubMed]
- Hackett, A.J.; Malmstrom, J.; Travas-Sejdic, J. Functionalization of conducting polymers for biointerface applications. Prog. Polym. Sci. 2017, 70, 18–33. [Google Scholar] [CrossRef]
- Hoshiba, T.; Tanaka, M. Integrin-independent Cell Adhesion Substrates: Possibility of Applications for Mechanobiology Research. Anal. Sci. 2016, 32, 1151–1158. [Google Scholar] [CrossRef] [Green Version]
- Jedrzejczak-Silicka, M.; Trukawka, M.; Dudziak, M.; Piotrowska, K.; Mijowska, E. Hexagonal Boron Nitride Functionalized with Au Nanoparticles-Properties and Potential Biological Applications. Nanomaterials 2018, 8, 605. [Google Scholar] [CrossRef]
- Liu, X.Q.; Tang, R.Z. Biological responses to nanomaterials: Understanding nano-bio effects on cell behaviors. Drug Deliv. 2017, 24, 1–15. [Google Scholar] [CrossRef]
- Finke, B.; Luethen, F.; Schroeder, K.; Mueller, P.D.; Bergemann, C.; Frant, M.; Ohl, A.; Nebe, B.J. The effect of positively charged plasma polymerization on initial osteoblastic focal adhesion on titanium surfaces. Biomaterials 2007, 28, 4521–4534. [Google Scholar] [CrossRef] [PubMed]
- Keselowsky, B.G.; Collard, D.M.; Garcia, A.J. Integrin binding specificity regulates biomaterial surface chemistry effects on cell differentiation. Proc. Natl. Acad. Sci. USA 2005, 102, 5953–5957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Zhang, X.; Guo, Y.; Meng, F.; Sachs, F.; Guo, J. Mechanical dynamics in live cells and fluorescence-based force/tension sensors. Biochim. Biophys. Acta 2015, 1853, 1889–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sicard, R.E. Hormones, neurosecretions, and growth factors as signal molecules for intercellular communication. Dev. Comp. Immunol. 1986, 10, 269–272. [Google Scholar] [CrossRef]
- Uings, I.J.; Farrow, S.N. Cell receptors and cell signalling. Mol. Pathol. 2000, 53, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. Integrins: Versatility, modulation, and signaling in cell adhesion. Cell 1992, 69, 11–25. [Google Scholar] [CrossRef]
- Takada, Y.; Ye, X.; Simon, S. The integrins. Genome Biol. 2007, 8, 215. [Google Scholar] [CrossRef]
- Cooke, M.J.; Phillips, S.R.; Shah, D.S.; Athey, D.; Lakey, J.H.; Przyborski, S.A. Enhanced cell attachment using a novel cell culture surface presenting functional domains from extracellular matrix proteins. Cytotechnology 2008, 56, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Xiao, T.; Takagi, J.; Coller, B.S.; Wang, J.H.; Springer, T.A. Structural basis for allostery in integrins and binding to fibrinogen-mimetic therapeutics. Nature 2004, 432, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Guo, S.S.; Fassler, R. Integrin-mediated mechanotransduction. J. Cell Biol. 2016, 215, 445–456. [Google Scholar] [CrossRef]
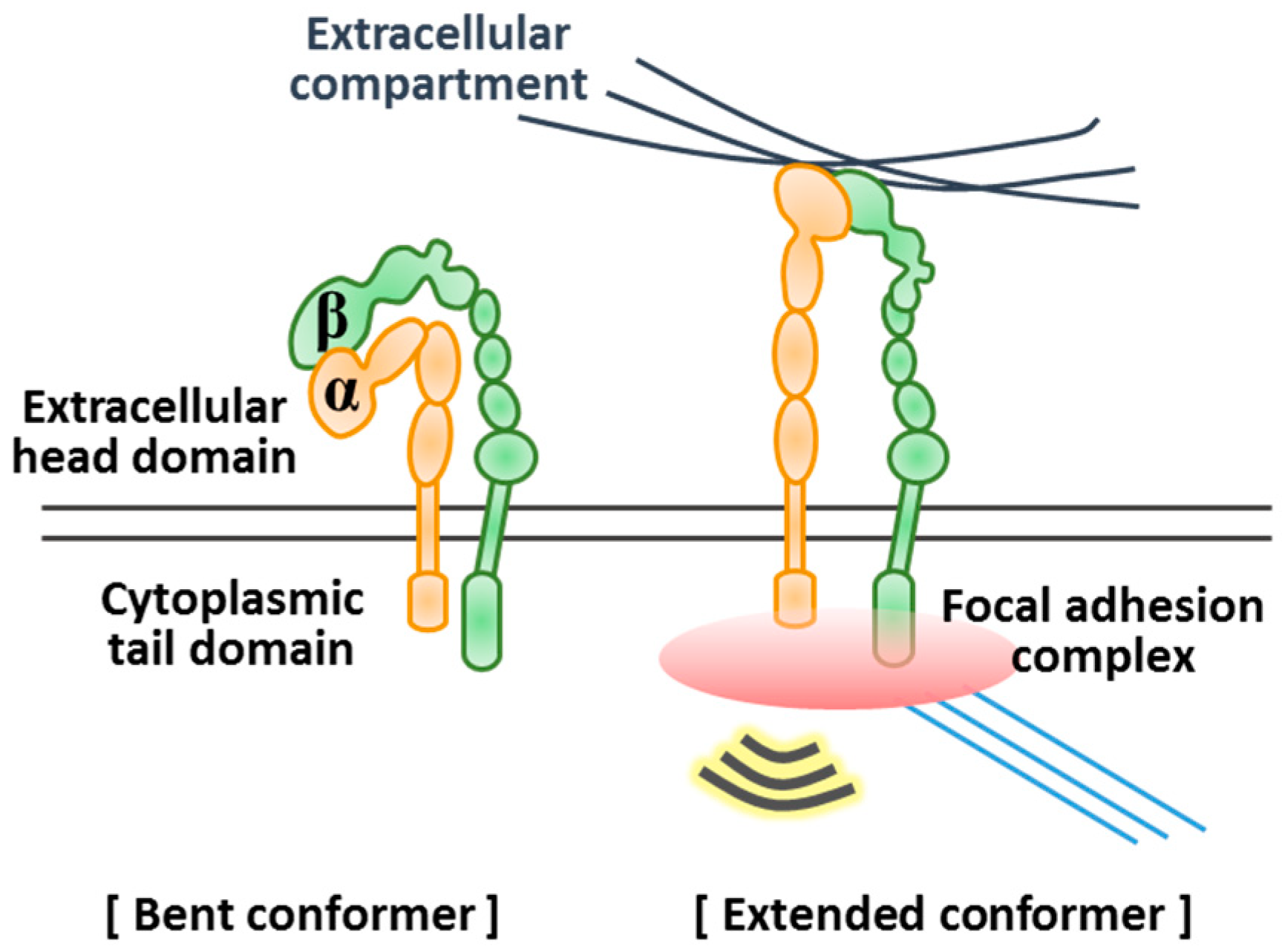
- Askari, J.A.; Buckley, P.A.; Mould, A.P.; Humphries, M.J. Linking integrin conformation to function. J Cell Sci 2009, 122, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphries, J.D.; Byron, A.; Humphries, M.J. Integrin ligands at a glance. J. Cell Sci. 2006, 119, 3901–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, R.P.; Chiquet-Ehrismann, R. Tenascin-C: Its functions as an integrin ligand. Int. J. Biochem. Cell B 2015, 65, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Van der Vieren, M.; Le Trong, H.; Wood, C.L.; Moore, P.F.; St John, T.; Staunton, D.E.; Gallatin, W.M. A novel leukointegrin, alpha d beta 2, binds preferentially to ICAM-3. Immunity 1995, 3, 683–690. [Google Scholar] [CrossRef]
- Yeh, Y.C.; Ling, J.Y.; Chen, W.C.; Lin, H.H.; Tang, M.J. Mechanotransduction of matrix stiffness in regulation of focal adhesion size and number: Reciprocal regulation of caveolin-1 and beta1 integrin. Sci. Rep. 2017, 7, 15008. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.Y.; Dreiss, A.D.; Zhou, Z.; Hansen, J.C.; Siedlecki, C.A.; Hengstebeck, R.W.; Cheng, J.; Winograd, N.; Donahue, H.J. The regulation of integrin-mediated osteoblast focal adhesion and focal adhesion kinase expression by nanoscale topography. Biomaterials 2007, 28, 1787–1797. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, S. Calcium integrin-binding protein activates platelet integrin alpha IIbbeta 3. J. Biol. Chem. 2002, 277, 1919–1923. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Li, Z.; Parks, W.M.; Dumbauld, D.W.; Garcia, A.J.; Mould, A.P.; Humphries, M.J.; Zhu, C. Cyclic mechanical reinforcement of integrin-ligand interactions. Mol. Cell 2013, 49, 1060–1068. [Google Scholar] [CrossRef]
- Sukharev, S.; Sachs, F. Molecular force transduction by ion channels: Diversity and unifying principles. J. Cell Sci. 2012, 125, 3075–3083. [Google Scholar] [CrossRef]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef]
- Cox, C.D.; Bae, C.; Ziegler, L.; Hartley, S.; Nikolova-Krstevski, V.; Rohde, P.R.; Ng, C.A.; Sachs, F.; Gottlieb, P.A.; Martinac, B. Removal of the mechanoprotective influence of the cytoskeleton reveals PIEZO1 is gated by bilayer tension. Nat. Commun. 2016, 7, 10366. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hou, B.; Tumova, S.; Muraki, K.; Bruns, A.; Ludlow, M.J.; Sedo, A.; Hyman, A.J.; McKeown, L.; Young, R.S.; et al. Piezo1 integration of vascular architecture with physiological force. Nature 2014, 515, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Ranade, S.S.; Woo, S.H.; Dubin, A.E.; Moshourab, R.A.; Wetzel, C.; Petrus, M.; Mathur, J.; Begay, V.; Coste, B.; Mainquist, J.; et al. Piezo2 is the major transducer of mechanical forces for touch sensation in mice. Nature 2014, 516, 121–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Servin-Vences, M.R.; Moroni, M.; Lewin, G.R.; Poole, K. Direct measurement of TRPV4 and PIEZO1 activity reveals multiple mechanotransduction pathways in chondrocytes. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Chalfie, M. Neurosensory mechanotransduction. Nat. Rev. Mol. Cell Biol. 2009, 10, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Haswell, E.S.; Phillips, R.; Rees, D.C. Mechanosensitive channels: What can they do and how do they do it? Structure 2011, 19, 1356–1369. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Mathur, J.; Vessieres, E.; Hammack, S.; Nonomura, K.; Favre, J.; Grimaud, L.; Petrus, M.; Francisco, A.; Li, J.; et al. GPR68 Senses Flow and Is Essential for Vascular Physiology. Cell 2018, 173, 762–775. [Google Scholar] [CrossRef] [PubMed]
- Chachisvilis, M.; Zhang, Y.L.; Frangos, J.A. G protein-coupled receptors sense fluid shear stress in endothelial cells. Proc. Natl. Acad. Sci. USA 2006, 103, 15463–15468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, M.G.; Vanek, M.; Guerini, D.; Gasser, J.A.; Jones, C.E.; Junker, U.; Hofstetter, H.; Wolf, R.M.; Seuwen, K. Proton-sensing G-protein-coupled receptors. Nature 2003, 425, 93–98. [Google Scholar] [CrossRef]
- Mederos y Schnitzler, M.; Storch, U.; Meibers, S.; Nurwakagari, P.; Breit, A.; Essin, K.; Gollasch, M.; Gudermann, T. Gq-coupled receptors as mechanosensors mediating myogenic vasoconstriction. EMBO J. 2008, 27, 3092–3103. [Google Scholar] [CrossRef]
- Maniotis, A.J.; Chen, C.S.; Ingber, D.E. Demonstration of mechanical connections between integrins cytoskeletal filaments, and nucleoplasm that stabilize nuclear structure. Proc. Natl. Acad. Sci. USA 1997, 94, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Giancotti, F.G.; Ruoslahti, E. Integrin signaling. Science 1999, 285, 1028–1032. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Teramoto, H.; Coso, O.A.; Gutkind, J.S.; Burbelo, P.D.; Akiyama, S.K.; Yamada, K.M. Integrin function: Molecular hierarchies of cytoskeletal and signaling molecules. J. Cell Biol. 1995, 131, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Levental, K.R.; Yu, H.; Kass, L.; Lakins, J.N.; Egeblad, M.; Erler, J.T.; Fong, S.F.; Csiszar, K.; Giaccia, A.; Weninger, W.; et al. Matrix crosslinking forces tumor progression by enhancing integrin signaling. Cell 2009, 139, 891–906. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Luo, B.H. Integrin bi-directional signaling across the plasma membrane. J. Cell Physiol. 2013, 228, 306–312. [Google Scholar] [CrossRef]
- Turner, C.E. Paxillin and focal adhesion signalling. Nat. Cell Biol. 2000, 2, E231–E236. [Google Scholar] [CrossRef] [PubMed]
- Haynie, D.T. Molecular physiology of the tensin brotherhood of integrin adaptor proteins. Proteins 2014, 82, 1113–1127. [Google Scholar] [CrossRef]
- Otey, C.A.; Carpen, O. Alpha-actinin revisited: A fresh look at an old player. Cell Motil. Cytoskelet. 2004, 58, 104–111. [Google Scholar] [CrossRef]
- Montanez, E.; Ussar, S.; Schifferer, M.; Bosl, M.; Zent, R.; Moser, M.; Fassler, R. Kindlin-2 controls bidirectional signaling of integrins. Genes Dev. 2008, 22, 1325–1330. [Google Scholar] [CrossRef] [Green Version]
- Ron, A.; Azeloglu, E.U.; Calizo, R.C.; Hu, M.; Bhattacharya, S.; Chen, Y.; Jayaraman, G.; Lee, S.; Neves-Zaph, S.R.; Li, H.; et al. Cell shape information is transduced through tension-independent mechanisms. Nat. Commun. 2017, 8, 2145. [Google Scholar] [CrossRef]
- Yamashita, H.; Ichikawa, T.; Matsuyama, D.; Kimura, Y.; Ueda, K.; Craig, S.W.; Harada, I.; Kioka, N. The role of the interaction of the vinculin proline-rich linker region with vinexin alpha in sensing the stiffness of the extracellular matrix. J. Cell Sci. 2014, 127, 1875–1886. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Sharif-Naeini, R.; Folgering, J.R.; Bichet, D.; Duprat, F.; Honore, E. Canonical TRP channels and mechanotransduction: From physiology to disease states. Pflug. Arch. 2010, 460, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of yap/taz in mechanotransduction. Nature 2011, 474, 179. [Google Scholar] [CrossRef] [PubMed]
- Uhler, C.; Shivashankar, G.V. Regulation of genome organization and gene expression by nuclear mechanotransduction. Nat. Rev. Mol. Cell Biol. 2017, 18, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Guilluy, C.; Burridge, K. Nuclear mechanotransduction: Forcing the nucleus to respond. Nucleus 2015, 6, 19–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosa, B.A.; Rothballer, A.; Kutay, U.; Schwartz, T.U. LINC complexes form by binding of three KASH peptides to domain interfaces of trimeric SUN proteins. Cell 2012, 149, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Panciera, T.; Azzolin, L.; Cordenonsi, M.; Piccolo, S. Mechanobiology of YAP and TAZ in physiology and disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 758–770. [Google Scholar] [CrossRef]
- Lanctot, C.; Cheutin, T.; Cremer, M.; Cavalli, G.; Cremer, T. Dynamic genome architecture in the nuclear space: Regulation of gene expression in three dimensions. Nat. Rev. Genet. 2007, 8, 104–115. [Google Scholar] [CrossRef]
- Dekker, J.; Mirny, L. The 3D Genome as Moderator of Chromosomal Communication. Cell 2016, 164, 1110–1121. [Google Scholar] [CrossRef] [Green Version]
- Poh, Y.C.; Shevtsov, S.P.; Chowdhury, F.; Wu, D.C.; Na, S.; Dundr, M.; Wang, N. Dynamic force-induced direct dissociation of protein complexes in a nuclear body in living cells. Nat. Commun. 2012, 3, 866. [Google Scholar] [CrossRef]
- Pajerowski, J.D.; Dahl, K.N.; Zhong, F.L.; Sammak, P.J.; Discher, D.E. Physical plasticity of the nucleus in stem cell differentiation. Proc. Natl. Acad. Sci. USA 2007, 104, 15619–15624. [Google Scholar] [CrossRef] [PubMed]
- Swift, J.; Ivanovska, I.L.; Buxboim, A.; Harada, T.; Dingal, P.C.; Pinter, J.; Pajerowski, J.D.; Spinler, K.R.; Shin, J.W.; Tewari, M.; et al. Nuclear lamin—A scales with tissue stiffness and enhances matrix-directed differentiation. Science 2013, 341, 1240104. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.Y.; Jaalouk, D.E.; Vartiainen, M.K.; Lammerding, J. Lamin A/C and emerin regulate MKL1-SRF activity by modulating actin dynamics. Nature 2013, 497, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Tajik, A.; Zhang, Y.; Wei, F.; Sun, J.; Jia, Q.; Zhou, W.; Singh, R.; Khanna, N.; Belmont, A.S.; Wang, N. Transcription upregulation via force-induced direct stretching of chromatin. Nat. Mater. 2016, 15, 1287–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.S.; Hale, C.M.; Panorchan, P.; Khatau, S.B.; George, J.P.; Tseng, Y.; Stewart, C.L.; Hodzic, D.; Wirtz, D. Nuclear lamin A/C deficiency induces defects in cell mechanics, polarization, and migration. Biophys. J. 2007, 93, 2542–2552. [Google Scholar] [CrossRef] [PubMed]
- Guilluy, C.; Osborne, L.D.; Van Landeghem, L.; Sharek, L.; Superfine, R.; Garcia-Mata, R.; Burridge, K. Isolated nuclei adapt to force and reveal a mechanotransduction pathway in the nucleus. Nat. Cell Biol. 2014, 16, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Zhou, W.; Yao, W.; Yang, F.; Zhang, S.; Singh, R.; Chen, J.; Chen, J.J.; Zhang, Y.; Wei, F.; et al. Downregulation of YAP-dependent Nupr1 promotes tumor-repopulating cell growth in soft matrices. Oncogenesis 2016, 5, e220. [Google Scholar] [CrossRef]
- Jaalouk, D.E.; Lammerding, J. Mechanotransduction gone awry. Nat. Rev. Mol. Cell Biol. 2009, 10, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, M.A.; Chen, C.S. Mechanotransduction in development: A growing role for contractility. Nat. Rev. Mol. Cell Biol. 2009, 10, 34–43. [Google Scholar] [CrossRef]
- Discher, D.E.; Mooney, D.J.; Zandstra, P.W. Growth factors, matrices, and forces combine and control stem cells. Science 2009, 324, 1673–1677. [Google Scholar] [CrossRef]
- Folkman, J.; Moscona, A. Role of cell shape in growth control. Nature 1978, 273, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Bruce, M.; Spiegelman, C.A.G. Fibronectin modulation of cell shape and lipogenic gene expression in 3T3-adipocytes. Cell 1983, 35, 657–666. [Google Scholar] [CrossRef]
- Singhvi, R.; Kumar, A.; Lopez, G.P.; Stephanopoulos, G.N.; Wang, D.I.; Whitesides, G.M.; Ingber, D.E. Engineering cell shape and function. Science 1994, 264, 696–698. [Google Scholar] [CrossRef] [PubMed]
- McBeath, R.; Pirone, D.M.; Nelson, C.M.; Bhadriraju, K.; Chen, C.S. Cell shape, cytoskeletal tension, and RhoA regulate stem cell lineage commitment. Dev. Cell 2004, 6, 483–495. [Google Scholar] [CrossRef]
- Wada, K.; Itoga, K.; Okano, T.; Yonemura, S.; Sasaki, H. Hippo pathway regulation by cell morphology and stress fibers. Development 2011, 138, 3907–3914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.; Tajik, A.; Chen, J.; Jia, Q.; Chowdhury, F.; Wang, L.; Chen, J.; Zhang, S.; Hong, Y.; Yi, H.; et al. Matrix softness regulates plasticity of tumour-repopulating cells via H3K9 demethylation and Sox2 expression. Nat. Commun. 2014, 5, 4619. [Google Scholar] [CrossRef] [PubMed]
- Aragona, M.; Panciera, T.; Manfrin, A.; Giulitti, S.; Michielin, F.; Elvassore, N.; Dupont, S.; Piccolo, S. A Mechanical Checkpoint Controls Multicellular Growth through YAP/TAZ Regulation by Actin-Processing Factors. Cell 2013, 154, 1047–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.C.; Yeh, Y.T.; Nguyen, P.; Limqueco, E.; Lopez, J.; Thorossian, S.; Guan, K.L.; Li, Y.S.J.; Chien, S. Flow-dependent YAP/TAZ activities regulate endothelial phenotypes and atherosclerosis. Proc. Natl. Acad. Sci. USA 2016, 113, 11525–11530. [Google Scholar] [CrossRef] [Green Version]
- Schlegelmilch, K.; Mohseni, M.; Kirak, O.; Pruszak, J.; Rodriguez, J.R.; Zhou, D.W.; Kreger, B.T.; Vasioukhin, V.; Avruch, J.; Brummelkamp, T.R.; et al. Yap1 Acts Downstream of alpha-Catenin to Control Epidermal Proliferation. Cell 2011, 144, 782–795. [Google Scholar] [CrossRef]
- Hong, J.H.; Hwang, E.S.; McManus, M.T.; Amsterdam, A.; Tian, Y.; Kalmukova, R.; Mueller, E.; Benjamin, T.; Spiegelman, B.M.; Sharp, P.A.; et al. TAZ, a transcriptional modulator of mesenchymal stem cell differentiation. Science 2005, 309, 1074–1078. [Google Scholar] [CrossRef]
- Kuroda, M.; Wada, H.; Kimura, Y.; Ueda, K.; Kioka, N. Vinculin promotes nuclear localization of TAZ to inhibit ECM stiffness-dependent differentiation into adipocytes. J. Cell Sci. 2017, 130, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Rowe, R.G.; Botvinick, E.L.; Kurup, A.; Putnam, A.J.; Seiki, M.; Weaver, V.M.; Keller, E.T.; Goldstein, S.; Dai, J.L.; et al. MT1-MMP-Dependent Control of Skeletal Stem Cell Commitment via a beta 1-Integrin/YAP/TAZ Signaling Axis. Dev. Cell 2013, 25, 402–416. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.L.; Tian, K.; Zheng, X.F.; Li, L.N.; Zhang, W.G.; Wang, S.Y.; Qin, J.H. Mesenchymal Stem Cell and Chondrocyte Fates in a Multishear Microdevice Are Regulated by Yes-Associated Protein. Stem Cells Dev. 2013, 22, 2083–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, B.; Li, L.; Wang, L.; Wang, C.Y.; Yu, J.D.; Guan, K.L. Cell detachment activates the Hippo pathway via cytoskeleton reorganization to induce anoikis. Genes Dev. 2012, 26, 54–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Cheng, F.L.; Wei, Y.J.; Zhang, L.J.; Guo, D.S.; Wang, B.F.; Li, W. Inhibition of TAZ contributes radiation-induced senescence and growth arrest in glioma cells. Oncogene 2019, 38, 2788–2799. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Wei, X.; Li, W.; Udan, R.S.; Yang, Q.; Kim, J.; Xie, J.; Ikenoue, T.; Yu, J.; Li, L.; et al. Inactivation of YAP oncoprotein by the Hippo pathway is involved in cell contact inhibition and tissue growth control. Genes Dev. 2007, 21, 2747–2761. [Google Scholar] [CrossRef] [Green Version]
- Halder, G.; Dupont, S.; Piccolo, S. Transduction of mechanical and cytoskeletal cues by YAP and TAZ. Nat. Rev. Mol. Cell Biol. 2012, 13, 591–600. [Google Scholar] [CrossRef]
- Haraguchi, T.; Holaska, J.M.; Yamane, M.; Koujin, T.; Hashiguchi, N.; Mori, C.; Wilson, K.L.; Hiraoka, Y. Emerin binding to Btf, a death-promoting transcriptional repressor, is disrupted by a missense mutation that causes Emery-Dreifuss muscular dystrophy. Eur. J. Biochem. 2004, 271, 1035–1045. [Google Scholar] [CrossRef]
- Dreuillet, C.; Tillit, J.; Kress, M.; Ernoult-Lange, M. In vivo and in vitro interaction between human transcription factor MOK2 and nuclear lamin A/C. Nucleic Acids Res. 2002, 30, 4634–4642. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, F.L.; Holaska, J.M.; Zhang, Z.; Sharma, A.; Manilal, S.; Holt, I.; Stamm, S.; Wilson, K.L.; Morris, G.E. Emerin interacts in vitro with the splicing-associated factor, YT521-B. Eur. J. Biochem. 2003, 270, 2459–2466. [Google Scholar] [CrossRef]
- Stein, G.S.; Lian, J.B.; van Wijnen, A.J.; Stein, J.L.; Javed, A.; Montecino, M.; Choi, J.Y.; Vradii, D.; Zaidi, S.K.; Pratap, J.; et al. Organization of transcriptional regulatory machinery in nuclear microenvironments: Implications for biological control and cancer. Cancer Treat. Rev. 2007, 47, 242–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingber, D.E. The riddle of morphogenesis: A question of solution chemistry or molecular cell engineering? Cell 1993, 75, 1249–1252. [Google Scholar] [CrossRef]
- Wang, N.; Tytell, J.D.; Ingber, D.E. Mechanotransduction at a distance: Mechanically coupling the extracellular matrix with the nucleus. Nat. Rev. Mol. Cell Biol. 2009, 10, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Jain, N.; Iyer, K.V.; Kumar, A.; Shivashankar, G.V. Cell geometric constraints induce modular gene-expression patterns via redistribution of HDAC3 regulated by actomyosin contractility. Proc. Natl. Acad. Sci. USA 2013, 110, 11349–11354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, H.Q.; Ghatak, S.; Yeung, C.Y.C.; Tellkamp, F.; Gunschmann, C.; Dieterich, C.; Yeroslaviz, A.; Habermand, B.; Pombo, A.; Niessen, C.M.; et al. Mechanical regulation of transcription controls Polycomb-mediated gene silencing during lineage commitment. Nat. Cell Biol. 2016, 18, 864. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nagarajan, M.; Uhler, C.; Shivashankar, G.V. Orientation and repositioning of chromosomes correlate with cell geometry-dependent gene expression. Mol. Biol. Cell 2017, 28, 1997–2009. [Google Scholar] [CrossRef]
- Cho, S.; Vashisth, M.; Abbas, A.; Majkut, S.; Vogel, K.; Xia, Y.T.; Ivanovska, I.L.; Irianto, J.; Tewari, M.; Zhu, K.Z.; et al. Mechanosensing by the Lamina Protects against Nuclear Rupture, DNA Damage, and Cell-Cycle Arrest. Dev. Cell 2019, 49, 920–935. [Google Scholar] [CrossRef]
- Ivanovska, I.L.; Swift, J.; Spinler, K.; Dingal, D.; Cho, S.; Discher, D.E. Cross-linked matrix rigidity and soluble retinoids synergize in nuclear lamina regulation of stem cell differentiation. Mol. Biol. Cell 2017, 28, 2010–2022. [Google Scholar] [CrossRef]
- Elosegui-Artola, A.; Andreu, I.; Beedle, A.E.M.; Lezamiz, A.; Uroz, M.; Kosmalska, A.J.; Oria, R.; Kechagia, J.Z.; Rico-Lastres, P.; Le Roux, A.L.; et al. Force Triggers YAP Nuclear Entry by Regulating Transport across Nuclear Pores. Cell 2017, 171, 1397–1410. [Google Scholar] [CrossRef]
- Itano, N.; Okamoto, S.; Zhang, D.X.; Lipton, S.A.; Ruoslahti, E. Cell spreading controls endoplasmic and nuclear calcium: A physical gene regulation pathway from the cell surface to the nucleus. Proc. Natl. Acad. Sci. USA 2003, 100, 5181–5186. [Google Scholar] [CrossRef] [Green Version]
- Enyedi, B.; Jelcic, M.; Niethammer, P. The Cell Nucleus Serves as a Mechanotransducer of Tissue Damage-Induced Inflammation. Cell 2016, 165, 1160–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, J.; Goetze, S.; Heng, H.; Krawetz, S.A.; Benham, C. From DNA structure to gene expression: Mediators of nuclear compartmentalization and dynamics. Chromosome Res. 2003, 11, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Lee, I.W.; Jo, Y.J.; Namgoong, S.; Kim, N.H. Depletion of the LINC complex disrupts cytoskeleton dynamics and meiotic resumption in mouse oocytes. Sci. Rep. 2016, 6, 20408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arsenovic, P.T.; Ramachandran, I.; Bathula, K.; Zhu, R.J.; Narang, J.D.; Noll, N.A.; Lemmon, C.A.; Gundersen, G.G.; Conway, D.E. Nesprin-2G, a Component of the Nuclear LINC Complex, Is Subject to Myosin-Dependent Tension. Biophys. J. 2016, 110, 34–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutscheidt, S.; Zhu, R.; Antoku, S.; Luxton, G.W.; Stagljar, I.; Fackler, O.T.; Gundersen, G.G. FHOD1 interaction with nesprin-2G mediates TAN line formation and nuclear movement. Nat. Cell Biol. 2014, 16, 708–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoku, S.; Zhu, R.; Kutscheidt, S.; Fackler, O.T.; Gundersen, G.G. Reinforcing the LINC complex connection to actin filaments: The role of FHOD1 in TAN line formation and nuclear movement. Cell Cycle 2015, 14, 2200–2205. [Google Scholar] [CrossRef] [PubMed]
- Borrego-Pinto, J.; Jegou, T.; Osorio, D.S.; Aurade, F.; Gorjanaacz, M.; Koch, B.; Mattaj, I.W.; Gomes, E.R. Samp1 is a component of TAN lines and is required for nuclear movement. J. Cell Sci. 2012, 125, 1099–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lammerding, J.; Schulze, P.C.; Takahashi, T.; Kozlov, S.; Sullivan, T.; Kamm, R.D.; Stewart, C.L.; Lee, R.T. Lamin A/C deficiency causes defective nuclear mechanics and mechanotransduction. J. Clin. Investig. 2004, 113, 370–378. [Google Scholar] [CrossRef] [Green Version]
- Ihalainen, T.O.; Aires, L.; Herzog, F.A.; Schwartlander, R.; Moeller, J.; Vogel, V. Differential basal-to-apical accessibility of lamin A/C epitopes in the nuclear lamina regulated by changes in cytoskeletal tension. Nat. Mater. 2015, 14, 1252–1261. [Google Scholar] [CrossRef] [Green Version]
- Zuo, B.; Yang, J.; Wang, F.; Wang, L.; Yin, Y.; Dan, J.; Liu, N.; Liu, L. Influences of lamin A levels on induction of pluripotent stem cells. Biol. Open 2012, 1, 1118–1127. [Google Scholar] [CrossRef] [Green Version]
- Constantinescu, D.; Gray, H.L.; Sammak, P.J.; Schatten, G.P.; Csoka, A.B. Lamin A/C expression is a marker of mouse and human embryonic stem cell differentiation. Stem Cells 2006, 24, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Buxboim, A.; Swift, J.; Irianto, J.; Spinler, K.R.; Dingal, P.C.; Athirasala, A.; Kao, Y.R.; Cho, S.; Harada, T.; Shin, J.W.; et al. Matrix elasticity regulates lamin-A,C phosphorylation and turnover with feedback to actomyosin. Curr. Biol. 2014, 24, 1909–1917. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Irianto, J.; Discher, D.E. Mechanosensing by the nucleus: From pathways to scaling relationships. J. Cell Biol. 2017, 216, 305–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruenbaum, Y.; Margalit, A.; Goldman, R.D.; Shumaker, D.K.; Wilson, K.L. The nuclear lamina comes of age. Nat. Rev. Mol. Cell Biol. 2005, 6, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Worman, H.J. Interaction between an integral protein of the nuclear envelope inner membrane and human chromodomain proteins homologous to Drosophila HP1. J. Biol. Chem. 1996, 271, 14653–14656. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, S.M.; Koo, P.K.; Zhao, Y.; Mochrie, S.G.; King, M.C. The tethering of chromatin to the nuclear envelope supports nuclear mechanics. Nat. Commun. 2015, 6, 7159. [Google Scholar] [CrossRef] [PubMed]
- Finan, J.D.; Guilak, F. The effects of osmotic stress on the structure and function of the cell nucleus. J. Cell Biochem. 2010, 109, 460–467. [Google Scholar] [CrossRef]
- Finan, J.D.; Leddy, H.A.; Guilak, F. Osmotic stress alters chromatin condensation and nucleocytoplasmic transport. Biochem. Biophys. Res. Commun. 2011, 408, 230–235. [Google Scholar] [CrossRef] [Green Version]
- Stephens, A.D.; Banigan, E.J.; Adam, S.A.; Goldman, R.D.; Marko, J.F. Chromatin and lamin A determine two different mechanical response regimes of the cell nucleus. Mol. Biol. Cell 2017, 28, 1984–1996. [Google Scholar] [CrossRef]
- Heo, S.J.; Thorpe, S.D.; Driscoll, T.P.; Duncan, R.L.; Lee, D.A.; Mauck, R.L. Biophysical Regulation of Chromatin Architecture Instills a Mechanical Memory in Mesenchymal Stem Cells. Sci. Rep. 2015, 5, 16895. [Google Scholar] [CrossRef]
- Iyer, K.V.; Pulford, S.; Mogilner, A.; Shivashankar, G.V. Mechanical activation of cells induces chromatin remodeling preceding MKL nuclear transport. Biophys. J. 2012, 103, 1416–1428. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Lee, C.I.; Lin, T.E.; Lim, S.H.; Zhou, J.; Tseng, Y.C.; Chien, S.; Chiu, J.J. Role of histone deacetylases in transcription factor regulation and cell cycle modulation in endothelial cells in response to disturbed flow. Proc. Natl. Acad. Sci. USA 2012, 109, 1967–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnsdorf, E.J.; Tummala, P.; Castillo, A.B.; Zhang, F.; Jacobs, C.R. The epigenetic mechanism of mechanically induced osteogenic differentiation. J. Biomech. 2010, 43, 2881–2886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downing, T.L.; Soto, J.; Morez, C.; Houssin, T.; Fritz, A.; Yuan, F.; Chu, J.; Patel, S.; Schaffer, D.V.; Li, S. Biophysical regulation of epigenetic state and cell reprogramming. Nat. Mater. 2013, 12, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Tibbitt, M.W.; Basta, L.; Anseth, K.S. Mechanical memory and dosing influence stem cell fate. Nat. Mater. 2014, 13, 645–652. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material/Substrates | Properties | Cell Type | Effects | Ref. |
|---|---|---|---|---|
| PAAm gel coated with fibronectin | Stiffness: ~8 kPa and ~100 kPa | NIH3T3 fibroblast cells | The mobilities of the structural proteins are directly influenced by the stiffness of the substrate. The turnover rates of talin, vinculin and tensin1 decreased with increasing ECM stiffness. | [20] |
| PAAm gel coated with collagen | Stiffness: 150 Pa and 5700 Pa | Human MCF10A and mouse Eph4Ras cells | High matrix stiffness promotes nuclear translocation of TWIST1. | [30] |
| PDMS micropatterned with laminin | Stiffness: 5 kPa (soft) and 1.72 MPa (hard) | PC12 rat adrenal pheochromocytoma cells | Soft PDMS resulted in significant increase in neurite length | [24] |
| PDMS micropatterned with fibronectin | Stiffness: 5 kPa, 50 kPa, 130 kPa, 830 kPa, and 1.72 MPa | C2C12 cells | The number of myotube clusters was increased with softer PDMS substrates (5 kPa) | [24] |
| Arg-Gly-Asp (RGD)-functionalized lipid bilayers composed of either fluid-DOPC or gel-DPPC deposited on glass substrate | Viscosity: 8.4 ×10−11 Pa⋅s⋅m (DOPC) 3.0 × 10−9 Pa⋅s⋅m (DPPC) | C2C12 cells | Substrates with low viscosity prevented protein unfolding and increased actin flow | [31] |
| Material/Substrates | Properties | Cell Type | Effects | Ref. |
|---|---|---|---|---|
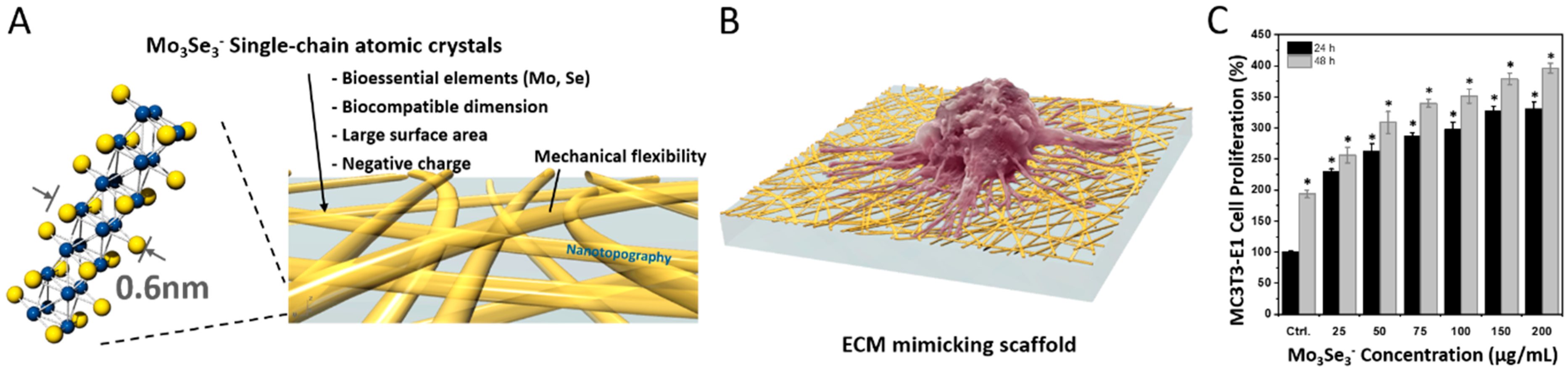
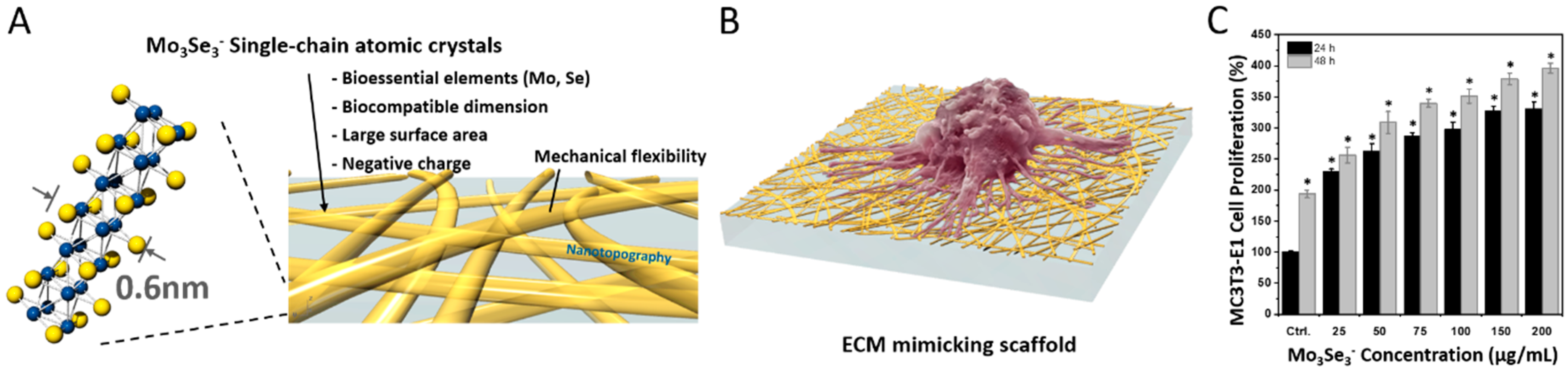
| Mo3Se3− SCAC nanowire | Inorganic 1D nanowire of 0.6 nm in diameter | L929 fibroblast cells and MC3T3-E1 osteoblast cells | Significant increase in the proliferation of cells was observed in the presence of 1D nanowires. | [34] |
| Au nanomaterials coated with bovine serum albumin (BSA) | Nanospheres, nanostars, and nanorods of sizes 40 nm, 70 nm, and 100 nm | hMSCs | Size and shape dependent osteogenic differentiation of cells occurred. Nanospheres (40 nm and 70 nm) and nanorods (70 nm) increased the alkaline phosphate activity (ALP) and calcium deposition of the cells. | [35] |
| PAAm gel micropatterned with collagen I | Stiffness of PAAm gel: Soft (~1 kPa) and stiff (~7 kPa). Diverse shapes of micropatterns with identical area: Square, triangular, and rectangular | MCF-10A cells | Cell−cell junctions could be impaired as matrix became stiffer and the cell shapes became more elongated by the micropatterns. The cell generated tractions that were increased progressively as the pattern shapes were changed from squares to triangles and rectangles. | [40] |
| Au islands coated with fibronectin | Geometry: Square (250 μm or 500 μm edge), rectangular (125 × 500 μm), and circular (564 μm in diameter) | Normal rat kidney epithelial cells | Geometries of micropatterns altered the cell proliferation by affecting cytoskeletal tension. High cell proliferation was observed on the edges and corners of the square islands. | [37] |
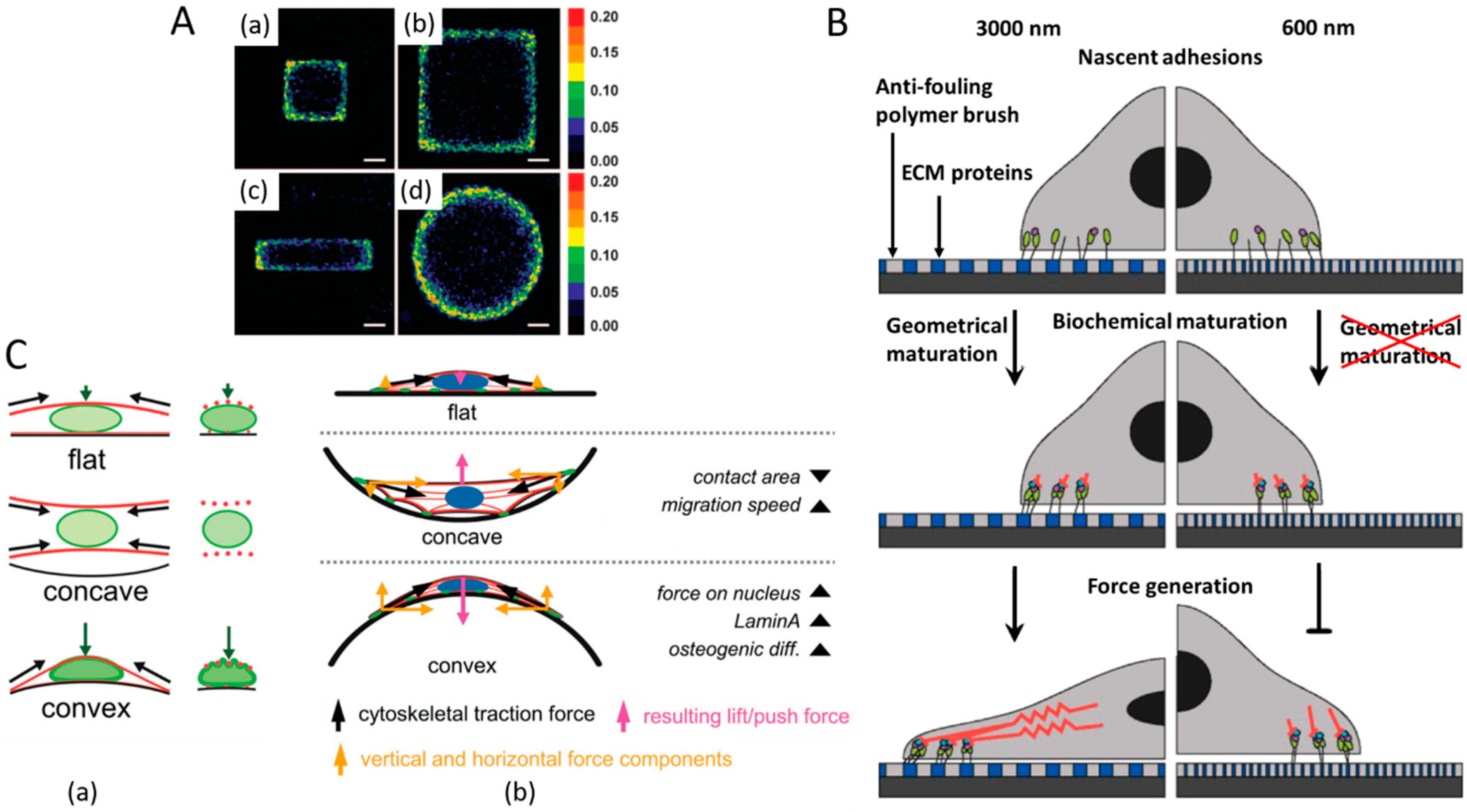
| Au substrates coated with fibronectin | Circular shape with different diameters (100, 300, 600, and 3000 nm) | Epidermal stem cells | Nanoscale adhesion geometry determined the fate of epidermal stem cells by changing cell shape and AP-1 transcription activity. | [42] |
| Poly(trimethylene carbonate) | 3D microtopographic cell culture chips with concave and convex spherical structures (250 μm in diameter and 1/125 μm−1 as principal curvature) | hMSCs | Cytoskeleton-tension-associated pull force on the concave surface: enhanced the cell attachment and increased its migration speed. Push force on convex surface: caused increases in osteogenic differentiation, lamin-A levels, and nuclear deformation. | [43] |
| Fibronectin fibers and poly oligo(ethylene glycol methyl ether methacrylate) brushes | Quasi-2D fibrous pattern (Dimension: 250, 550, 800, and 1000 nm width, Density: 22 ± 8% and 60 ± 5%) | HaCaT cells | Nanoscale geometry of the ECM acted as an important regulator for cell adhesion, spreading, and shaping. Nanofibrous structures allowed cell adhesions to develop along one axis. | [44] |
| Au | 3D leaf-like structure (nanospikes) | hMSCs | 3D nanostructured architecture promoted MSC alignment and neurogenic differentiation | [45] |
| PDMS coated with collagen I | Smooth and microgrooved topography (10 μm wide, 10 μm apart, and 5 μm deep) Stiffness: 90 ± 8 kPa (soft) and 1500 ± 110 kPa (hard) | hMSCs | Microgrooved stiff substrate led to high cell viscoelastic properties and expression of α-actin and h1-calponin | [48] |
| PDMS coated with fibronectin | Nanoscale gratings and pillars: 300 nm, 500 nm, and 1000 nm width and diameter Height: 150 nm, 300 nm, and 560 nm | NHLF cells | Nanoscale gratings and pillars facilitated focal adhesion of cells. Nanogratings oriented focal adhesions and nuclei along the nanograting directions. | [49] |
| PDMS coated with gelatin | Micropatterned substrate: Height (1.5 μm), Groove width (2, 3, 4, and 5 μm), Ridge width (2, 3, 4, and 5 μm) | An accelerated aging cell model derived from induced pluripotent stem cells (iPSCs) | Substrates with specific micropatterns, such as groove width of 5 μm and ridge width of 5 μm, led to higher cell aging via disruption of the connection between the cytoskeleton and nucleoskeleton and triggering of DNA damage | [50] |
| Ti surface | Nanotopographic pattern, wettability, and mechanical strength | hGF cells | Ti surfaces with pore diameter (74 nm), surface roughness (41.6 nm), surface area (30.4 μm2), and hydrophilicity (65.5°) resulted in enhanced cell attachment, proliferation, and differentiation | [56] |
| Material/Substrates | Properties | Cell Type | Effects | Ref. |
|---|---|---|---|---|
| Ppy array on Ti surface | Highly adhesive hydrophobic nanotubes and poorly adhesive hydrophilic nanotips | MSCs | The dynamic switching of nanotube/nanotip induced osteogenic differentiation of the cells | [59] |
| RGD-grafted Fe3O4 coated silica | Magnetic field induced variation in RGD tether length and mobility on material surface | hMSCs | Restriction in the mobility of RGD on material surface, caused by magnetic field, resulted in enhanced cell adhesion, spreading, and osteogenic differentiation | [60] |
| Material/Substrates | Properties | Cell Type | Effects | Ref. |
|---|---|---|---|---|
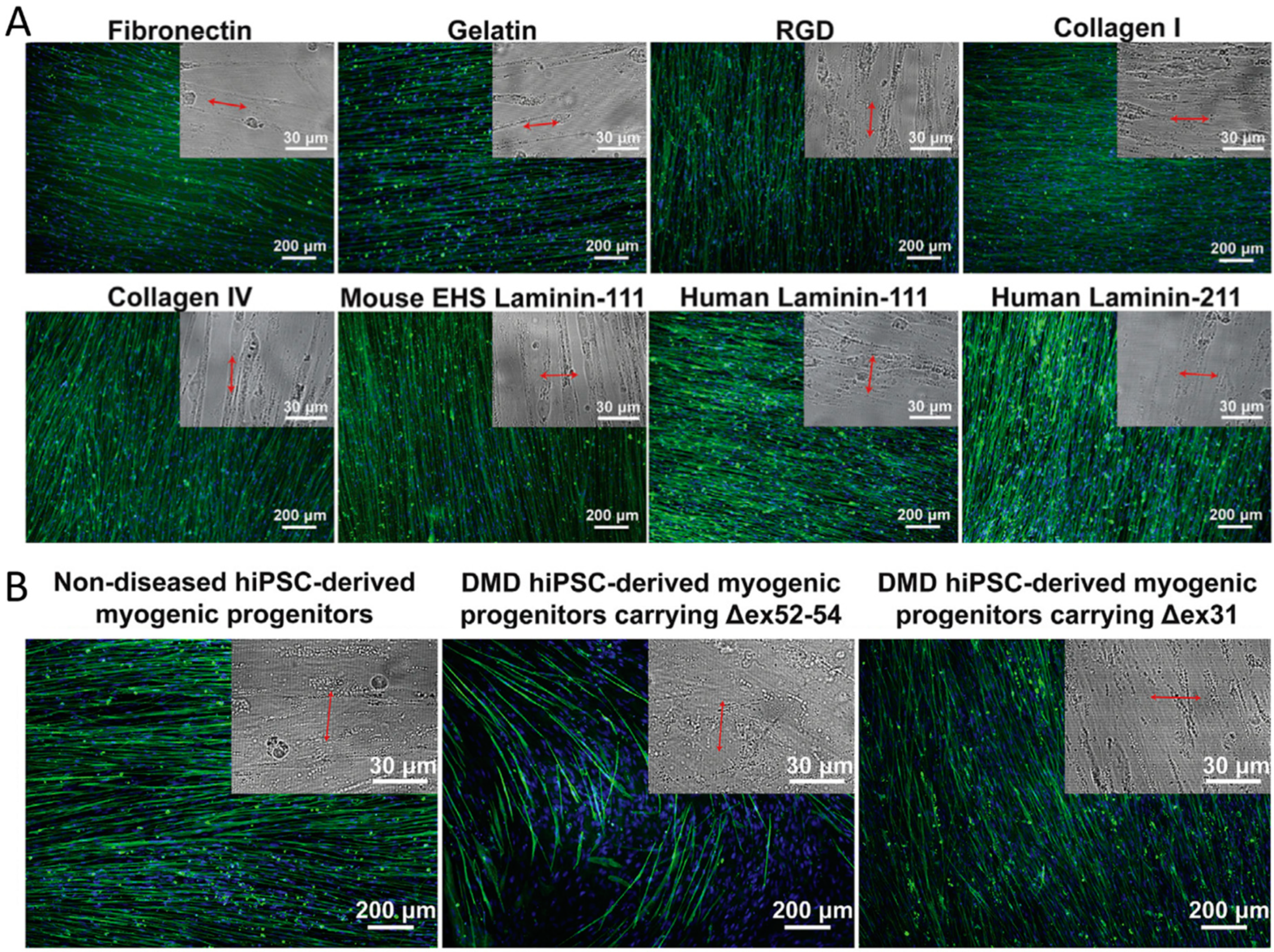
| PDMS Topography: 500, 800, 1000, 1500, and 3000 nm width parallel grooves (400 nm depth) | Functionalization with Matrigel, laminin-111, laminin-211, gelatin, RGD peptide, fibronectin, collagen I, and collagen IV | hESCs | Myotubes aligned perpendicularly on matrigel-functionalized 800 nm nanogrooved substrate through DAPC-mediated cytoskeleton–ECM linkage | [65] |
| PDMS Topography: Nanopillars, microwells, and micropillars | Functionalization: Fibronectin mixed with collagen I (FC) and laminin mixed with chondroitin sulfate | Human corneal endothelial cells | Micropillars functionalized with FC had high Na+/K+ ATPase and zonula occludens-1 (ZO-1) expression, resulting in enhanced circularity | [66] |
| Titanium (Ti) | Functionalization: Allylamine plasma polymer layer (PPAAm) | MG-63 osteoblastic cells | Amino groups promoted focal contact formation, such as vinculin, paxillin, p-FAK | [71] |
| Au | Functionalization: Self-assembled monolayers of alkanethiols like 1-dodecanethiol [*CH3 (hydrophobic)], 11-mercapto-1-undecanol [*OH (neutral and hydrophilic)], 11-mercaptoundecanoic acid [*COOH (negatively charged at pH 7.4)], and 12-amino-1-mercaptododecane [*NH2 (positively charged at pH 7.4)] * - functional groups | MC3T3-E1 osteoblast cells | OH- and NH2-terminated Au surfaces resulted in the selective binding of α5β1 and αVβ3 integrin for better focal adhesion composition, osteoblast differentiation, signaling, and mineralization | [72] |
| Integrin Subunit | Ligand | |
|---|---|---|
| β | α | |
| β1 | α1 | Collagen, Laminin |
| α2 | Collagen, Laminin, Thrombospondin, E-cadherin, Tenascin C | |
| α10 | Collagen, Laminin | |
| α11 | Collagen | |
| α3 | Laminin, Thrombospondin | |
| α6 | Laminin | |
| α7 | Laminin, Tenascin C | |
| β4 | α6 | Laminin, Thrombospondin |
| β1 | α4 | Fibronectin, Thrombospondin, Osteopontin, VCAM-1, ICAM-4 |
| α5 | Fibronectin, Osteopontin | |
| α8 | Fibronectin, Vitronectin, Osteopontin, Tenascin C | |
| α9 | Osteopontin, Tenascin C, VCAM-1 | |
| αV | Fibronectin, Osteopontin, LAP TGF-β | |
| β5 | αV | Vitronectin, Osteopontin, LAP TGF-β |
| β6 | Fibronectin, Osteopontin, Tenascin C, LAP TGF-β | |
| β8 | LAP TGF-β | |
| β3 | Fibrinogen, Fibronectin, vWF, Vitronectin, Thrombospondin, Osteopontin, ICAM-4, Tenascin C | |
| β3 | αIIb | Fibrinogen, Fibronectin, vWF, Vitronectin, Thrombospondin, ICAM-4 |
| β7 | α4 | Fibronectin, Osteopontin, VCAM-1, |
| Leukocyte-Specific | ||
| β7 | αE | E-cadherin |
| β2 | αL | ICAM-4 |
| αM | Fibrinogen, ICAM-4 | |
| αX | Fibrinogen, ICAM-4, Collagen | |
| αD | Fibronectin, Vitronectin, Fibrinogen, VCAM-1, ICAM-3 | |
| Receptors | Main Findings | Ref. |
|---|---|---|
| Integrins | Integrin can sense diverse physical characteristics of engineered materials such as topography and viscosity. | [7,31] |
| Integrin also sense ECM proteins and specific motifs of those proteins when incorporated on engineered materials. | [31,78] | |
| Integrin is a heterodimer composed of α and β subunits and each integrin selectively binds to different ligands. | [31,76,77,78,85] | |
| Ligand binding of integrins is controlled by conformational rearrangement between an inactive bent form and an active extended form. | [79,87,88] | |
| Mechanosensitive channels | Piezo channels have been identified as the channels that sense various physical stimuli through transmission by lipid bilayer tension. | [89,90,91,92,93] |
| Piezo channels act on coupling of the mechanical stimuli with ion flux. | [90,92,93] | |
| Piezo1 channel is activated by various physical stimuli, including pressure, indentation, deflection, and membrane stretch, while TRPV4 is activated only by deflection stimulus. | [94] | |
| GPCRs | Several cellular environmental stimuli such as shear stress, osmotic changes, and mechanical pressure can lead to a conformational change of GPCR from an inactive state to an active state. | [97,98,99] |
| Fluid-induced shear stress, hypotonic stress, and fluidizing agents have the same effect on GPCR. | [98] | |
| Various engineered materials have been used to mimic cellular dynamic environment to investigate GPCR-mediated sensing. | [98,99,100] |
| Signal Transfer | Main Findings | Ref. |
|---|---|---|
| Integrin-mediated transfer and roles of signaling molecules | Integrin–substrate binding promotes the formation of focal adhesion complex along with recruitment of signaling molecules that subsequently activate or localize other proteins. | [20,80,102,103] |
| Beginning with the autophosphorylation of a tyrosine residue in FAK, various focal adhesion proteins can be activated sequentially through phosphorylation. | [20,102] | |
| Diverse physical features of cell surroundings such as stiffness and topography lead to changes in localization of focal adhesion proteins. | [4,86,105,106] | |
| Integrin-mediated transfer and roles of cytoskeletal structure | Transduction of physical signals from substrates into cells requires intact connection between integrin and the actin cytoskeleton. | [107] |
| The integrin–ligand interaction induces recruitment of structural proteins such as vinculin, talin, and tensin1 and adaptor proteins that stabilize the cytoskeletal structure. | [108,109] | |
| β3 integrin and the ERM family, which acts as integrin adapter protein and signaling molecule, mediate transduction of the information of 3D biomimetic microchips. | [110] | |
| GPCR-mediated signal transfer | Receptors coupling with Gq/11 proteins activate IP3 and DAG formation and GS protein-coupled receptors activate cAMP formation. | [100,112] |
| GPR4, which senses changes in pH, acts as a GS-coupled receptor and GPR68 acts as a Gq/11-coupled receptor. | [97,99] |
| Main Findings | Ref. |
|---|---|
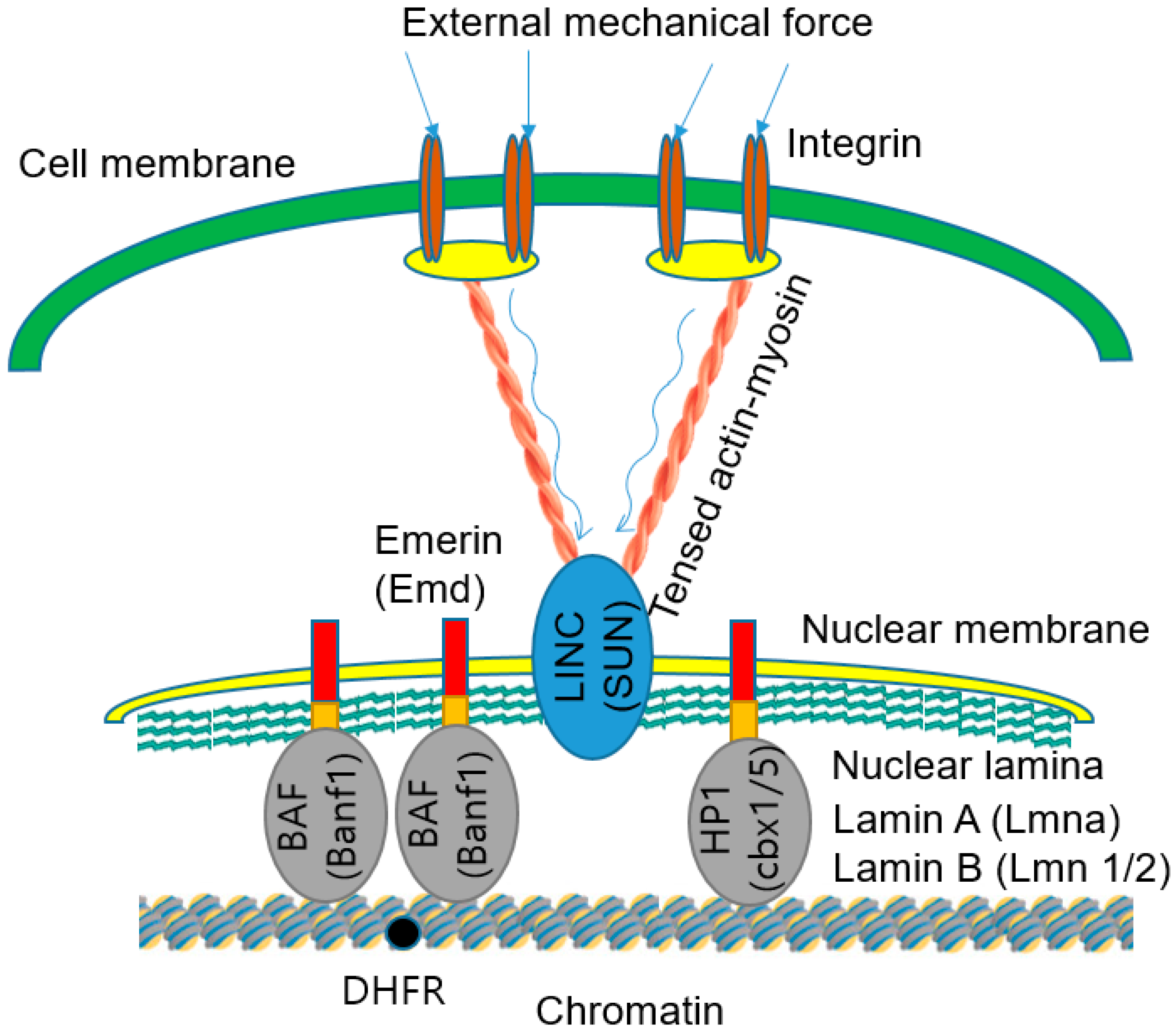
| LINC complexes act as bridges across the perinuclear space by coupling KASH family members and SUN family members. | [101,115,116] |
| The cytoplasmic domains of the KASH proteins interact with cytoskeletal elements and the exposed residues of the KASH proteins bind to the C-termini of SUN proteins. | [116] |
| N-termini of SUN proteins bind to the nuclear lamins. | [116] |
| YAP/TAZ complex provides bidirectional biochemical connections. | [113,114,115] |
| YAP/TAZ-mediated regulation requires Rho GTPase activity and tension of the actin cytoskeleton. | [113] |
| LINC complex-mediated nuclear mechanotransduction can be induced when the signal molecules phosphorylate several structural proteins. | [115] |
| YAP/TAZ States and the Affected Cellular Behaviors | Main Findings | Ref. | |
|---|---|---|---|
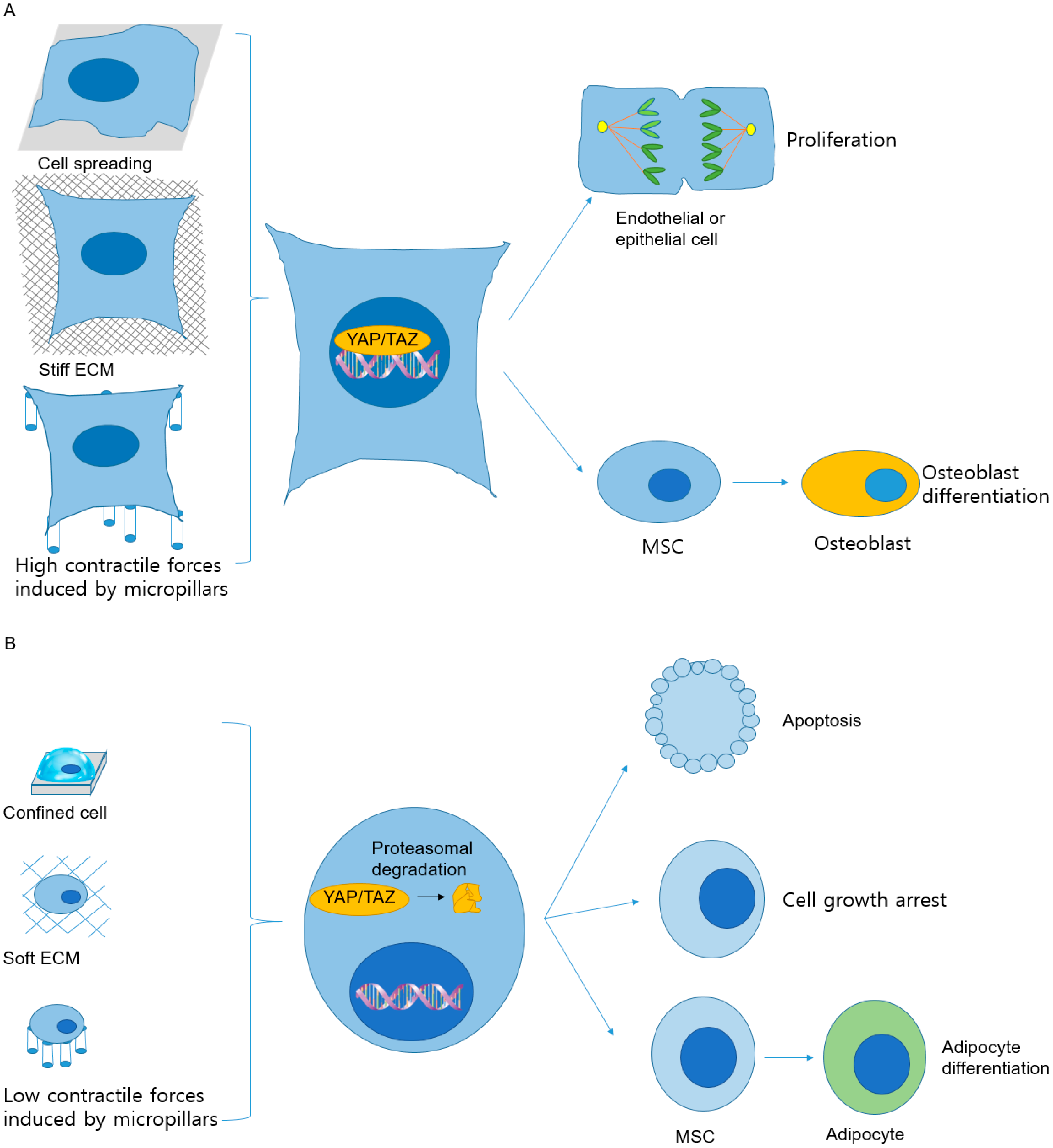
| YAP/TAZ activation | Proliferation | YAP and TAZ activity, regulated by mechanical properties of multicellular sheets, controls the proliferative capacity of cells. | [137] |
| YAP distribution and cell density/cell adhesion area (NIH 3T3 cells) are correlated. | [135] | ||
| The proliferation of endothelial cells is promoted by disturbed flow that causes the activation of YAP/TAZ. | [138] | ||
| YAP1 is an essential modulator for the proliferation of epidermal stem cell and tissue expansion. | [139] | ||
| Osteoblast differentiation | MSCs differentiation is affected by YAP/TAZ activity, which links to mechanical cues from ECM. | [113] | |
| Runx2-involved gene transcription, repression of PPARγ-involved gene transcription, and differentiation of MSCs are regulated by TAZ. | [140] | ||
| ECM stiffness-dependent osteogenesis of MSCs is promoted by vinculin and enhanced nuclear localization of TAZ. | [141] | ||
| MSC differentiation is extremely sensitive to tissue level elasticity of ECMs. | [8] | ||
| Shapes of mesenchymal progenitors are regulated by MT1-MMP, which results in nuclear localization of YAP and TAZ. | [142] | ||
| YAP activity-dependent MSCs differentiation is regulated by shear stress of cellular environment. | [143] | ||
| YAP/TAZ inactivation | Apoptosis | YAP inactivation, caused by the detachment of MCF10A cells, induces anoikis, a kind of apoptosis. | [144] |
| Cell growth arrest | Inactivation of TAZ results in growth arrest of glioma cells. | [145] | |
| YAP inactivation is involved in cell growth arrest and cell contact inhibition. | [146] | ||
| Adipocyte differentiation | MSCs differentiation is regulated by YAP/TAZ activity responding to mechanical cues from ECM stiffness. | [113] | |
| Runx2-involved gene transcription, repression of PPARγ-involved gene transcription and differentiation of MSCs are regulated by TAZ. | [140] | ||
| MSC differentiation is extremely sensitive to tissue level elasticity of ECMs. | [8] | ||
| Shapes of mesenchymal progenitors is regulated by MT1-MMP. | [142] | ||
| YAP activity-dependent MSCs differentiation is regulated by the shear stress of the cellular environment. | [143] | ||
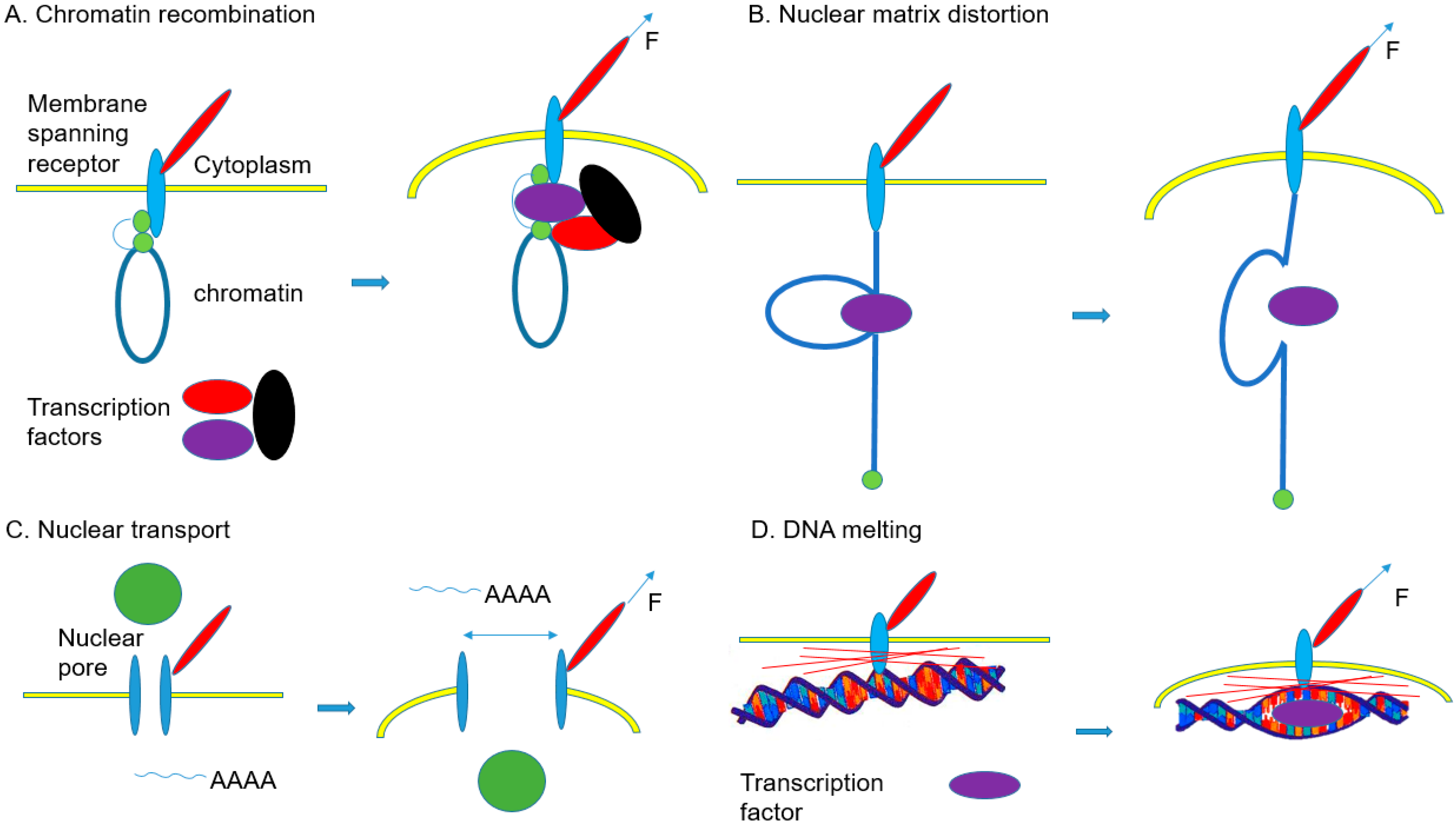
| Nuclear Changes | Main Findings | Ref. |
|---|---|---|
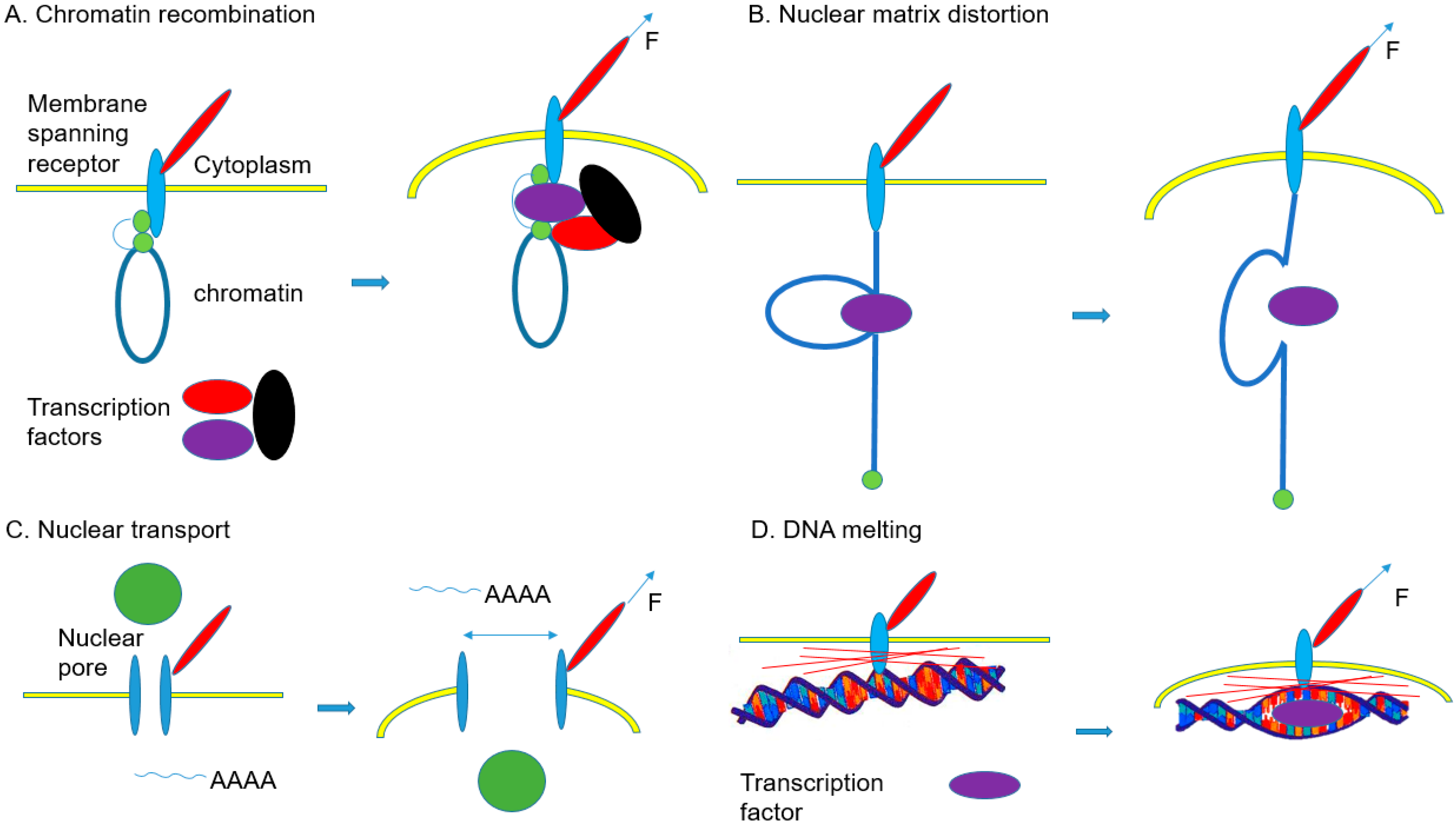
| Chromatin recombination | Micropattern-induced reduction of HDAC3 nuclear localization results in decondensation of chromatin. Gene transcription is regulated by chromatin compaction. | [154] |
| Decrease of Emd at the inner membrane of the nucleus by extrinsic biaxial mechanical strain leads to the reduction of H3K9me2,3 on chromatin and rearrangements of chromatin for the regulation of gene expression. | [155] | |
| There are rearrangements of specific chromosomes containing the genes that are regulated by cell geometries. These rearrangements are caused by physical cues from the patterns of cell culturing substrates. | [156] | |
| Chromatin deformation by magnetic force-induced local stress on CHO cells upregulates the DHFR expression. | [124] | |
| Nuclear matrix distortion | Enhancement of tissue-specific differentiation by mechanotransduction through nuclear lamin A High level of lamin A of cells on stiff matrix stabilizes the nucleus, lamina, and chromatin, which may affect the epigenetic stability and the extent of DNA breaks. Tissue-specific gene expression is regulated by lamin A levels. | [122] |
| Acute perturbations of ECM elasticity results in alterations in the levels of lamin A and DNA damage. Slow degradation of lamin A by low phosphorylation leads to lower DNA damage in contractile cells cultured on stiff ECM. | [157] | |
| There are synergetic effects of collagen matrix rigidity and retinoids on the differentiation of MSCs to osteoblasts. Retinoic acid receptor transcription factors regulate the expression of lamin A. | [158] | |
| Nuclear transport | A mechanotransduction-induced stretch of nuclear pores leads to increase of YAP nuclear localization on stiff ECM. | [159] |
| A mechanotransduction-induced stretch of nuclei during cell spreading caused the release of perinuclear Ca2+ and elevation of Ca2+ level in the nucleus | [160] | |
| Translocation of cytosolic phospholipase A2 and elevation of Ca2+ by nuclear swelling | [161] | |
| DNA melting | Tethering of destabilized DNA regions on MARs results in melting of the double helix. | [162] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, Y.-h.; Jin, X.; Shankar, P.; Lee, J.H.; Jo, K.; Lim, K.-i. Molecular-Level Interactions between Engineered Materials and Cells. Int. J. Mol. Sci. 2019, 20, 4142. https://doi.org/10.3390/ijms20174142
Jang Y-h, Jin X, Shankar P, Lee JH, Jo K, Lim K-i. Molecular-Level Interactions between Engineered Materials and Cells. International Journal of Molecular Sciences. 2019; 20(17):4142. https://doi.org/10.3390/ijms20174142
Chicago/Turabian StyleJang, Yoon-ha, Xuelin Jin, Prabakaran Shankar, Jung Heon Lee, Kyubong Jo, and Kwang-il Lim. 2019. "Molecular-Level Interactions between Engineered Materials and Cells" International Journal of Molecular Sciences 20, no. 17: 4142. https://doi.org/10.3390/ijms20174142
APA StyleJang, Y.-h., Jin, X., Shankar, P., Lee, J. H., Jo, K., & Lim, K.-i. (2019). Molecular-Level Interactions between Engineered Materials and Cells. International Journal of Molecular Sciences, 20(17), 4142. https://doi.org/10.3390/ijms20174142