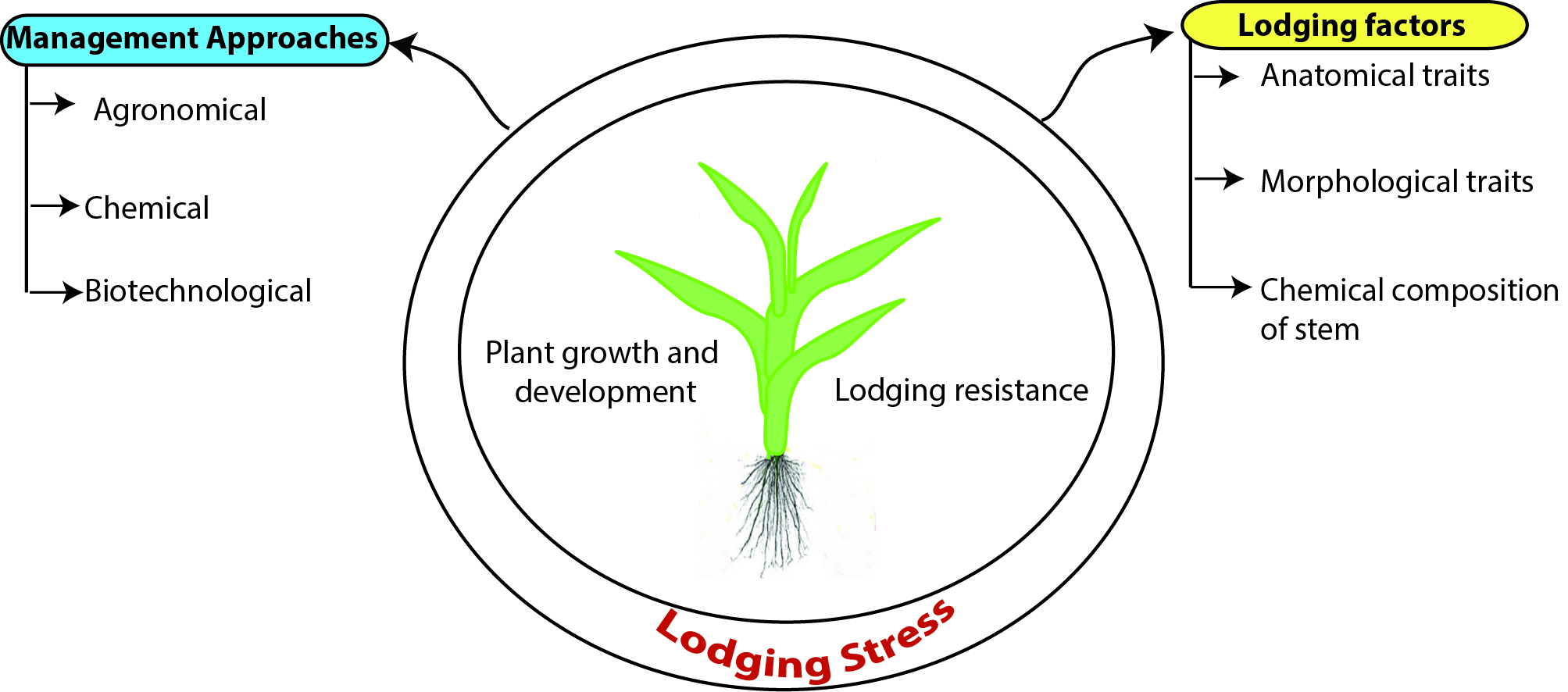
Improving Lodging Resistance: Using Wheat and Rice as Classical Examples

Abstract
: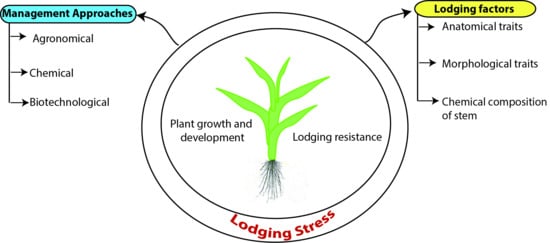
1. Introduction
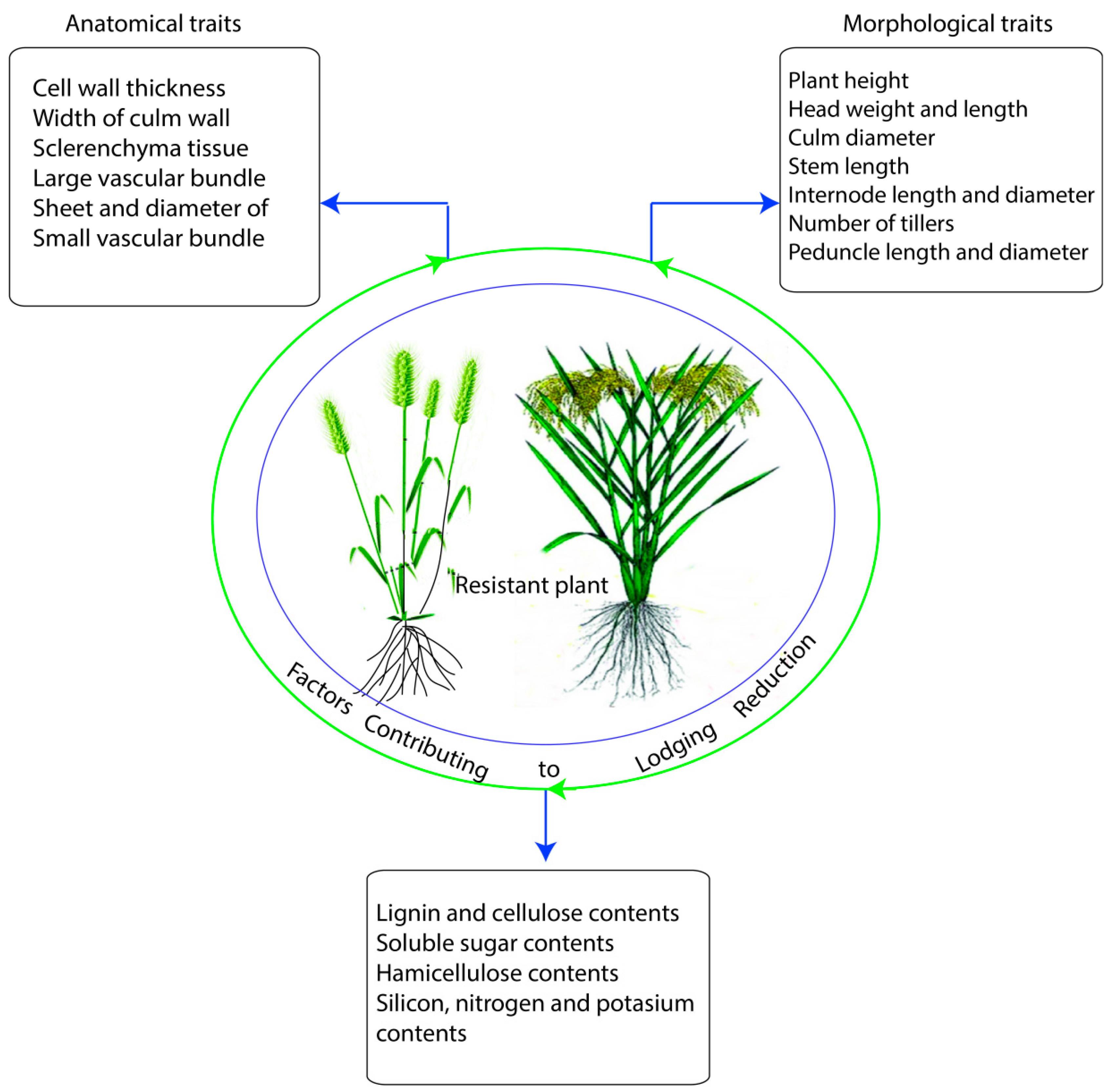
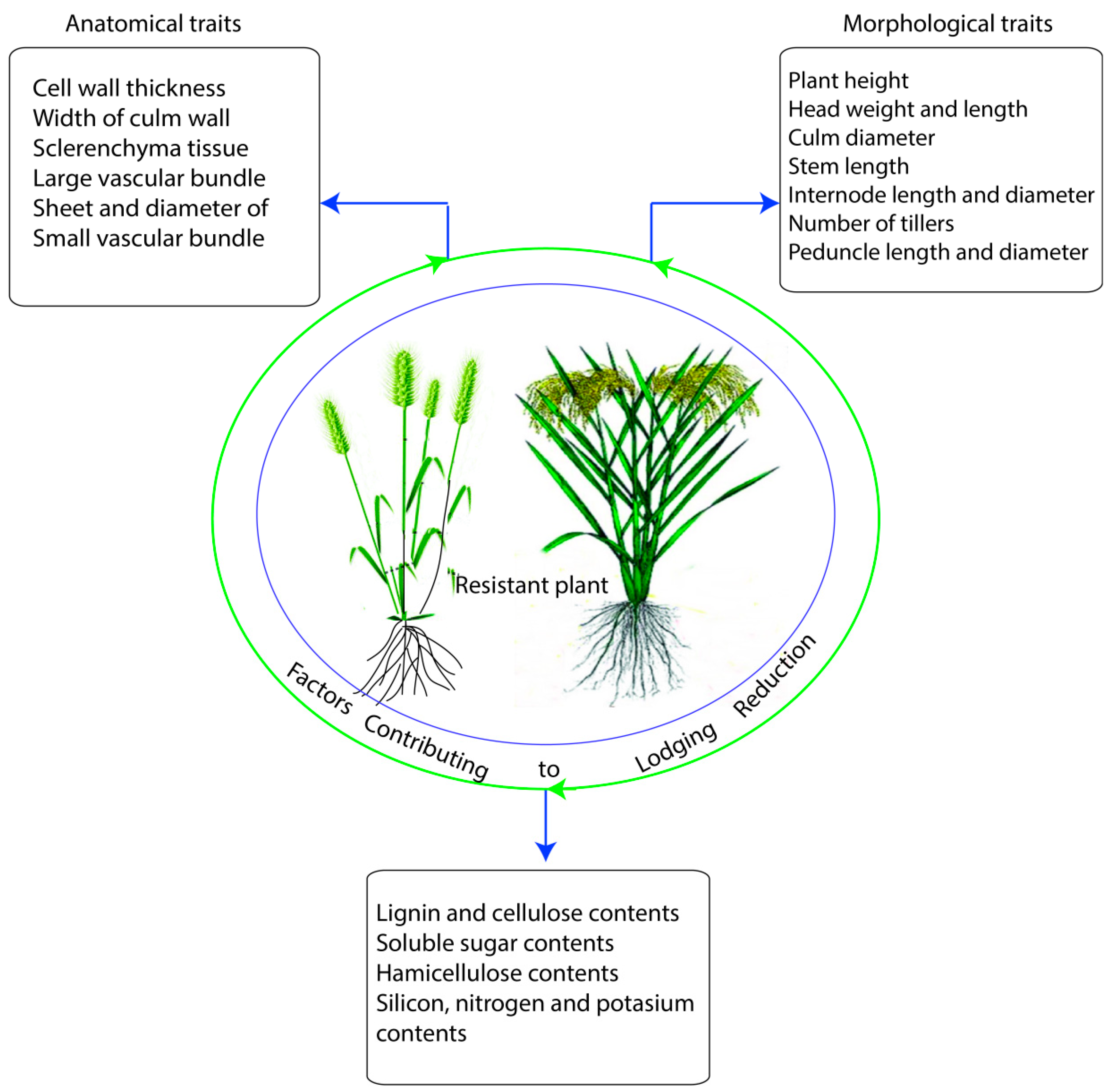
2. Morphological Traits and their Responses under Lodging Stress
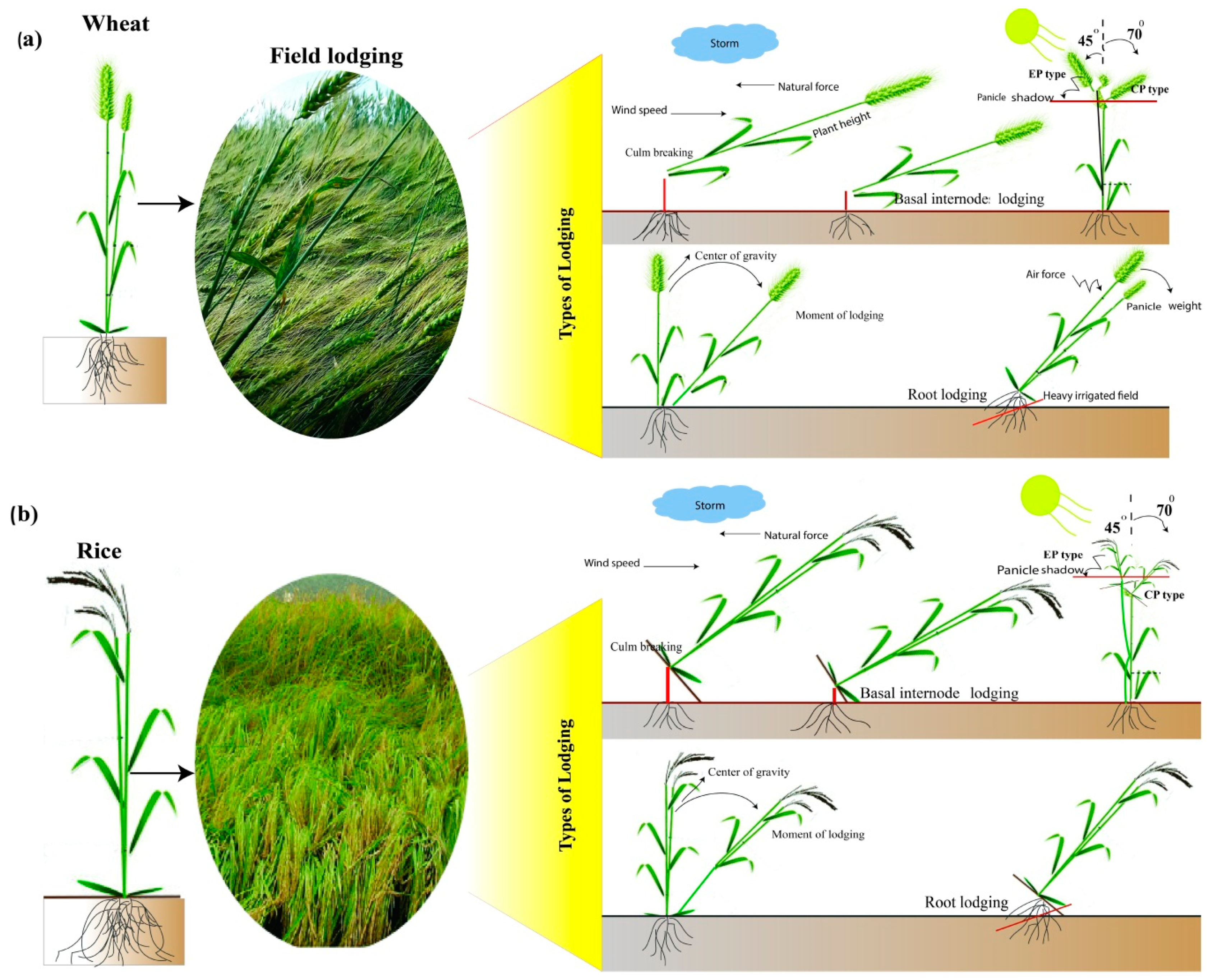
2.1. Plant Height and Inter-Nodal Length
2.2. Culm Wall Thickness, Culm Diameter, and Panicle Weight
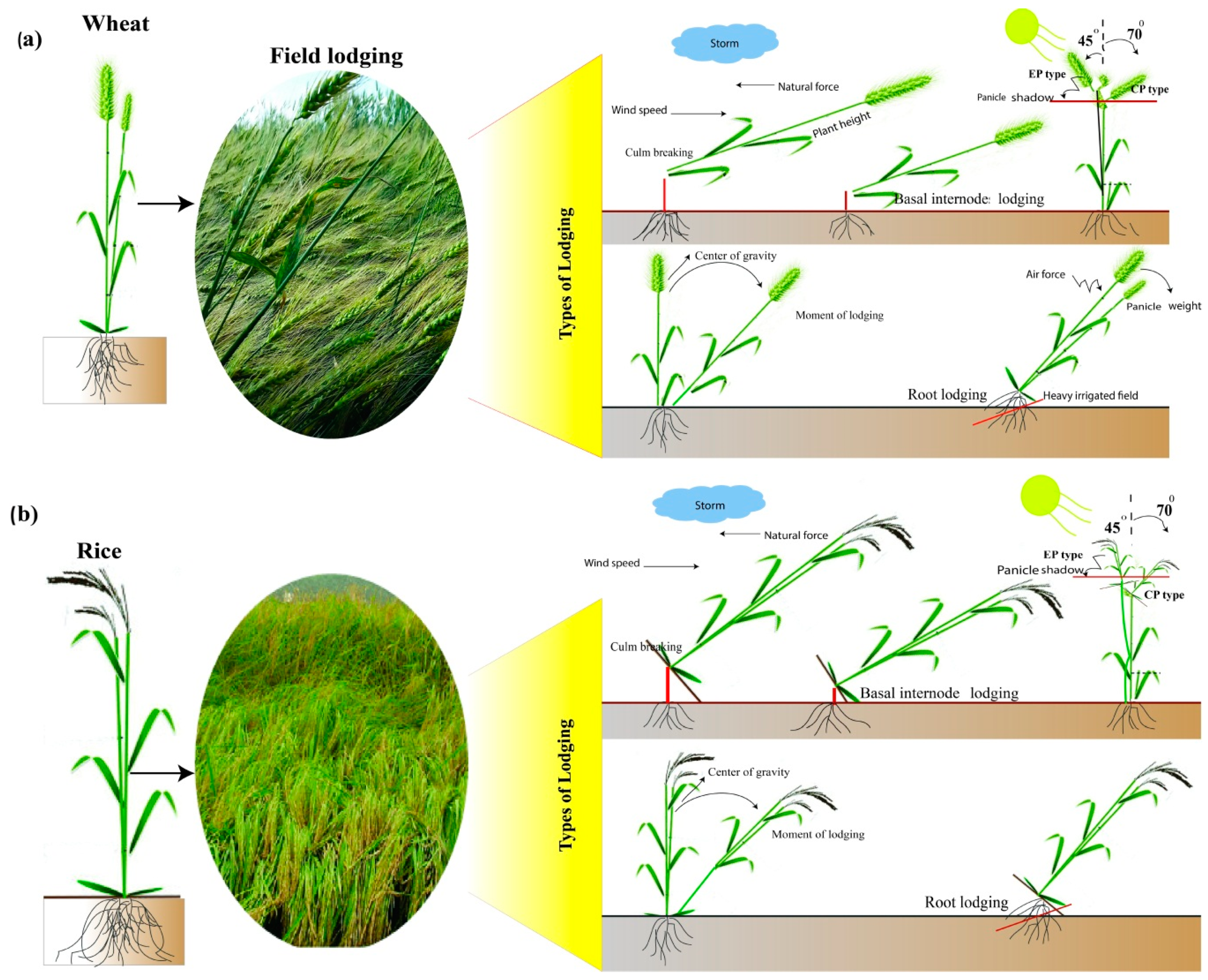
2.3. Root Lodging
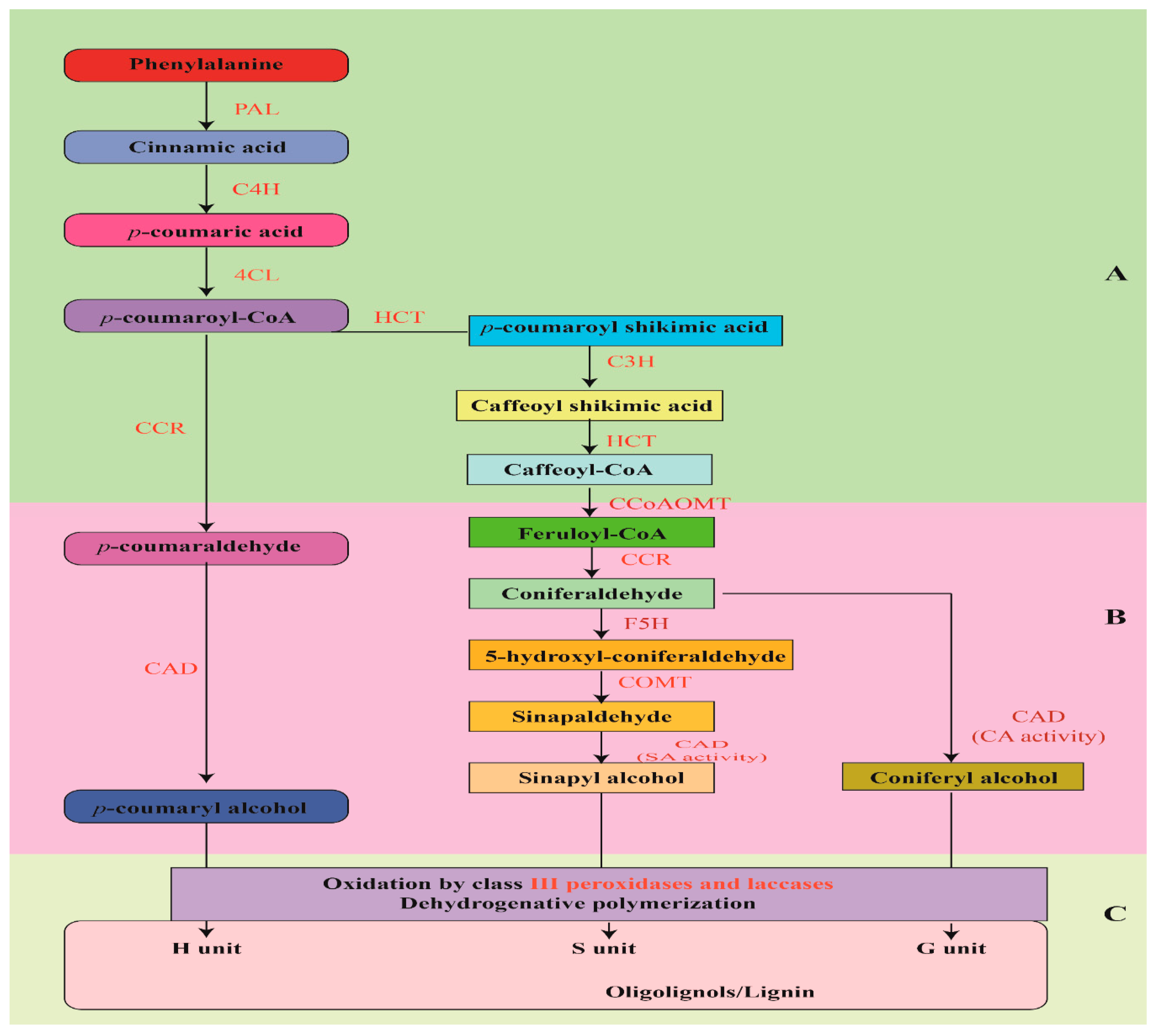
2.4. Carbohydrates and Role of Lignin Biosynthesis
3. Plant Hormones in Lodging Stress as a Key Regulator
4. Genetic Manipulation for Lodging Resistance
4.1. Quantitative Trait Loci
4.2. Gene’s Associated with Lodging Tolerance
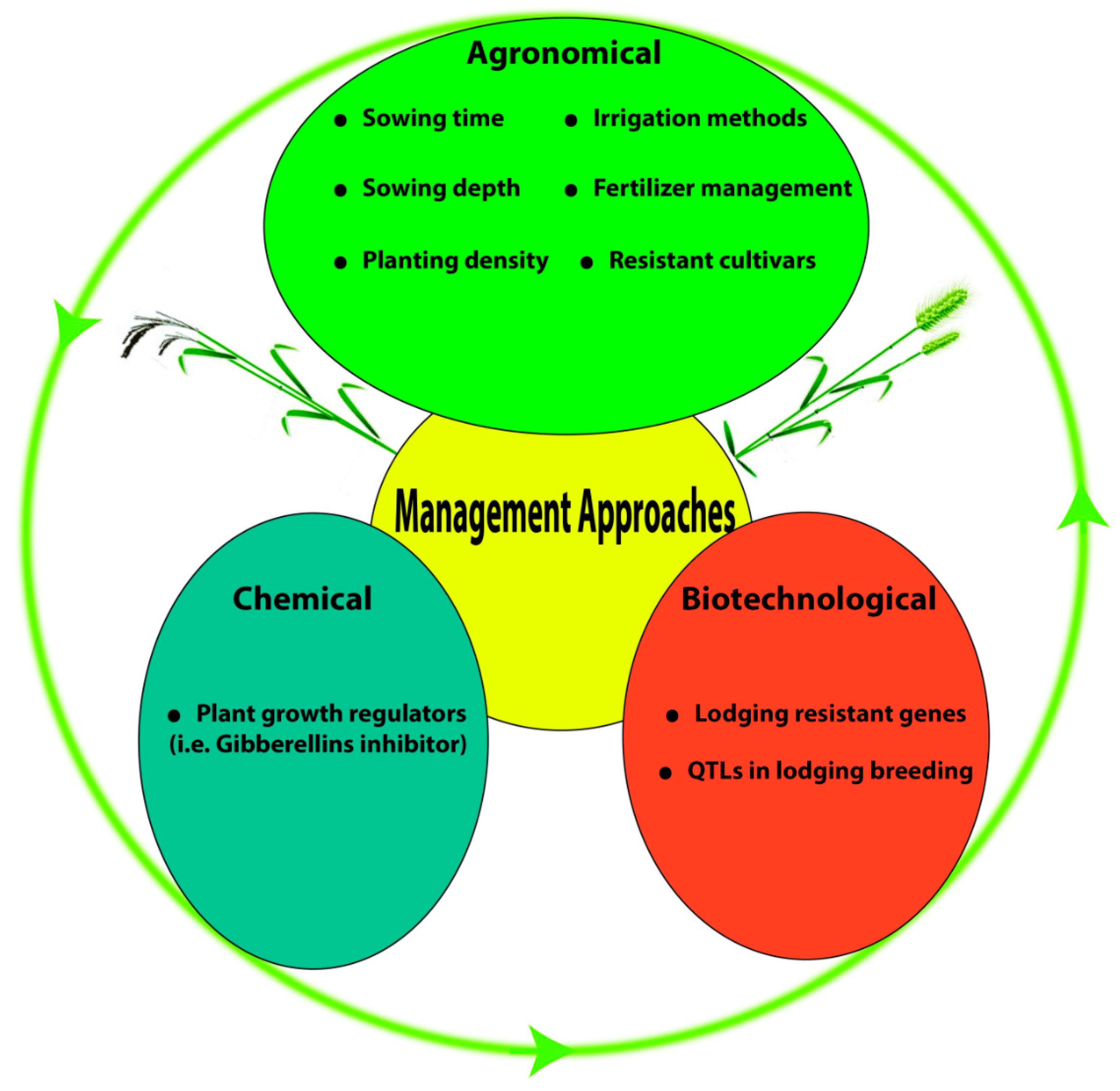
5. Agronomical Management
5.1. Sowing Time
5.2. Sowing Depth
5.3. Planting Density
5.4. Irrigation Method
5.5. Effect of Nitrogen
5.6. Effect of Potassium
5.7. Effect of Silicon
5.8. Lodging-Resistant Cultivars
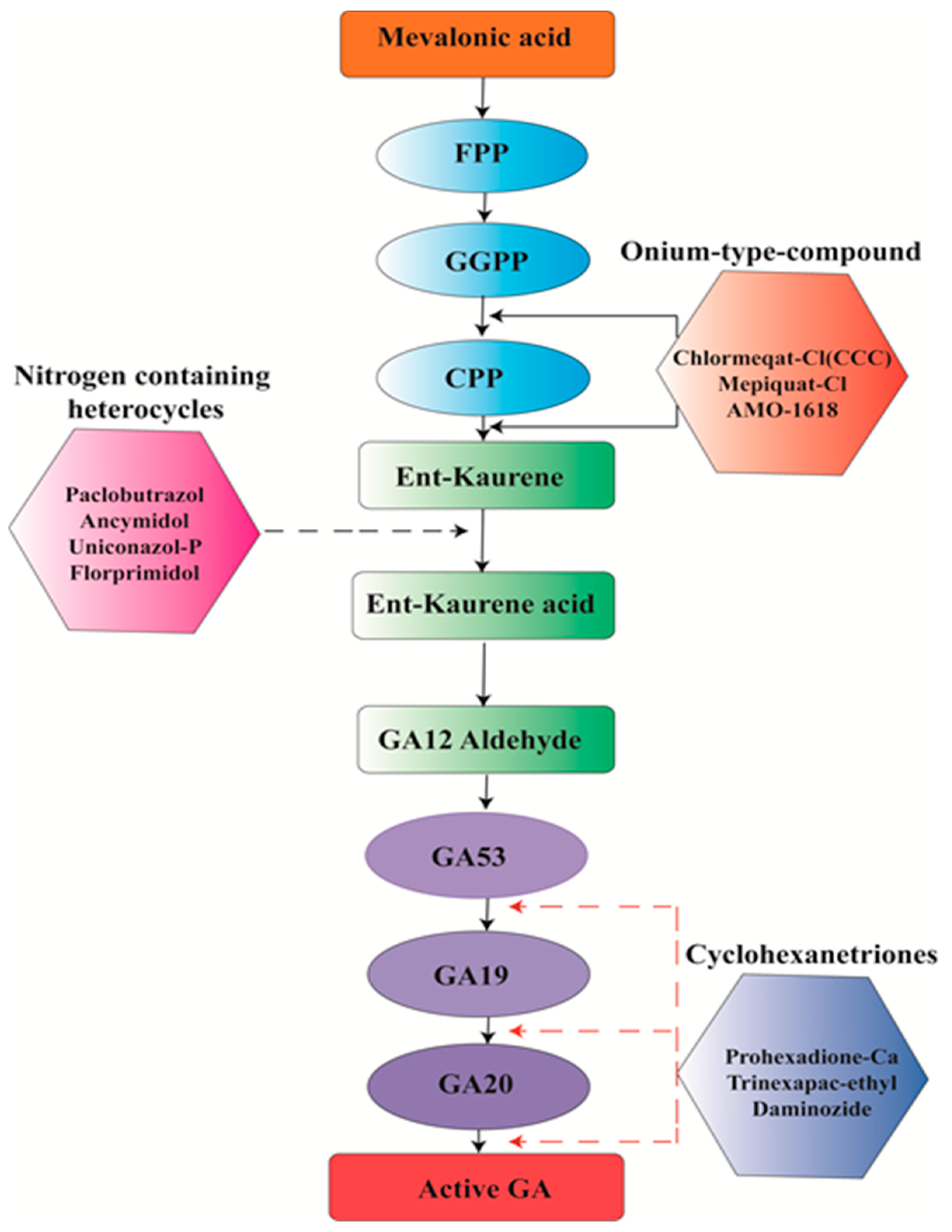
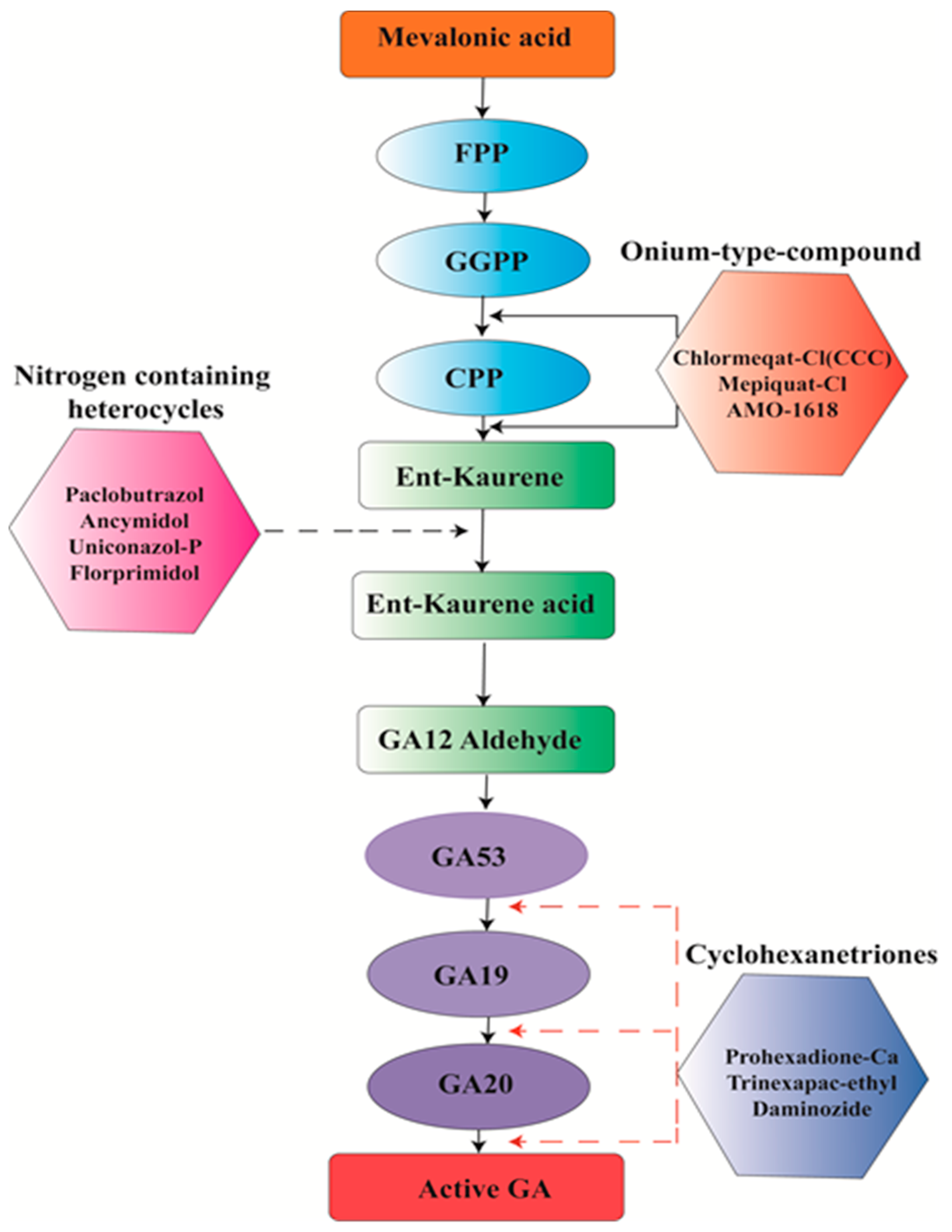
6. Chemical Management
7. Future Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Mulsanti, I.W.; Yamamoto, T.; Ueda, T.; Samadi, A.F.; Kamahora, E.; Rumanti, I.A.; Thanh, V.C.; Adachi, S.; Suzuki, S.; Kanekatsu, M. Finding the superior allele of japonica-type for increasing stem lodging resistance in indica rice varieties using chromosome segment substitution lines. Rice 2018, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Q.; Kuai, J.; Zhao, L.; Hu, Z.; Wu, J.; Zhou, G. The effect of sowing depth and soil compaction on the growth and yield of rapeseed in rice straw returning field. Field Crop. Res. 2017, 203, 47–54. [Google Scholar] [CrossRef]
- Kuai, J.; Yang, Y.; Sun, Y.; Zhou, G.; Zuo, Q.; Wu, J.; Ling, X. Paclobutrazol increases canola seed yield by enhancing lodging and pod shatter resistance in Brassica napus L. Field Crop. Res. 2015, 180, 10–20. [Google Scholar] [CrossRef]
- Berry, P.; Spink, J. Predicting yield losses caused by lodging in wheat. Field Crop. Res. 2012, 137, 19–26. [Google Scholar] [CrossRef]
- Peng, D.; Chen, X.; Yin, Y.; Lu, K.; Yang, W.; Tang, Y.; Wang, Z. Lodging resistance of winter wheat (Triticum aestivum L.): Lignin accumulation and its related enzymes activities due to the application of paclobutrazol or gibberellin acid. Field Crop. Res. 2014, 157, 1–7. [Google Scholar] [CrossRef]
- Zhang, W.-J.; Li, G.-H.; Yang, Y.-M.; Quan, L.; Zhang, J.; Liu, J.-Y.; Shaohua, W.; She, T.; Ding, Y.-F. Effects of nitrogen application rate and ratio on lodging resistance of super rice with different genotypes. J. Integr. Agric. 2014, 13, 63–72. [Google Scholar] [CrossRef]
- Berry, P.M. Predicting Lodging in Winter Wheat. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 1998. [Google Scholar]
- Acreche, M.M.; Slafer, G.A. Lodging yield penalties as affected by breeding in Mediterranean wheats. Field Crop. Res. 2011, 122, 40–48. [Google Scholar] [CrossRef]
- Niu, L.; Feng, S.; Ding, W.; Li, G. Influence of speed and rainfall on large-scale wheat lodging from 2007 to 2014 in China. PLoS ONE 2016, 11, e0157677. [Google Scholar] [CrossRef] [PubMed]
- Kheiralla, K.; Mahdy, E.; Dawood, R. Evaluation of Some Wheat Cultivars for Traits Related to Lodging Resistance Under Different Levels of Nitrogen. Assiut J. Agric. Sci. 1993, 24, 257–271. [Google Scholar]
- Berry, P.; Sterling, M.; Spink, J.; Baker, C.; Sylvester-Bradley, R.; Mooney, S.; Tams, A.; Ennos, A. Understanding and reducing lodging in cereals. Adv. Agron. 2004, 84, 215–269. [Google Scholar]
- Foulkes, M.J.; Slafer, G.A.; Davies, W.J.; Berry, P.M.; Sylvester-Bradley, R.; Martre, P.; Calderini, D.F.; Griffiths, S.; Reynolds, M.P. Raising yield potential of wheat. III. Optimizing partitioning to grain while maintaining lodging resistance. J. Exp. Bot. 2010, 62, 469–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piñera-Chavez, F.; Berry, P.; Foulkes, M.; Molero, G.; Reynolds, M. Avoiding lodging in irrigated spring wheat. II. Genetic variation of stem and root structural properties. Field Crop. Res. 2016, 196, 64–74. [Google Scholar] [CrossRef]
- Setter, T.; Laureles, E.; Mazaredo, A. Lodging reduces yield of rice by self-shading and reductions in canopy photosynthesis. Field Crop. Res. 1997, 49, 95–106. [Google Scholar] [CrossRef]
- Weng, F.; Zhang, W.; Wu, X.; Xu, X.; Ding, Y.; Li, G.; Liu, Z.; Wang, S. Impact of low-temperature, overcast and rainy weather during the reproductive growth stage on lodging resistance of rice. Sci. Rep. 2017, 7, 46596. [Google Scholar] [CrossRef] [PubMed]
- Haghdoost, P.; Daliri, M.S.; Mohamadian, M.; Ghasm, T. The study of lodging occurrence stages on grain yield damages and agronomical traits of two rice cultivars (Tarom and Shiroodi). Adv. Environ. Biol. 2014, 8, 914–918. [Google Scholar]
- Lang, Y.-Z.; Yang, X.-D.; Wang, M.-E.; Zhu, Q.-S. Effects of lodging at different filling stages on rice yield and grain quality. Rice Sci. 2012, 19, 315–319. [Google Scholar] [CrossRef]
- Setter, T.; Ellis, M.; Laureles, E.; Ella, E.; Senadhira, D.; Mishra, S.; Sarkarung, S.; Datta, S. Physiology and genetics of submergence tolerance in rice. Ann. Bot. 1997, 79 (Suppl. 1), 67–77. [Google Scholar] [CrossRef]
- Liu, F.; Wang, P.; Zhang, X.; Li, X.; Yan, X.; Fu, D.; Wu, G. The genetic and molecular basis of crop height based on a rice model. Planta 2018, 247, 1–26. [Google Scholar] [CrossRef]
- ÜNAN, R.; Sezer, I.; ŞAHİN, M.; Mur, L.A. Control of lodging and reduction in plant length in rice (Oryza sativa L.) with the treatment of trinexapac-ethyl and sowing density. Turk. J. Agric. For. 2013, 37, 257–264. [Google Scholar] [CrossRef]
- Jennings, P.; Sornchai, S. The influence of lodging on rice yields. Int. Rice Com. Newsl. 1964, 13, 10–21. [Google Scholar]
- Mobasser, H.; Yadi, R.; Azizi, M.; Ghanbari, A.; Samdalari, M. Effect of density on morphological characteristics related-lodging on yield and yield components in varieties rice (Oryza sativa L.) in Iran. J. Agric. Environ. Sci. 2009, 5, 745–754. [Google Scholar]
- Lim, J.; Kwon, B.; Jung, B. Relationship between lodging-related characteristics and field lodging in rice. Korean J. Crop Sci. 1991, 36, 319–323. [Google Scholar]
- Wang, Y.; Wang, X.; Yang, L.; Li, P.; Zhu, J.; Kazuhiko, K.; Wang, Y. Ozone stress increases lodging risk of rice cultivar Liangyoupeijiu: A FACE study. Shengtai Xuebao/Acta Ecol. Sin. 2011, 31, 6098–6107. [Google Scholar]
- Pinthus, M.J. Lodging in wheat, barley, and oats: The phenomenon, its causes, and preventive measures. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 1974; Volume 25, pp. 209–263. [Google Scholar]
- Neenan, M.; Spencer-Smith, J. An analysis of the problem of lodging with particular reference to wheat and barley. J. Agric. Sci. 1975, 85, 495–507. [Google Scholar] [CrossRef]
- Kelbert, A.; Spaner, D.; Briggs, K.; King, J. The association of culm anatomy with lodging susceptibility in modern spring wheat genotypes. Euphytica 2004, 136, 211–221. [Google Scholar] [CrossRef]
- Chen, L.; Yang, Y.; Cui, C.; Lu, S.; Lu, Q.; Du, Y.; Su, R.; Chai, Y.; Li, H.; Chen, F. Effects of Vrn-B1 and Ppd-D1 on developmental and agronomic traits in Rht5 dwarf plants of bread wheat. Field Crop. Res. 2018, 219, 24–32. [Google Scholar] [CrossRef]
- Navabi, A.; Iqbal, M.; Strenzke, K.; Spaner, D. The relationship between lodging and plant height in a diverse wheat population. Can. J. Plant Sci. 2006, 86, 723–726. [Google Scholar] [CrossRef]
- Berry, P.; Berry, S. Understanding the genetic control of lodging-associated plant characters in winter wheat (Triticum aestivum L.). Euphytica 2015, 205, 671–689. [Google Scholar] [CrossRef]
- Yu, J.-K.; Graznak, E.; Breseghello, F.; Tefera, H.; Sorrells, M.E. QTL mapping of agronomic traits in tef [Eragrostis tef (Zucc) Trotter]. BMC Plant Biol. 2007, 7, 30. [Google Scholar] [CrossRef]
- Todaka, D.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Recent advances in the dissection of drought-stress regulatory networks and strategies for development of drought-tolerant transgenic rice plants. Front. Plant Sci. 2015, 6, 84. [Google Scholar] [CrossRef]
- Li, J.; Zhang, H.; Gong, J.; Chang, Y.; Huo, Z.; Xu, K.; Wei, H. Effects of different planting methods on the culm lodging resistance of super rice. Sci. Agric. Sin. 2011, 44, 2234–2243. [Google Scholar]
- Crook, M.; Ennos, A. Stem and root characteristics associated with lodging resistance in four winter wheat cultivars. J. Agric. Sci. 1994, 123, 167–174. [Google Scholar] [CrossRef]
- Xie, J.; Li, J.; Wei, F.; Wu, H.; Yu, S.; Wang, L.; Ma, H.; Shen, Y. The analysis of culm lodging resistance in main wheat cultivars in the plain of the Yangtze-Huai Rivers. Chin. Agric. Sci. Bull. 2009, 25, 108–111. [Google Scholar]
- Islam, M.S.; Peng, S.; Visperas, R.M.; Ereful, N.; Bhuiya, M.S.U.; Julfiquar, A. Lodging-related morphological traits of hybrid rice in a tropical irrigated ecosystem. Field Crop. Res. 2007, 101, 240–248. [Google Scholar] [CrossRef]
- Kong, E.; Liu, D.; Guo, X.; Yang, W.; Sun, J.; Li, X.; Zhan, K.; Cui, D.; Lin, J.; Zhang, A. Anatomical and chemical characteristics associated with lodging resistance in wheat. Crop J. 2013, 1, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Zhang, W.; Ding, Y.; Zhang, J.; Cambula, E.D.; Weng, F.; Liu, Z.; Ding, C.; Tang, S.; Chen, L. Shading contributes to the reduction of stem mechanical strength by decreasing cell wall synthesis in japonica rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 881. [Google Scholar] [CrossRef]
- Okuno, A.; Hirano, K.; Asano, K.; Takase, W.; Masuda, R.; Morinaka, Y.; Ueguchi-Tanaka, M.; Kitano, H.; Matsuoka, M. New approach to increasing rice lodging resistance and biomass yield through the use of high gibberellin producing varieties. PLoS ONE 2014, 9, e86870. [Google Scholar] [CrossRef]
- Hirano, K.; Ordonio, R.L.; Matsuoka, M. Engineering the lodging resistance mechanism of post-Green Revolution rice to meet future demands. Proc. Jpn. Acad. Ser. B 2017, 93, 220–233. [Google Scholar] [CrossRef] [Green Version]
- Spielmeyer, W.; Ellis, M.H.; Chandler, P.M. Semidwarf (sd-1),“green revolution” rice, contains a defective gibberellin 20-oxidase gene. Proc. Natl. Acad. Sci. 2002, 99, 9043–9048. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, S.; Gui, J.; Fu, C.; Yu, H.; Song, D.; Shen, J.; Qin, P.; Liu, X.; Han, B. Shortened basal internodes encodes a gibberellin 2-oxidase and contributes to lodging resistance in rice. Mol. Plant 2018, 11, 288–299. [Google Scholar] [CrossRef]
- Chang, L.; Lai-geng, L. Advances in molecular understanding of rice lodging resistance. Chin. J. Rice Sci. 2016, 2, 14. [Google Scholar]
- Zhang, J.; Li, G.; Huang, Q.; Liu, Z.; Ding, C.; Tang, S.; Chen, L.; Wang, S.; Ding, Y.; Zhang, W. Effects of culm carbohydrate partitioning on basal stem strength in a high-yielding rice population. Crop J. 2017, 5, 478–487. [Google Scholar] [CrossRef]
- Sarker, Z.; Shamsuddin, A.; Rahman, L.; Ara, R. Genotypic and phenotypic correlation and path analysis for lodging resistance traits in bread wheat (Triticum aestivum L.). Bangladesh J. Plant Breed. Genet. 2007, 20, 51–57. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, H.; Yi, Y.; Ding, J.; Zhu, M.; Li, C.; Guo, W.; Feng, C.; Zhu, X. Effect of nitrogen levels and nitrogen ratios on lodging resistance and yield potential of winter wheat (Triticum aestivum L.). PLoS ONE 2017, 12, e0187543. [Google Scholar] [CrossRef] [PubMed]
- Berry, P.; Spink, J.; Gay, A.P.; Craigon, J. A comparison of root and stem lodging risks among winter wheat cultivars. J. Agric. Sci. 2003, 141, 191–202. [Google Scholar] [CrossRef]
- Mariani, L.; Ferrante, A. Agronomic management for enhancing plant tolerance to abiotic stresses—drought, salinity, hypoxia, and lodging. Horticulturae 2017, 3, 52. [Google Scholar] [CrossRef]
- Zuber, U.; Winzeler, H.; Messmer, M.; Keller, M.; Keller, B.; Schmid, J.; Stamp, P. Morphological traits associated with lodging resistance of spring wheat (Triticum aestivum L.). J. Agron. Crop Sci. 1999, 182, 17–24. [Google Scholar] [CrossRef]
- Upadhya, D.; Dhakal, R.; Khadka, K.; Rana, S.; Acharya, P.; Rana, R.; Chaudhary, P. Local knowledge on climate-induced traits in rice for improving crop yield, food security and climate resilience. Int. Agric. Innov. Res. J. 2016, 5, 385–396. [Google Scholar]
- Kashiwagi, T.; Togawa, E.; Hirotsu, N.; Ishimaru, K. Improvement of lodging resistance with QTLs for stem diameter in rice (Oryza sativa L.). Theor. Appl. Genet. 2008, 117, 749–757. [Google Scholar] [CrossRef]
- Ookawa, T.; Hobo, T.; Yano, M.; Murata, K.; Ando, T.; Miura, H.; Asano, K.; Ochiai, Y.; Ikeda, M.; Nishitani, R. New approach for rice improvement using a pleiotropic QTL gene for lodging resistance and yield. Nat. Commun. 2010, 1, 132. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Tan, L.; Fu, Y.; Liu, F.; Cai, H.; Xie, D.; Wu, F.; Wu, J.; Matsumoto, T.; Sun, C. Genetic control of inflorescence architecture during rice domestication. Nat. Commun. 2013, 4, 2200. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Wei, J.; Nie, L.; Chen, F. Study on the genetic model stem characters in wheat. J. Henan Agric. Sci. 2002, 9, 4–7. [Google Scholar]
- Xiang, D.; Song, Y.; Wu, Q.; Ma, C.; Zhao, J.; Wan, Y.; Zhao, G. Relationship between stem characteristics and lodging resistance of Tartary buckwheat (Fagopyrum tataricum). Plant Prod. Sci. 2019, 22, 202–210. [Google Scholar] [CrossRef]
- Tripathi, S.C.; Sayre, K.; Kaul, J.; Narang, R. Growth and morphology of spring wheat (Triticum aestivum L.) culms and their association with lodging: Effects of genotypes, N levels and ethephon. Field Crop. Res. 2003, 84, 271–290. [Google Scholar] [CrossRef]
- Zheng, M.; Chen, J.; Shi, Y.; Li, Y.; Yin, Y.; Yang, D.; Luo, Y.; Pang, D.; Xu, X.; Li, W. Manipulation of lignin metabolism by plant densities and its relationship with lodging resistance in wheat. Sci. Rep. 2017, 7, 41805. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Luan, S.; Zhang, L.; Liu, Y.; Zhang, L.; Li, H. Penalties in yield and yield associated traits caused by stem lodging at different developmental stages in summer and spring foxtail millet cultivars. Field Crop. Res. 2018, 217, 104–112. [Google Scholar] [CrossRef]
- Yadav, S.; Singh, U.M.; Naik, S.M.; Venkateshwarlu, C.; Ramayya, P.J.; Raman, K.A.; Sandhu, N.; Kumar, A. Molecular mapping of QTLs associated with lodging resistance in dry direct-seeded rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 1431. [Google Scholar] [CrossRef]
- Wang, D.; Ding, W.; Feng, S.; Hu, T.; Li, G.; Li, X.; Yang, Y.; Ru, Z. Stem characteristics of different wheat varieties and its relationship with lodging-resistance. Ying Yong Sheng Tai Xue Bao=J. Appl. Ecol. 2016, 27, 1496–1502. [Google Scholar]
- Kono, M. Physiological aspects of lodging. In Science of the Rice Plant, Vol. 2, Physiology, Vol. 2; Matsuo, T., Kumazawa, K., Ishii, R., Ishihara, K., Hirata, H., Eds.; Food and Agriculture Policy Research Center: Tokyo, Japan, 1995; pp. 971–982. [Google Scholar]
- Xiao, Y.; Liu, J.; Li, H.; Cao, X.; Xia, X.; Zhonghu, H. Lodging resistance and yield potential of winter wheat: Effect of planting density and genotype. Front. Agric. Sci. Eng. 2015, 2, 168–178. [Google Scholar] [CrossRef]
- Xu, M.; Quan, X.; Shi, T.; Zheng, C.; Liu, X. Study on conducting bundle character of neck and correlation of several rice breeds. J. Agric. Sci. Yanbian Univ. 2000, 22, 81–85. [Google Scholar]
- Yan, C.-J.; Zhou, J.-H.; Yan, S.; Chen, F.; Yeboah, M.; Tang, S.-Z.; Liang, G.-H.; Gu, M.-H. Identification and characterization of a major QTL responsible for erect panicle trait in japonica rice (Oryza sativa L.). Theor. Appl. Genet. 2007, 115, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, M.R.; Rahimi-Ajdadi, F.; Dabbaghi, A. Cutting energy of rice stem as influenced by internode position and dimensional characteristics of different varieties. Aust. J. Crop Sci. 2011, 5, 681. [Google Scholar]
- Gowda, V.R.; Henry, A.; Yamauchi, A.; Shashidhar, H.; Serraj, R. Root biology and genetic improvement for drought avoidance in rice. Field Crop. Res. 2011, 122, 1–13. [Google Scholar] [CrossRef]
- Atkinson, J.A.; Rasmussen, A.; Traini, R.; Voß, U.; Sturrock, C.; Mooney, S.J.; Wells, D.M.; Bennett, M.J. Branching out in roots: Uncovering form, function, and regulation. Plant Physiol. 2014, 166, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Apweiler, R.; Martin, M.J.; O’Donovan, C.; Magrane, M.; Alam-Faruque, Y.; Antunes, R.; Barrell, D.; Bely, B.; Bingley, M.; Binns, D. The universal protein resource (UniProt) in 2010. Nucleic Acids Res. 2010, 38, D142–D148. [Google Scholar]
- Siddique, K.; Chen, Y.; Rengel, Z. Efficient root system for abiotic stress tolerance in crops. Procedia Environ. Sci. 2015, 29, 295. [Google Scholar] [CrossRef]
- Terashima, K.; Akita, S.; Sakai, N. Eco-physiological Characteristics Related with Lodging Tolerance of Rice in Direct Sowing Cultivation: III. Relationship between the characteristics of root distribution in the soil and lodging tolerance. Jpn. J. Crop Sci. 1995, 64, 243–250. [Google Scholar] [CrossRef]
- Terashima, K. Eco-physiological study of root lodging tolerance in direct-seeded rice [Oryza sativa] cultivars. JARQ 1997, 31, 155–162. [Google Scholar]
- Ennos, A.R. The mechanics of anchorage in wheat Triticum aestivum L.: II. Anchorage of mature wheat against lodging. J. Exp. Bot. 1991, 42, 1607–1613. [Google Scholar] [CrossRef]
- Crook, M.J.; Ennos, A. The mechanics of root lodging in winter wheat, Triticum aestivum L. J. Exp. Bot. 1993, 44, 1219–1224. [Google Scholar] [CrossRef]
- Yamauchi, T.; Abe, F.; Kawaguchi, K.; Oyanagi, A.; Nakazono, M. Adventitious roots of wheat seedlings that emerge in oxygen-deficient conditions have increased root diameters with highly developed lysigenous aerenchyma. Plant Signal. Behav. 2014, 9, e28506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamauchi, T.; Shimamura, S.; Nakazono, M.; Mochizuki, T. Aerenchyma formation in crop species: A review. Field Crop. Res. 2013, 152, 8–16. [Google Scholar] [CrossRef]
- Pinthus, M.J.; Rudich, J. Increase in Grain Yield of CCC-Treated Wheat (Triticum Aestivum) in the Absence of Lodging. Agrochimica 1967, 11, 565–570. [Google Scholar]
- Berry, P.; Griffin, J.; Sylvester-Bradley, R.; Scott, R.; Spink, J.; Baker, C.; Clare, R. Controlling plant form through husbandry to minimise lodging in wheat. Field Crop. Res. 2000, 67, 59–81. [Google Scholar] [CrossRef]
- Pinthus, M.J. Spread of the root system as indicator for evaluating lodging resistance of wheat 1. Crop Sci. 1967, 7, 107–110. [Google Scholar] [CrossRef]
- Sterling, M.; Baker, C.; Joseph, G.; Gillmeier, S.; Mohammadi, M.; D Blackburn, G.; Wyatt, D.; Berry, P.; Hatley, D.; Spink, J. Mitigating yield losses due to lodging of cereal crops. In Proceedings of the International Workshop on Wind-Related Disasters and Mitigation Tohoku University, Sendai, Japan, 11–14 March 2018. [Google Scholar]
- Morita, S.; Suga, T.; Yamazaki, K. The relationship between root length density and yield in rice plants. Jpn. J. Crop Sci. 1988, 57, 438–443. [Google Scholar] [CrossRef]
- Izumi, Y.; Kono, Y.; Yamauchi, A.; Iijima, M. Timecourse changes in two different topological indices with seminal root system development of rice [Oryza sativa]. Jpn. J. Crop Sci. 1996, 65, 303–308. [Google Scholar] [CrossRef]
- Abe, J.; Morita, S. Growth direction of nodal roots in rice: Its variation and contribution to root system formation. Plant Soil 1994, 165, 333–337. [Google Scholar] [CrossRef]
- Ogata, T.; Matsue, Y. Studies on Direct Sowing Culture of Rice in Northern Kyushu: I. Evaluation of lodging tolerance. Jpn. J. Crop Sci. 1996, 65, 87–92. [Google Scholar] [CrossRef]
- Zhu, C.; Cheng, W.; Sakai, H.; Oikawa, S.; Laza, R.C.; Usui, Y.; Hasegawa, T. Effects of elevated [CO 2] on stem and root lodging among rice cultivars. Chin. Sci. Bull. 2013, 58, 1787–1794. [Google Scholar] [CrossRef]
- Kang, S.-Y.; Yang, W.-H.; Shin, H.-T. Evaluation of Root Characters Associated with Lodging Tolerance by Seedling Test in Rice. Korean J. Crop Sci. 1999, 44, 309–315. [Google Scholar]
- Chen, X.; Shi, C.; Yin, Y.; Wang, Z.; Shi, Y.; Peng, D.; Ni, Y.; Cai, T. Relationship between lignin metabolism and lodging resistance in wheat. Acta Agron. Sin. 2011, 37, 1616–1622. [Google Scholar] [CrossRef]
- Del Río, J.C.; Rencoret, J.; Prinsen, P.; Martínez, A. n. T.; Ralph, J.; Gutiérrez, A. Structural characterization of wheat straw lignin as revealed by analytical pyrolysis, 2D-NMR, and reductive cleavage methods. J. Agric. Food Chem. 2012, 60, 5922–5935. [Google Scholar] [CrossRef] [PubMed]
- Hyles, J.; Vautrin, S.; Pettolino, F.; MacMillan, C.; Stachurski, Z.; Breen, J.; Berges, H.; Wicker, T.; Spielmeyer, W. Repeat-length variation in a wheat cellulose synthase-like gene is associated with altered tiller number and stem cell wall composition. J. Exp. Bot. 2017, 68, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Knapp, J.; Harms, C.; Volenec, J. Growth Regulator Effects on Wheat Culm Nonstructural and Structural Carbohydrates and Lignin 1. Crop Sci. 1987, 27, 1201–1205. [Google Scholar] [CrossRef]
- Li, Y.; Qian, Q.; Zhou, Y.; Yan, M.; Sun, L.; Zhang, M.; Fu, Z.; Wang, Y.; Han, B.; Pang, X. BRITTLE CULM1, which encodes a COBRA-like protein, affects the mechanical properties of rice plants. Plant Cell 2003, 15, 2020–2031. [Google Scholar] [CrossRef] [PubMed]
- Khan, W.; Prithiviraj, B.; Smith, D.L. Chitosan and chitin oligomers increase phenylalanine ammonia-lyase and tyrosine ammonia-lyase activities in soybean leaves. J. Plant Physiol. 2003, 160, 859–863. [Google Scholar] [CrossRef]
- Nguyen, T.-N.; Son, S.; Jordan, M.C.; Levin, D.B.; Ayele, B.T. Lignin biosynthesis in wheat (Triticum aestivum L.): Its response to waterlogging and association with hormonal levels. BMC Plant Biol. 2016, 16, 28. [Google Scholar] [CrossRef]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef]
- Bi, C.; Chen, F.; Jackson, L.; Gill, B.S.; Li, W. Expression of lignin biosynthetic genes in wheat during development and upon infection by fungal pathogens. Plant Mol. Biol. Report. 2011, 29, 149–161. [Google Scholar] [CrossRef]
- Ma, Q.-H. The expression of caffeic acid 3-O-methyltransferase in two wheat genotypes differing in lodging resistance. J. Exp. Bot. 2009, 60, 2763–2771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.-H. Characterization of a cinnamoyl-CoA reductase that is associated with stem development in wheat. J. Exp. Bot. 2007, 58, 2011–2021. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.-H. Functional analysis of a cinnamyl alcohol dehydrogenase involved in lignin biosynthesis in wheat. J. Exp. Bot. 2010, 61, 2735–2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadeem, M.; Li, J.; Yahya, M.; Wang, M.; Ali, A.; Cheng, A.; Wang, X.; Ma, C. Grain legumes and fear of salt stress: Focus on mechanisms and management strategies. Int. J. Mol. Sci. 2019, 20, 799. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Li, J.; Wang, M.; Shah, L.; Lu, S.; Wang, X.; Ma, C. Unraveling field crops sensitivity to heat stress: Mechanisms, approaches, and future prospects. Agronomy 2018, 8, 128. [Google Scholar] [CrossRef]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research Progress and Perspective on Drought Stress in Legumes: A Review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef]
- Gutiérrez, J.; Núñez-Flores, M.J.L.; Gómez-Ros, L.V.; Uzal, E.N.; Carrasco, A.E.; Díaz, J.; Sottomayor, M.; Cuello, J.; Barceló, A.R. Hormonal regulation of the basic peroxidase isoenzyme from Zinnia elegans. Planta 2009, 230, 767–778. [Google Scholar] [CrossRef]
- Tamaoki, D.; Karahara, I.; Nishiuchi, T.; Wakasugi, T.; Yamada, K.; Kamisaka, S. Involvement of auxin dynamics in hypergravity-induced promotion of lignin-related gene expression in Arabidopsis inflorescence stems. J. Exp. Bot. 2011, 62, 5463–5469. [Google Scholar] [CrossRef] [Green Version]
- Gallego-Giraldo, L.; Escamilla-Trevino, L.; Jackson, L.A.; Dixon, R.A. Salicylic acid mediates the reduced growth of lignin down-regulated plants. Proc. Natl. Acad. Sci. USA 2011, 108, 20814–20819. [Google Scholar] [CrossRef] [Green Version]
- Meisrimler, C.-N.; Buck, F.; Lüthje, S. Alterations in soluble Class III peroxidases of maize shoots by flooding stress. Proteomes 2014, 2, 303–322. [Google Scholar] [CrossRef]
- Musgrave, M.E. Waterlogging effects on yield and photosynthesis in eight winter wheat cultivars. Crop Sci. 1994, 34, 1314–1318. [Google Scholar] [CrossRef]
- Albrecht, G.; Kammerer, S.; Praznik, W.; Wiedenroth, E. Fructan content of wheat seedlings (Triticum aestivum L.) under hypoxia and following re-aeration. New Phytol. 1993, 123, 471–476. [Google Scholar] [CrossRef]
- Huang, B.; Johnson, J.W. Root respiration and carbohydrate status of two wheat genotypes in response to hypoxia. Ann. Bot. 1995, 75, 427–432. [Google Scholar] [CrossRef]
- Huang, B.; Johnson, J.W.; Box, J.E.; NeSmith, D.S. Root characteristics and hormone activity of wheat in response to hypoxia and ethylene. Crop Sci. 1997, 37, 812–818. [Google Scholar] [CrossRef]
- Vanholme, R.; Storme, V.; Vanholme, B.; Sundin, L.; Christensen, J.H.; Goeminne, G.; Halpin, C.; Rohde, A.; Morreel, K.; Boerjan, W. A systems biology view of responses to lignin biosynthesis perturbations in Arabidopsis. Plant Cell 2012, 24, 3506–3529. [Google Scholar] [CrossRef] [PubMed]
- Lastdrager, J.; Hanson, J.; Smeekens, S. Sugar signals and the control of plant growth and development. J. Exp. Bot. 2014, 65, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Usadel, B.; Bläsing, O.E.; Gibon, Y.; Retzlaff, K.; Höhne, M.; Günther, M.; Stitt, M. Global transcript levels respond to small changes of the carbon status during progressive exhaustion of carbohydrates in Arabidopsis rosettes. Plant Physiol. 2008, 146, 1834–1861. [Google Scholar] [CrossRef]
- Ke, S.; Luan, X.; Liang, J.; Hung, Y.-H.; Hsieh, T.-F.; Zhang, X.-Q. Rice OsPEX1, an extensin-like protein, affects lignin biosynthesis and plant growth. Plant Mol. Biol. 2019, 100, 151–161. [Google Scholar] [CrossRef]
- Miyamoto, T.; Takada, R.; Tobimatsu, Y.; Takeda, Y.; Suzuki, S.; Yamamura, M.; Osakabe, K.; Osakabe, Y.; Sakamoto, M.; Umezawa, T. Os MYB 108 loss-of-function enriches p-coumaroylated and tricin lignin units in rice cell walls. Plant J. 2019, 96, 975–987. [Google Scholar] [CrossRef]
- Si, T.; Wang, X.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.; Jiang, D. Double benefits of mechanical wounding in enhancing chilling tolerance and lodging resistance in wheat plants. Plant Biol. 2019, 21, 813–824. [Google Scholar] [CrossRef]
- Cai, T.; Meng, X.; Liu, X.; Liu, T.; Wang, H.; Jia, Z.; Yang, D.; Ren, X. Exogenous hormonal application regulates the occurrence of wheat tillers by changing endogenous hormones. Front. Plant Sci. 2018, 9, 1886. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gu, D.; Ding, Y.; Wang, Q.; Li, G.; Wang, S. The relationship between nitrogen, auxin and cytokinin in the growth regulation of rice (‘Oryza sativa’l.) Tiller buds. Aust. J. Crop Sci. 2011, 5, 1019. [Google Scholar]
- Chatfield, S.P.; Stirnberg, P.; Forde, B.G.; Leyser, O. The hormonal regulation of axillary bud growth in Arabidopsis. Plant J. 2000, 24, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Genschik, P. Releasing the brakes of plant growth: How GAs shutdown DELLA proteins. J. Exp. Bot. 2009, 60, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.M.; Singh, D.P. Tall tales from sly dwarves: Novel functions of gibberellins in plant development. Trends Plant Sci. 2005, 10, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-H.; Kang, K.; Lee, S.-H.; Lee, I.-J.; Paek, N.-C. OsWOX3A is involved in negative feedback regulation of the gibberellic acid biosynthetic pathway in rice (Oryza sativa). J. Exp. Bot. 2016, 67, 1677–1687. [Google Scholar] [CrossRef]
- Yin, C.-C.; Zhao, H.; Ma, B.; Chen, S.-Y.; Zhang, J.-S. Diverse roles of ethylene in regulating agronomic traits in rice. Front. Plant Sci. 2017, 8, 1676. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Nagai, K.; Furukawa, S.; Song, X.-J.; Kawano, R.; Sakakibara, H.; Wu, J.; Matsumoto, T.; Yoshimura, A.; Kitano, H. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature 2009, 460, 1026. [Google Scholar] [CrossRef]
- Fukao, T.; Yeung, E.; Bailey-Serres, J. The submergence tolerance gene SUB1A delays leaf senescence under prolonged darkness through hormonal regulation in rice. Plant Physiol. 2012, 160, 1795–1807. [Google Scholar] [CrossRef]
- Qi, W.; Sun, F.; Wang, Q.; Chen, M.; Huang, Y.; Feng, Y.-Q.; Luo, X.; Yang, J. Rice ethylene-response AP2/ERF factor OsEATB restricts internode elongation by down-regulating a gibberellin biosynthetic gene. Plant Physiol. 2011, 157, 216–228. [Google Scholar] [CrossRef]
- Yukiyoshi, K.; Karahara, I. Role of ethylene signalling in the formation of constitutive aerenchyma in primary roots of rice. AoB Plants 2014, 6, plu043. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Drummond, B.J.; Habben, J.E.; Brugire, N.; Weers, B.P.; Hakimi, S.M.; Lafitte, H.R.; Schussler, J.R.; Mo, H.; Beatty, M. Ectopic expression of ARGOS 8 reveals a role for ethylene in root-lodging resistance in maize. Plant J. 2019, 97, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Divte, P.; Yadav, P.; kumar Jain, P.; Paul, S.; Singh, B. Ethylene regulation of root growth and phytosiderophore biosynthesis determines iron deficiency tolerance in wheat (Triticum spp). Environ. Exp. Bot. 2019, 162, 1–13. [Google Scholar] [CrossRef]
- Yano, K.; Ookawa, T.; Aya, K.; Ochiai, Y.; Hirasawa, T.; Ebitani, T.; Takarada, T.; Yano, M.; Yamamoto, T.; Fukuoka, S. Isolation of a novel lodging resistance QTL gene involved in strigolactone signaling and its pyramiding with a QTL gene involved in another mechanism. Mol. Plant 2015, 8, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Ookawa, T.; Aoba, R.; Yamamoto, T.; Ueda, T.; Takai, T.; Fukuoka, S.; Ando, T.; Adachi, S.; Matsuoka, M.; Ebitani, T. Precise estimation of genomic regions controlling lodging resistance using a set of reciprocal chromosome segment substitution lines in rice. Sci. Rep. 2016, 6, 30572. [Google Scholar] [CrossRef]
- Schmid, J.; Winzeler, H. Genetic studies of crosses between common wheat (Triticum aestivum L.) and spelt (Triticum spelta L.). J. Genet. Breed. 1990, 44, 75–80. [Google Scholar]
- Keller, M.; Karutz, C.; Schmid, J.; Stamp, P.; Winzeler, M.; Keller, B.; Messmer, M. Quantitative trait loci for lodging resistance in a segregating wheat× spelt population. Theor. Appl. Genet. 1999, 98, 1171–1182. [Google Scholar] [CrossRef]
- Verma, V.; Worland, A.; Savers, E.; Fish, L.; Caligari, P.; Snape, J. Identification and characterization of quantitative trait loci related to lodging resistance and associated traits in bread wheat. Plant Breed. 2005, 124, 234–241. [Google Scholar] [CrossRef]
- Hai, L.; Guo, H.; Xiao, S.; Jiang, G.; Zhang, X.; Yan, C.; Xin, Z.; Jia, J. Quantitative trait loci (QTL) of stem strength and related traits in a doubled-haploid population of wheat (Triticum aestivum L.). Euphytica 2005, 141, 1–9. [Google Scholar] [CrossRef]
- Ellis, M.; Rebetzke, G.; Azanza, F.; Richards, R.; Spielmeyer, W. Molecular mapping of gibberellin-responsive dwarfing genes in bread wheat. Theor. Appl. Genet. 2005, 111, 423–430. [Google Scholar] [CrossRef]
- Li, X.; Xia, X.; Xiao, Y.; He, Z.; Wang, D.; Trethowan, R.; Wang, H.; Chen, X. QTL mapping for plant height and yield components in common wheat under water-limited and full irrigation environments. Crop Pasture Sci. 2015, 66, 660–670. [Google Scholar] [CrossRef]
- Griffiths, S.; Simmonds, J.; Leverington, M.; Wang, Y.; Fish, L.; Sayers, L.; Alibert, L.; Orford, S.; Wingen, L.; Snape, J. Meta-QTL analysis of the genetic control of crop height in elite European winter wheat germplasm. Mol. Breed. 2012, 29, 159–171. [Google Scholar] [CrossRef]
- Tian, X.; Wen, W.; Xie, L.; Fu, L.; Xu, D.; Fu, C.; Wang, D.; Chen, X.; Xia, X.; Chen, Q. Molecular mapping of reduced plant height gene Rht24 in bread wheat. Front. Plant Sci. 2017, 8, 1379. [Google Scholar] [CrossRef] [PubMed]
- Spielmeyer, W.; Hyles, J.; Joaquim, P.; Azanza, F.; Bonnett, D.; Ellis, M.; Moore, C.; Richards, R. A QTL on chromosome 6A in bread wheat (Triticum aestivum) is associated with longer coleoptiles, greater seedling vigour and final plant height. Theor. Appl. Genet. 2007, 115, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Xu, Y.; Liu, H.; Mao, Z.; Zhang, C.; Ma, Y.; Zhang, Q.; Meng, Z.; Chong, K. The interaction between OsMADS57 and OsTB1 modulates rice tillering via DWARF14. Nat. Commun. 2013, 4, 1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashiwagi, T.; Ishimaru, K. Identification and functional analysis of a locus for improvement of lodging resistance in rice. Plant Physiol. 2004, 134, 676–683. [Google Scholar] [CrossRef]
- Mu, P.; Li, Z.; Li, C.; Zhang, H.; Wang, X. QTL analysis for lodging resistance in rice using a DH population under lowland and upland ecosystems. Yi Chuan Xue Bao=Acta Genet. Sin. 2004, 31, 717–723. [Google Scholar]
- Matsubara, K.; Yamamoto, E.; Kobayashi, N.; Ishii, T.; Tanaka, J.; Tsunematsu, H.; Yoshinaga, S.; Matsumura, O.; Yonemaru, J.-I.; Mizobuchi, R. Improvement of rice biomass yield through QTL-based selection. PLoS ONE 2016, 11, e0151830. [Google Scholar] [CrossRef]
- Arai-Sanoh, Y.; Takai, T.; Yoshinaga, S.; Nakano, H.; Kojima, M.; Sakakibara, H.; Kondo, M.; Uga, Y. Deep rooting conferred by DEEPER ROOTING 1 enhances rice yield in paddy fields. Sci. Rep. 2014, 4, 5563. [Google Scholar] [CrossRef] [Green Version]
- Rajaram, S. Prospects and promise of wheat breeding in the 21 st century. In Wheat in a Global Environment; Springer: Berlin/Heidelberg, Germany, 2001; pp. 37–52. [Google Scholar]
- Pearce, S.; Saville, R.; Vaughan, S.P.; Chandler, P.M.; Wilhelm, E.P.; Sparks, C.A.; Al-Kaff, N.; Korolev, A.; Boulton, M.I.; Phillips, A.L. Molecular characterization of Rht-1 dwarfing genes in hexaploid wheat. Plant Physiol. 2011, 157, 1820–1831. [Google Scholar] [CrossRef]
- Wilhelm, E.P.; Howells, R.M.; Al-Kaff, N.; Jia, J.; Baker, C.; Leverington-Waite, M.A.; Griffiths, S.; Greenland, A.J.; Boulton, M.I.; Powell, W. Genetic characterization and mapping of the Rht-1 homoeologs and flanking sequences in wheat. Theor. Appl. Genet. 2013, 126, 1321–1336. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Richards, D.E.; Hartley, N.M.; Murphy, G.P.; Devos, K.M.; Flintham, J.E.; Beales, J.; Fish, L.J.; Worland, A.J.; Pelica, F. Green revolution’genes encode mutant gibberellin response modulators. Nature 1999, 400, 256. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Smith, S.M.; Ayele, M.; Yang, L.; Jogi, A.; Chaluvadi, S.R.; Bennetzen, J.L. High-throughput discovery of mutations in tef semi-dwarfing genes by next-generation sequencing analysis. Genetics 2012, 192, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Jamali, K.; Ali, S.A. Yield and yield components with relation to plant height in semi-dwarf wheat. Pak. J. Bot. 2008, 40, 1805–1808. [Google Scholar]
- Radley, M. Comparison of endogenous gibberellins and response to applied gibberellin of some dwarf and tall wheat cultivars. Planta 1970, 92, 292–300. [Google Scholar] [CrossRef] [PubMed]
- King, R.; Gale, M.; Quarrie, S. Effects of Norin 10 and Tom Thumb dwarfing genes on morphology, physiology and abscisic acid production in wheat. Ann. Bot. 1983, 51, 201–208. [Google Scholar] [CrossRef]
- Flintham, J.; Börner, A.; Worland, A.; Gale, M. Optimizing wheat grain yield: Effects of Rht (gibberellin-insensitive) dwarfing genes. J. Agric. Sci. 1997, 128, 11–25. [Google Scholar] [CrossRef]
- Amram, A.; Fadida-Myers, A.; Golan, G.; Nashef, K.; Ben-David, R.; Peleg, Z. Effect of GA-sensitivity on wheat early vigor and yield components under deep sowing. Front. Plant Sci. 2015, 6, 487. [Google Scholar] [CrossRef] [Green Version]
- Shearman, V.J. Changes in the Yield Limiting Processes Associated with the Genetic Improvement of Wheat. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 2001. [Google Scholar]
- Rebetzke, G.; Ellis, M.; Bonnett, D.; Mickelson, B.; Condon, A.; Richards, R. Height reduction and agronomic performance for selected gibberellin-responsive dwarfing genes in bread wheat (Triticum aestivum L.). Field Crop. Res. 2012, 126, 87–96. [Google Scholar] [CrossRef]
- Daoura, B.G.; Chen, L.; Du, Y.; Hu, Y.-G. Genetic effects of dwarfing gene Rht-5 on agronomic traits in common wheat (Triticum aestivum L.) and QTL analysis on its linked traits. Field Crop. Res. 2014, 156, 22–29. [Google Scholar] [CrossRef]
- Daoura, B.G.; Chen, L.; Hu, Y.-G. Agronomic traits affected by dwarfing gene’Rht-5′in common wheat (‘Triticum aestivum’L.). Aust. J. Crop Sci. 2013, 7, 1270. [Google Scholar]
- Worland, A.; Law, C.; Shakoor, A. The genetical analysis of an induced height mutant in wheat. Heredity 1980, 45, 61. [Google Scholar] [CrossRef]
- Korzun, V.; Röder, M.; Ganal, M.; Worland, A.; Law, C. Genetic analysis of the dwarfing gene (Rht8) in wheat. Part I. Molecular mapping of Rht8 on the short arm of chromosome 2D of bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 1998, 96, 1104–1109. [Google Scholar] [CrossRef]
- Würschum, T.; Leiser, W.L.; Weissmann, S.; Maurer, H.P. Genetic architecture of male fertility restoration of Triticum timopheevii cytoplasm and fine-mapping of the major restorer locus Rf3 on chromosome 1B. Theor. Appl. Genet. 2017, 130, 1253–1266. [Google Scholar] [CrossRef] [PubMed]
- Gasperini, D.; Greenland, A.; Hedden, P.; Dreos, R.; Harwood, W.; Griffiths, S. Genetic and physiological analysis of Rht8 in bread wheat: An alternative source of semi-dwarfism with a reduced sensitivity to brassinosteroids. J. Exp. Bot. 2012, 63, 4419. [Google Scholar] [PubMed]
- Wang, Y.; Li, J. Molecular basis of plant architecture. Annu. Rev. Plant Biol. 2008, 59, 253–279. [Google Scholar] [CrossRef]
- Botwright, T.L.; Rebetzke, G.J.; Condon, A.G.; Richards, R.A. Influence of the gibberellin-sensitive Rht8 dwarfing gene on leaf epidermal cell dimensions and early vigour in wheat (Triticum aestivum L.). Ann. Bot. 2005, 95, 631–639. [Google Scholar] [CrossRef]
- Rebetzke, G.; Richards, R.; Fettell, N.; Long, M.; Condon, A.G.; Forrester, R.; Botwright, T. Genotypic increases in coleoptile length improves stand establishment, vigour and grain yield of deep-sown wheat. Field Crop. Res. 2007, 100, 10–23. [Google Scholar] [CrossRef]
- Worland, A.; Korzun, V.; Röder, M.; Ganal, M.; Law, C. Genetic analysis of the dwarfing gene Rht8 in wheat. Part II. The distribution and adaptive significance of allelic variants at the Rht8 locus of wheat as revealed by microsatellite screening. Theor. Appl. Genet. 1998, 96, 1110–1120. [Google Scholar] [CrossRef]
- Konzak, C.F. Mutations and mutation breeding. In Wheat and wheat improvement, 2nd ed.; Heyne, E.G., Ed.; American Society of Agronomy, Inc.; Crop Science Society of America, Inc.; Soil Science Society of America, Inc.: Madison, WI, USA, 1987; pp. 428–443. [Google Scholar]
- Haque, M.; Martinek, P.; Watanabe, N.; Kuboyama, T. Genetic mapping of gibberellic acid-sensitive genes for semi-dwarfism in durum wheat. Cereal Res. Commun. 2011, 39, 171–178. [Google Scholar] [CrossRef]
- Vikhe, P.; Patil, R.; Chavan, A.; Oak, M.; Tamhankar, S. Mapping gibberellin-sensitive dwarfing locus Rht18 in durum wheat and development of SSR and SNP markers for selection in breeding. Mol. Breed. 2017, 37, 28. [Google Scholar] [CrossRef]
- Peng, J.H.; Sun, D.; Nevo, E. Domestication evolution, genetics and genomics in wheat. Mol. Breed. 2011, 28, 281. [Google Scholar] [CrossRef]
- Ma, Q.H.; Xu, Y.; Lin, Z.B.; He, P. Cloning of cDNA encoding COMT from wheat which is differentially expressed in lodging-sensitive and-resistant cultivars. J. Exp. Bot. 2002, 53, 2281–2282. [Google Scholar] [CrossRef] [PubMed]
- Asano, K.; Takashi, T.; Miura, K.; Qian, Q.; Kitano, H.; Matsuoka, M.; Ashikari, M. Genetic and molecular analysis of utility of sd1 alleles in rice breeding. Breed. Sci. 2007, 57, 53–58. [Google Scholar] [CrossRef]
- Liu, Q.; Ma, J.; Zhao, Q.; Zhou, X. Physical Traits Related to Rice Lodging Resistance under Different Simplified-Cultivation Methods. Agron. J. 2018, 110, 127–132. [Google Scholar] [CrossRef]
- Ashikari, M.; Sasaki, A.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Datta, S.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S. Loss-of-function of a rice gibberellin biosynthetic gene, GA20 oxidase (GA20ox-2), led to the rice ‘green revolution’. Breed. Sci. 2002, 52, 143–150. [Google Scholar] [CrossRef]
- Wang, L.; Guo, K.; Li, Y.; Tu, Y.; Hu, H.; Wang, B.; Cui, X.; Peng, L. Expression profiling and integrative analysis of the CESA/CSL superfamily in rice. BMC Plant Biol. 2010, 10, 282. [Google Scholar] [CrossRef]
- Kotake, T.; Aohara, T.; Hirano, K.; Sato, A.; Kaneko, Y.; Tsumuraya, Y.; Takatsuji, H.; Kawasaki, S. Rice Brittle culm 6 encodes a dominant-negative form of CesA protein that perturbs cellulose synthesis in secondary cell walls. J. Exp. Bot. 2011, 62, 2053–2062. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.; Peng, Z.; Peng, Q.; Cai, Q.; Peng, W.; Chen, M.; Yao, J. Regulation of plant height in rice by the Polycomb group genes OsEMF2b, OsFIE2 and OsCLF. Plant Sci. 2018, 267, 157–167. [Google Scholar] [CrossRef]
- Yang, J.; Lee, S.; Hang, R.; Kim, S.R.; Lee, Y.S.; Cao, X.; Amasino, R.; An, G. O s VIL 2 functions with PRC 2 to induce flowering by repressing O s LFL 1 in rice. Plant J. 2013, 73, 566–578. [Google Scholar] [CrossRef]
- Conrad, L.J.; Khanday, I.; Johnson, C.; Guiderdoni, E.; An, G.; Vijayraghavan, U.; Sundaresan, V. The polycomb group gene EMF 2B is essential for maintenance of floral meristem determinacy in rice. Plant J. 2014, 80, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Gu, F.; Dong, S.; Liu, W.; Wang, H.; Chen, Z.; Wang, J. CONSTANS-like 9 (COL9) delays the flowering time in Oryza sativa by repressing the Ehd1 pathway. Biochem. Biophys. Res. Commun. 2016, 479, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.-H.; Luo, H.-R. Biochemical characterization of caffeoyl coenzyme A 3-O-methyltransferase from wheat. Planta 2015, 242, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Berry, P.; Sylvester-Bradley, R.; Berry, S. Ideotype design for lodging-resistant wheat. Euphytica 2007, 154, 165–179. [Google Scholar] [CrossRef]
- Dai, X.; Wang, Y.; Dong, X.; Qian, T.; Yin, L.; Dong, S.; Chu, J.; He, M. Delayed sowing can increase lodging resistance while maintaining grain yield and nitrogen use efficiency in winter wheat. Crop J. 2017, 5, 541–552. [Google Scholar] [CrossRef]
- Spink, J.; Semere, T.; Sparkes, D.; Whaley, J.; Foulkes, M.; Clare, R.; Scott, R. Effect of sowing date on the optimum plant density of winter wheat. Ann. Appl. Biol. 2000, 137, 179–188. [Google Scholar] [CrossRef]
- Groth, D.; Lee, F. Rice: Origin, History, Technology, and Production; Smith, C.W., Dilday, R.H., Eds.; John Wiley Sons: New York, NY, USA, 2002; pp. 413–436. [Google Scholar]
- Stapper, M.; Fischer, R. Genotype, sowing date and plant spacing influence on high-yielding irrigated wheat in southern New South Wales. II. Growth, yield and nitrogen use. Aust. J. Agric. Res. 1990, 41, 1021–1041. [Google Scholar] [CrossRef]
- Kirby, E.; Appleyard, M.; Fellowes, G. Leaf emergence and tillering in barley and wheat. Agronomie 1985, 5, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Craigie, R.; Brown, H.; George, M. Grain yield of winter feed wheat in response to sowing date and sowing rate. Agron. N. Z. 2015, 45, 1–8. [Google Scholar]
- Jat, R.K.; Singh, P.; Jat, M.L.; Dia, M.; Sidhu, H.S.; Jat, S.L.; Bijarniya, D.; Jat, H.S.; Parihar, C.M.; Kumar, U. Heat stress and yield stability of wheat genotypes under different sowing dates across agro-ecosystems in India. Field Crop. Res. 2018, 218, 33–50. [Google Scholar] [CrossRef]
- Thompson, R.; Quisenberry, S.; N’guessan, F.; Heagler, A.; Giesler, G. Planting date as a potential cultural method for managing the rice water weevil (Coleoptera: Curculionidae) in water-seeded rice in southwest Louisiana. J. Econ. Entomol. 1994, 87, 1318–1324. [Google Scholar] [CrossRef]
- Faghani, R.; Mobasser, H.R.; Dehpor, A.A.; Kochaksarai, S.T. The effect of planting date and seedling age on yield and yield components of rice (Oryza sativa L.) varieties in North of Iran. Afr. J. Agric. Res. 2011, 6, 2571–2575. [Google Scholar]
- Shah, A.N.; Tanveer, M.; ur Rehman, A.; Anjum, S.A.; Iqbal, J.; Ahmad, R. Lodging stress in cereal—Effects and management: An overview. Environ. Sci. Pollut. Res. 2017, 24, 5222–5237. [Google Scholar] [CrossRef] [PubMed]
- Loeppky, H.; Lafond, G.; Fowler, D. The Effect of Seeding Depth on Crown Location, Plant Development, Cold Hardiness, and Yield in Winter Wheat. Soils and Crops Workshop, 20 February 1987. [Google Scholar]
- Easson, D.; White, E.; Pickles, S. The effects of weather, seed rate and cultivar on lodging and yield in winter wheat. J. Agric. Sci. 1993, 121, 145–156. [Google Scholar] [CrossRef]
- Rajkumara, S. Lodging in cereals-A review. Agric. Rev. 2008, 29, 55–60. [Google Scholar]
- Singh, H.; Jha, G.; Rawat, A.; Babu, S.; Jha, A. Low seed rate at surface sowing enhance resilience of physiological parameters and economics of wheat (Triticum aestivum). Indian J. Agric. Sci. 2013, 83, 881–884. [Google Scholar]
- Alam, M.; Sultana, M.; Hossain, M.; Salahin, M.; Roy, U. Effect of sowing depth on the yield of spring wheat. J. Environ. Sci. Nat. Resour. 2014, 7, 277–280. [Google Scholar] [CrossRef]
- Ali, M.A.; Ali, M.; Sattar, M.; Ali, L. Sowing date effect on yield of different wheat varieties. J. Agric. Res. 2010, 48, 157–162. [Google Scholar]
- Azhiri-Sigari, T.; Gines, H.; Sebastian, L.; Wade, L. Seedling vigor of rice cultivars in response to seeding depth and soil moisture. Philipp. J. Crop Sci. 2005, 30, 53–58. [Google Scholar]
- Xiang, D.B.; Zhao, G.; Wan, Y.; Tan, M.L.; Song, C.; Song, Y. Effect of planting density on lodging-related morphology, lodging rate, and yield of tartary buckwheat (Fagopyrum tataricum). Plant Prod. Sci. 2016, 19, 479–488. [Google Scholar] [CrossRef]
- Xing, Z.; Wu, P.; Zhu, M.; Qian, H.; Cao, W.; Hu, Y.; Guo, B.; Wei, H.; Xu, K.; Dai, Q. Effect of mechanized planting methods on plant type and lodging resistance of different rice varieties. Trans. Chin. Soc. Agric. Eng. 2017, 33, 52–62. [Google Scholar]
- Yamauchi, M. Development of anaerobic direct seeding technology for rice in the tropics. In Crop Research in Asia: Achievements and Perspective, Proceedings of the 2nd Asian Crop Science Conference; Fukui Prefectural University: Fukui, Japan, 1996; pp. 198–203. [Google Scholar]
- Sun, C.; Chen, D.; Fang, J.; Wang, P.; Deng, X.; Chu, C. Understanding the genetic and epigenetic architecture in complex network of rice flowering pathways. Protein Cell 2014, 5, 889–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, R.; Mahmood, N. Impact of raised bed technology on water productivity and lodging of wheat. Pak. J. Water Resour. 2005, 9, 29–32. [Google Scholar]
- Huang, H.; Chang, Y.; Liu, X.; Hu, W.; Wu, C.; Gu, Y. The response of maize root characteristics on population density. J. South China Agric. Univ. 2014, 35, 36–41. [Google Scholar]
- Liu, S.; Song, F.; Liu, F.; Zhu, X.; Xu, H. Effect of planting density on root lodging resistance and its relationship to nodal root growth characteristics in maize (Zea mays L.). J. Agric. Sci. 2012, 4, 182. [Google Scholar] [CrossRef]
- Baloch, M.; Shah, I.; Nadim, M.; Khan, M.; Khakwani, A. Effect of seeding density and planting time on growth and yield attributes of wheat. J. Anim. Plant Sci. 2010, 20, 239–240. [Google Scholar]
- Guyer, R.; Quadranti, M. Effect of seed rate and nitrogen level on the yield of direct wet-seeded rice. In Proceedings of the 10th Asian-Pacific Weed Science Society Conference 1985, Asia Pacific Weed Science Society, Chiangmai, Thailand, 24–30 November 1985; pp. 304–311. [Google Scholar]
- Bhattacharjee, I. Note on the seed rate as cultural method of weed control for direct seeding short duration paddy (Pusa 2-21). Sci. Cult. 1978, 44, 504. [Google Scholar]
- Islam, M.; Rashid, M.; Mondal, M.; Nath, S.; Karim, M. Effect of planting density on the performance of hybrid rice (Oryza sativa L.) under waterlogged condition. Agriculturists 2013, 11, 109–113. [Google Scholar] [CrossRef]
- Hu, Y.; Cao, W.; Qian, H.; Xing, Z.; Zhang, H.; Dai, Q.G.; Huo, Z.; Xu, K.; Wei, H.; Guo, B.; et al. Effect of planting density of mechanically transplanted pot seedlings on yield, plant type and lodging resistance in rice with different panicle types. Acta Agron Sin 2015, 41, 743–757. [Google Scholar] [CrossRef]
- Roshan, N.; Azarpour, E.; Moradi, M. Study effects of different nitrogen and micronutrients fertilizers rates on yield and yield components of rice. World Appl. Sci. J. 2011, 13, 419–423. [Google Scholar]
- Sterling, M.; Baker, C.; Berry, P.; Wade, A. An experimental investigation of the lodging of wheat. Agric. For. Meteorol. 2003, 119, 149–165. [Google Scholar] [CrossRef]
- Khakwani, A.A.; Baloch, M.S.; Nadim, M.A.; Zubair, M.; Shah, I.H.; Khan, A.W. Lodging: A determining factor in reducing yield and yield structure of wheat. Sarhad J. Agric. 2010, 26, 235–239. [Google Scholar]
- Baker, C.; Berry, P.; Spink, J.; Sylvester-Bradley, R.; Griffin, J.; Scott, R.; Clare, R. A method for the assessment of the risk of wheat lodging. J. Theor. Biol. 1998, 194, 587–603. [Google Scholar] [CrossRef] [PubMed]
- Peake, A.; Huth, N.; Carberry, P.; Raine, S.; Smith, R. Quantifying potential yield and lodging-related yield gaps for irrigated spring wheat in sub-tropical Australia. Field Crop. Res. 2014, 158, 1–14. [Google Scholar] [CrossRef]
- Robins, J.; Domingo, C. Moisture and Nitrogen Effects on Irrigated Spring Wheat 1. Agron. J. 1962, 54, 135–138. [Google Scholar] [CrossRef]
- Ma, S.C.; Duan, A.W.; Ma, S.T.; Yang, S.J. Effect of Early-Stage Regulated Deficit Irrigation on Stem Lodging Resistance, Leaf Photosynthesis, Root Respiration and Yield Stability of Winter Wheat under Post-Anthesis Water Stress Conditions. Irrig. Drain. 2016, 65, 673–681. [Google Scholar] [CrossRef]
- Graham, J. Crop Lodging in British Wheats and Barleys. Ph.D. Thesis, University of Reading, Reading, UK, 1983. [Google Scholar]
- Griffin, J.M. Understanding and Assessing Lodging Risk in Winter Wheat. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 1998. [Google Scholar]
- Kong, L.; Sun, M.; Wang, F.; Liu, J.; Feng, B.; Si, J.; Zhang, B.; Li, S.; Li, H. Effects of high NH+ 4 on K+ uptake, culm mechanical strength and grain filling in wheat. Front. Plant Sci. 2014, 5, 703. [Google Scholar] [CrossRef]
- Webster, J.; Jackson, L. Management practices to reduce lodging and maximize grain yield and protein content of fall-sown irrigated hard red spring wheat. Field Crop. Res. 1993, 33, 249–259. [Google Scholar] [CrossRef]
- Crook, M.; Ennos, A. The effect of nitrogen and growth regulators on stem and root characteristics associated with lodging in two cultivars of winter wheat. J. Exp. Bot. 1995, 46, 931–938. [Google Scholar] [CrossRef]
- Nil, N.L.; Kyaw, K.W.; Hla, T.; Aye, A.T.; Mar, M.K. Evaluation of morphological traits related to lodging of rice varieties under different applied nitrogen rates in Bago (West) Region. J. Agric. Res. 2016, 3, 1–7. [Google Scholar]
- Wei, F.-Z.; Li, J.-C.; Wang, C.-Y.; Qu, H.-J.; Shen, X.-S. Effects of nitrogenous fertilizer application model on culm lodging resistance in winter wheat. Acta Agron. Sin. 2008, 34, 1080–1085. [Google Scholar] [CrossRef]
- Wang, C.; Wu Ruan, R.; Hui Yuan, X.; Hu, D.; Yang, H.; Li, Y.; Lin Yi, Z. Effects of nitrogen fertilizer and planting density on the lignin synthesis in the culm in relation to lodging resistance of buckwheat. Plant Prod. Sci. 2015, 18, 218–227. [Google Scholar] [CrossRef]
- Yang, S.; Xie, L.; Zheng, S.; Li, J.; Yuan, J. Effects of nitrogen rate and transplanting density on physical and chemical characteristics and lodging resistance of culms in hybrid rice. Acta Agron. Sin. 2009, 35, 93–103. [Google Scholar] [CrossRef]
- Zhang, J.; Li, G.; Song, Y.; Zhang, W.; Yang, C.; Wang, S.; Ding, Y. Lodging resistance of super-hybrid rice Y Liangyou 2 in two ecological regions. Acta Agron. Sin. 2013, 39, 682–692. [Google Scholar] [CrossRef]
- Peake, A.; Bell, K.; Carberry, P.; Poole, N.; Raine, S. Vegetative nitrogen stress decreases lodging risk and increases yield of irrigated spring wheat in the subtropics. Crop Pasture Sci. 2016, 67, 907–920. [Google Scholar] [CrossRef]
- Zaman, U.; Ahmad, Z.; Farooq, M.; Saeed, S.; Ahmad, M.; Wakeel, A. Potassium fertilization may improve stem strength and yield of Basmati rice grown on nitrogen-fertilized soils. Pak. J. Agric. Sci. 2015, 52. [Google Scholar]
- Yousaf, M.; Li, J.; Lu, J.; Ren, T.; Cong, R.; Fahad, S.; Li, X. Effects of fertilization on crop production and nutrient-supplying capacity under rice-oilseed rape rotation system. Sci. Rep. 2017, 7, 1270. [Google Scholar] [CrossRef]
- Bhiah, K.; Guppy, C.; Lockwood, P.; Jessop, R. Effect of potassium on rice lodging under high nitrogen nutrition. In Proceedings of the 19th World Congress of Soil Science Soil Solution for a Changing World, Brisbane, Australia, 1–6 August 2010; pp. 1–6. [Google Scholar]
- Zhu, G.; Peng, S.; Huang, J.; Cui, K.; Nie, L.; Wang, F. Genetic improvements in rice yield and concomitant increases in radiation-and nitrogen-use efficiency in middle reaches of Yangtze river. Sci. Rep. 2016, 6, 21049. [Google Scholar] [CrossRef]
- Yuan, Z.; Li, Y.; Zhang, J.; He, D. Influence of phosphate and potash fertilizers on mechanical properties of wheat stem. Trans. Chin. Soc. Agric. Eng. 2010, 26, 147–150. [Google Scholar]
- White, P.; Karley, A.; Potassium, R.; Mendel, R. Cell Biology of Metals and Nutrients; Springer: Berlin/Heidelberg, Germany, 2010; pp. 199–224. [Google Scholar]
- Hafsi, C.; Debez, A.; Abdelly, C. Potassium deficiency in plants: Effects and signaling cascades. Acta Physiol. Plant. 2014, 36, 1055–1070. [Google Scholar] [CrossRef]
- Waseem, M.; Ahmad, R.; Randhawa, M.A.; Aziz, T.; Mahmood, N. Influence Of Silicon Application On Blast Incidence And Lodging Resistance In Rice. J. Agric. Res. 2016, 54, 435–443. [Google Scholar]
- Fallah, A. Studies effect of silicon on lodging parameters in rice plant under hydroponics culture in a greenhouse experiment. In Proceedings of the Silicon in Agriculture Conference, Wild Coast Sun, South Africa, 26–31 October 2008; pp. 26–31. [Google Scholar]
- Salman, D.; Morteza, S.; Dariush, Z.; Abbas, G.M.; Reza, Y.; Ehsan, G.D.; Reza, N.A. Application of nitrogen and silicon rates on morphological and chemical lodging related characteristics in rice (Oryza sativa L.) at North of Iran. J. Agric. Sci. 2012, 4, 12. [Google Scholar] [CrossRef]
- Fallah, A. Silicon effect on lodging parameters of rice plants under hydroponic culture. Int. J. Agric. Sci. 2012, 2, 630–634. [Google Scholar]
- Liang, Y.C.; Ma, T.S.; Li, F.J.; Feng, Y.J. Silicon availability and response of rice and wheat to silicon in calcareous soils. Commun. Soil Sci. Plant Anal. 1994, 25, 2285–2297. [Google Scholar] [CrossRef]
- Uchimura, Y.; Ogata, T.; Sato, H.; Matsue, Y. Effects of silicate application on lodging, yield and palatability of rice [Oryza sativa] grown by direct sowing culture. Jpn. J. Crop Sci. 2000, 69, 487–492. [Google Scholar] [CrossRef]
- Kim, S.G.; Kim, K.W.; Park, E.W.; Choi, D. Silicon-induced cell wall fortification of rice leaves: A possible cellular mechanism of enhanced host resistance to blast. Phytopathology 2002, 92, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- San, N.S.; Ootsuki, Y.; Adachi, S.; Yamamoto, T.; Ueda, T.; Tanabata, T.; Motobayashi, T.; Ookawa, T.; Hirasawa, T. A near-isogenic rice line carrying a QTL for larger leaf inclination angle yields heavier biomass and grain. Field Crop. Res. 2018, 219, 131–138. [Google Scholar] [CrossRef]
- Ma, J.; Ma, W.; Tian, Y.; Yang, J.; Zhou, K.; Zhu, Q. The culm lodging resistance of heavy panicle type of rice. Zuo Wu Xue Bao 2004, 30, 143–148. [Google Scholar]
- Rehman, B. Silicon elicited varied physiological and biochemical responses in Indian mustard (Brassica juncea): A concentration dependent study. Isr. J. Plant Sci. 2016, 63, 158–166. [Google Scholar] [CrossRef]
- Tanaka, A.; Park, Y. Significance of the absorption and distribution of silica in the growth of the rice plant. Soil Sci. Plant Nutr. 1966, 12, 23–28. [Google Scholar] [CrossRef]
- Hernandez-Apaolaza, L. Can silicon partially alleviate micronutrient deficiency in plants? A review. Planta 2014, 240, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Dorairaj, D.; Ismail, M.R.; Sinniah, U.R.; Kar Ban, T. Influence of silicon on growth, yield, and lodging resistance of MR219, a lowland rice of Malaysia. J. Plant Nutr. 2017, 40, 1111–1124. [Google Scholar] [CrossRef]
- Tripathi, S.; Sayre, K.; Kaul, J.; Narang, R. Lodging behavior and yield potential of spring wheat (Triticum aestivum L.): Effects of ethephon and genotypes. Field Crop. Res. 2004, 87, 207–220. [Google Scholar] [CrossRef]
- Sayre, K.D.; Rajaram, S.; Fischer, R. Yield potential progress in short bread wheats in northwest Mexico. Crop Sci. 1997, 37, 36–42. [Google Scholar] [CrossRef]
- Yao, J.; Ma, H.; Zhang, P.; Ren, L.; Yang, X.; Yao, G.; Zhang, P.; Zhou, M. Inheritance of stem strength and its correlations with culm morphological traits in wheat (Triticum aestivum L.). Can. J. Plant Sci. 2011, 91, 1065–1070. [Google Scholar] [CrossRef]
- Choudhary, B.; Gheysen, G.; Buysse, J.; van der Meer, P.; Burssens, S. Regulatory options for genetically modified crops in India. Plant Biotechnol. J. 2014, 12, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Edwards, M.A. Morphological features of wheat grain and genotype affecting flour yield. Ph.D. Thesis, Southern Cross University, Lismore, NSW, Australia, 2010. [Google Scholar]
- Matsumoto, T. Norin 10. a Dwarf Winter Wheat Variety; Agriculture Forestry and Fisheries Research Council: Tokoy, Japan, 1968. [Google Scholar]
- Chandler, R.F., Jr. The impact of improved tropical plant type on rice yields in South and Southeast Asia. Rice Breed. 1972, 77–85. [Google Scholar]
- IARI. IARI Perspective Plan-Vision 2025; Indian Agricultural Research Institute: New Delhi, India, 2007. [Google Scholar]
- Hayashi, S.; Kamoshita, A.; Yamagishi, J. Effect of planting density on grain yield and water productivity of rice (Oryza sativa L.) grown in flooded and non-flooded fields in Japan. Plant Prod. Sci. 2006, 9, 298–311. [Google Scholar] [CrossRef]
- Luo, Y.; Yin, Z. Marker-assisted breeding of Thai fragrance rice for semi-dwarf phenotype, submergence tolerance and disease resistance to rice blast and bacterial blight. Mol. Breed. 2013, 32, 709–721. [Google Scholar] [CrossRef]
- Xu, C.; Gao, Y.; Tian, B.; Ren, J.; Meng, Q.; Wang, P. Effects of EDAH, a novel plant growth regulator, on mechanical strength, stalk vascular bundles and grain yield of summer maize at high densities. Field Crop. Res. 2017, 200, 71–79. [Google Scholar] [CrossRef]
- Rajala, A. Plant Growth Regulators to Manipulate Cereal Growth in Northern Growing Conditions. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2003. [Google Scholar]
- Böse, G.; Graebe, J.E.; Grosselindemann, E.; Hedden, P.; Aach, H.; Schweimer, A.; Sydow, S.; Lange, T. The biosynthesis of ent-kaurene in germinating seeds and the function of 2-oxoglutarate in gibberellin biosynthesis. In Progress in Plant Growth Regulation; Springer: Berlin/Heidelberg, Germany, 1992; pp. 545–554. [Google Scholar]
- Hedden, P.; Phillips, A.L. Gibberellin metabolism: New insights revealed by the genes. Trends Plant Sci. 2000, 5, 523–530. [Google Scholar] [CrossRef]
- Rademacher, W. Growth retardants: Effects on gibberellin biosynthesis and other metabolic pathways. Annu. Rev. Plant Biol. 2000, 51, 501–531. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Cerezo, S.; Martínez-Montiel, N.; García-Sánchez, J.; Pérez-y-Terrón, R.; Martínez-Contreras, R.D. Gibberellin biosynthesis and metabolism: A convergent route for plants, fungi and bacteria. Microbiol. Res. 2018, 208, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaruni, S.V.; Maddila, S.; Gangu, K.K.; Jonnalagadda, S.B. A Review on multi-component green synthesis of N-containing heterocycles using mixed oxides as heterogeneous catalysts. Arab. J. Chem. 2017. [Google Scholar] [CrossRef]
- Bömke, C.; Tudzynski, B. Diversity, regulation, and evolution of the gibberellin biosynthetic pathway in fungi compared to plants and bacteria. Phytochemistry 2009, 70, 1876–1893. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P.; Sponsel, V. A century of gibberellin research. J. Plant Growth Regul. 2015, 34, 740–760. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.; Fedak, G. Effects of chlormequat on plant height, disease development and chemical constituents of cultivars of barley, oats, and wheat. Can. J. Plant Sci. 1977, 57, 31–36. [Google Scholar] [CrossRef]
- Lowe, L.; Carter, O. Response of four wheat cultivars to applications of (2-chloroethyl) trimethylammonium chloride (CCC). Aust. J. Exp. Agric. 1972, 12, 75–80. [Google Scholar] [CrossRef]
- Mathews, P.; Caldicott, J. The effect of chlormequat chloride formulated with choline chloride on the height and yield of winter wheat. Ann. Appl. Biol. 1981, 97, 227–236. [Google Scholar] [CrossRef]
- Caldwell, C.; Starratt, C. Response of Max spring wheat to management inputs. Can. J. Plant Sci. 1987, 67, 645–652. [Google Scholar] [CrossRef]
- Leśniowska-Nowak, J.; Nowak, M.; Zapalska, M.; Dudziak, K.; Kowalczyk, K. Influence of CCC and trinexapac-ethyl on the expression of genes involved in gibberellic biosynthesis and metabolism pathway in isogenic line with Rht12 dwarfing gene. Acta Sci. Polonorum. Hortorum Cultus 2017, 16. [Google Scholar] [CrossRef]
- Shekoofa, A.; Emam, Y. Effects of nitrogen fertilization and plant growth regulators (PGRs) on yield of wheat (Triticum aestivum L.) cv. Shiraz. J. Agric. Sci. Technol. 2008, 10, 101–108. [Google Scholar]
- Khan, A.; Spilde, L. Agronomic and economic response of spring wheat cultivars to ethephon. Agron. J. 1992, 84, 399–402. [Google Scholar] [CrossRef]
- Spitzer, T.; Matušinský, P.; Klemová, Z.; Kazda, J. Management of sunflower stand height using growth regulators. Plant Soil Environ. 2011, 57, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Kamran, M.; Ahmad, I.; Wu, X.; Liu, T.; Ding, R.; Han, Q. Application of paclobutrazol: A strategy for inducing lodging resistance of wheat through mediation of plant height, stem physical strength, and lignin biosynthesis. Environ. Sci. Pollut. Res. 2018, 25, 29366–29378. [Google Scholar] [CrossRef]
- Zhang, W.; Peng, C.; Sun, G.; Zhang, F.; Hu, S. Effect of different external phytohormones on leaves senescence in late growth period of late-season rice. Acta Agric Jiangxi 2007, 19, 11–13. [Google Scholar]
- Espindula, M.; Rocha, V.; Grossi, J.; Souza, M.; Souza, L.; Favarato, L. Use of growth retardants in wheat. Planta Daninha 2009, 27, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Koch, F.; Aisenberg, G.; Monteiro, M.; Pedó, T.; Zimmer, P.; Villela, F.; Aumonde, T. Growth of wheat plants submitted to the application of the growth regulator trinexapac-ethyl and vigor of the produced seeds. Agrociencia 2017, 21, 24–32. [Google Scholar]
- Zagonel, J.; Fernandes, E. Rates and application times of growth reducer affecting wheat cultivars at two nitrogen rates. Planta Daninha 2007, 25, 331–339. [Google Scholar] [CrossRef]
- Zagonel, J.; Venancio, W.; Kunz, R. Effect of growth regulator on wheat crop under different nitrogen rates and plant densities. Planta Daninha 2002, 20, 471–476. [Google Scholar] [CrossRef]
- Merry, A.M.; Dean, G.; Botwright Acuna, T. What are the effects of chlormequat and trinexapac-ethyl alone or in combination on lodging, height and yield of winter wheat in Tasmania? In Proceedings of the 17th Australian Society of Agronomy Conference, Hobart, Australia, 20–24 September 2015; pp. 1–4. [Google Scholar]
- Taiz, L.; Zeiger, E. Gibberellins: Regulators of Plant Height and Seed Germination. Plant Physiol. Sinauer Assoc. Sunderland 2010, 583–619. [Google Scholar]
- Plaza-Wüthrich, S.; Blösch, R.; Rindisbacher, A.; Cannarozzi, G.; Tadele, Z. Gibberellin deficiency confers both lodging and drought tolerance in small cereals. Front. Plant Sci. 2016, 7, 643. [Google Scholar] [CrossRef] [PubMed]
- French, P.; Matsuyuki, H.; Ueno, H. Paclobutrazol: Control of lodging in Japanese paddy rice. In Pest Management in Rice; Springer: Berlin/Heidelberg, Germany, 1990; pp. 474–485. [Google Scholar]
- Na, C.-I.; Hamayun, M.; Khan, A.L.; Kim, Y.-H.; Choi, K.-I.; Kang, S.-M.; Kim, S.-I.; Kim, J.-T.; Won, J.-G.; Lee, I.-J. Influence of prohexadione-calcium, trinexapac-ethyl and hexaconazole on lodging characteristic and gibberellin biosynthesis of rice (Oryza sativa L.). Afr. J. Biotechnol. 2011, 10, 13097–13106. [Google Scholar]
- Kim, H.; Lee, I.; Hamayun, M.; Kim, J.; Won, J.; Hwang, I.; Kim, K. Effect of prohexadione calcium on growth components and endogenous gibberellins contents of rice (Oryza sativa L.). J. Agron. Crop Sci. 2007, 193, 445–451. [Google Scholar] [CrossRef]
- Pan, S.; Rasul, F.; Li, W.; Tian, H.; Mo, Z.; Duan, M.; Tang, X. Roles of plant growth regulators on yield, grain qualities and antioxidant enzyme activities in super hybrid rice (Oryza sativa L.). Rice 2013, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Zhang, J.; Li, J.; Zhao, Q. Effects of exogenous hormones on grain plumpness and quality of rice. J. Henan Agric. Sci. 2010, 12, 22–25. [Google Scholar]
- Hashem, I.; Naeem, E.; Metwally, T.; El Sharkawi, H. Enhancement of lodging resistance and productivity of rice using growth regulators at different nitrogen levels. J. Plant Breed. Crop Sci. 2016, 8, 34–44. [Google Scholar]
- Bridgemohan, P.; Bridgemohan, R.S. Evaluation of anti-lodging plant growth regulators on the growth and development of rice (Oryza sativa). J. Cereals Oilseeds 2014, 5, 12–16. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait (s) | Crop | Behavior | Reference (s) |
|---|---|---|---|
| 1. Morphological Features | |||
| Plant height | Wheat and rice | Positively correlated with lodging | [27,29,30,37,39] |
| Primary inter-nodal length | Wheat and rice | Negatively correlated with lodging resistance | [45] |
| Culm diameter | Wheat and rice | Strongly positively association with resistance to lodging | [30,36,39,47] |
| Culm length | Wheat and rice | Positively correlated with lodging | [59] |
| Culm diameter, panicle weight, panicle length | Rice | Positively correlated with resistance to lodging | [59] |
| 2. Anatomical Aspects | |||
| Number of vascular bundles | Rice | Strongly positively associated with resistance to lodging | [63] |
| Width of mechanical tissue layer | Wheat | Strongly positively associated with resistance to lodging | [37] |
| 3. Biochemical Features | |||
| Lignin and cellulose contents | Wheat and rice | Strongly positively correlated with culm strength, secondary cell wall strength, and lodging resistance | [37] |
| 4-Plant Root System | |||
| Root lodging resistance | Wheat and rice | Associated to diameter of the root soil cone, bending strength of the crown, and the shear strength of the soil | [70,72,79] |
| Chromosomes/(QTLS) | Flanking Marker Loci | Impact on Plant Traits | Reference (s) |
|---|---|---|---|
| Wheat | |||
| 1BS, 4AS, 7BL | Xpsr949-Xgwm18, Xgwm397-Xglk315, Xpsr927-Xpsr350 | Shorter plant height | [131] |
| 2AS, 3AS, 5AL | Xpsr958-Xpsr566c, Xpsr598-Xpsr570, Xpsr918b-Xpsr1201a | Culm wall thickness | [131] |
| 4B, 4D | Xgmti538-Xgwm6, XgKm60S-Xgdml29 | Shorter plant height | [132] |
| 3A(QSs-3A), 3B (QSs-3B) | Xwmc527-Xwmc21, Xgwm108-Xwmc291 | Stem strength | [133] |
| QSd-3B | Xgwm108-Xwmc291 | Culm diameter | [133] |
| 1A(QPd-1A),2D (QPd-2D) | Xgwm135-Xwmc84, Xgwm311-Xgwm301 | Pith diameter | [133] |
| QCwt-2D | Xgwm311-Xwmc301 | Culm wall thickness | [133] |
| 3BL | GWM247 and GWM340 | Lignin contents | [37] |
| 1D, 2A, 3A, 4A, 6A, 7D | Xgwm642, Xgdm93, Xgdm93, Xwmc313, Xgwm570, wPt-9690, Xbarc184, respectively | Control Plant height | [30] |
| 3A, 4D | Xwmc264, Xwmc48 8, respectively | Meaningful effect on lodging risk | [30] |
| 3A | Xgwm369 | Association with lignin and cellulose | [30] |
| 1A, 6A | Xcfa2153, Xwmc32 | Association with gravity of the stem | [30] |
| 3BS | Xbarc102 | Reduce plant height | [134] |
| 6A | Xwmc256-Xbarc103 | Reduce plant height | [137] |
| Rice | |||
| Chr. 1 (SCM1) | RM8111-RM8067 | Culm strength | [52,128] |
| Chr. 6 (SCM2) | RM6395 and RM5509 | Culm strength, increased spikelet number, and grain yield | [52,128,129,138] |
| Chr. 3 (SCM3) | RM15761-RM15782 | Culm strength and spikelet number | [52,128,139,140] |
| Chr. 2 (SCM4) | RM3703-RM2468 | Culm lodging resistance | [52,128,140] |
| Chr. 4, 5 (prl5) 6,11(prl11), 12 | C946, C1081, R2171, C82, G1406 | Providing resistance to the basal portion of the stalk | [140] |
| Chr.1, 3, 6, 7, 8, 12 | C885, C595, C358, C451, C10122, R3375 | Culm diameter | [140] |
| Chr. 5, 5, 6 | R1838, C246, R2549 | Lodging resistance from typhoon | [140] |
| 1(qCL1.1); 2(qCL2.1); 7(qCL7.1) | id1021344, id2004861, id7002801 | Culm length | [59] |
| 1(qCD1.1); 2(qCD2.1); 7(qCD7.1) | id1003559, id2007818, id7001246 | Culm diameter | [59] |
| 1(qCS1.1); 2(qCS2.1); 2(qCS2.2); 2(qCS2.3); 6(qCS6.1); 6(qCS6.2) | id1003559, id2007818, id2006621, id2008112, id6001960, id6010515 | Culm strength | [59] |
| Gene (s) | Crop | Impact on Plant Traits | Reference (s) |
|---|---|---|---|
| Rht1 and Rht2 | Wheat | Reduced internode length, Reduce plant height (Wheat) | [145,146,147,148,149] |
| Rht3 | Wheat | Reduce plant height (Wheat) | [154] |
| Rht5 and Ppd-D1 | Wheat | Reduce plant height | [28,155,156,157] |
| Rht7 and Rh8 | Wheat | Reduce plant height | [158,159] |
| Rht-B1, Rht-B1a, Rht-B1a, Rht-D1, Rht8, Ppd-B1 and Ppd-D1 | Wheat | Reduce plant height | [160] |
| Rht4, Rht5, Rht8, Rht9, Rht12 and Rht13 | Wheat | Shortening plant height | [134] |
| Rht8 and Rht-B1b or Rht-D1b | Wheat | Increased roots and reduced plant height | [163,164] |
| Rht14, Rht15, Rht16, Rht18 and Rht19 | Wheat | Reduced plant height | [166,167] |
| Rht22, Rht-B1IC12196 and Rht-B1f | Wheat | Dwarfing genes | [166,167,169] |
| Rht24 | Wheat | Dwarfing genes | [137] |
| COMT (W-cm5-1) | Wheat | Expressed in root, plant stalk, and leaf tissues | [170] |
| semidwarf1 (sd1) | Rice | Reduce plant height | [19] |
| OsCesA1, OsCesA3 and OsCesA8 | Rice | Cellulose synthases | [174] |
| OsCesA4, OsCesA9, and OsCesA7 | Rice | Expressed in secondary cell walls | [175] |
| OsCLF, OsEMF2b and OsFIE2 | Rice | Dwarf phenotype | [176] |
| OsEMF2b | Rice | Dwarf phenotype | [178] |
| TaCM, | Wheat | Relationship with the accumulation of stem lignin and stalk lodging mechanical strength | [95] |
| PAL, CCR, CCoAOMT, COMT and CAD | Wheat | Lignin biosynthesis enzymes | [180] |
| Genotype (s) | Lodging Rating | Reference (s) |
|---|---|---|
| Wheat | ||
| PBW 343, UP 2338 | Tolerant | [249] |
| Baviacora 92, Seri 82, Star, Munia/Kauz, Weaver | Tolerant | [249] |
| Baviacora 92 | Moderately resistant | [250] |
| Zhoumai 18, Zhoumai 22 and Pingan 6, Aikang 58 | Resistant | [251] |
| Kalyansona and Sonalika | Resistant | [252] |
| Savannah, Rialto, Buster, Hereward | Resistant | [47] |
| Norin 10 | Resistant | [253,254] |
| Hereward, Spark, Cadenza, Mercia | Resistant | [222] |
| H4564 and C6001 | Resistant | [170] |
| Oslo, Sapphire, AC Foremost, ND695 and AC Vista and Kohika | Tolerant | [27] |
| Rice | ||
| Wuyunjing 23 | Resistant | [38,46] |
| IR-8 | Resistant | [255] |
| Pusa Basmati-1, Pusa-1121, RH-10 | Resistant | [256] |
| Dontokoi, IR24 | Resistant | [257] |
| T5105 | Resistant | [258] |
| Yliangyou2 | Resistant | [15] |
| Takanari | Resistant | [129] |
| Peiai64, Zhefu802, Liantangzhao, 76-27B, Chunjiang025, Xiushui04, Xiushui63, Jia02-43, ZH222, Bing02-133, Taihunuo, HanfenB, Kinmaze, HZ0302, Jia02-5, Minghui63, Teqing, 9308, Guangsi, K17B, QingreB, 486B, 5N-76B, Bing02-09, DiguB, V20B, Aijao-Nante, Zhaiyeqing8,GuangB, Guichao, lemont, M202, M201, 98–110, Bing02-105, R0308, Shirasenbon, Fukuhibiki | High-yielding semi-dwarf rice cultivars contain Sd1 gene | [171] |
| Crop | Application Time | Height Reduction | Grain Yield | Reference (s) |
|---|---|---|---|---|
| Effect of Chlormequat | ||||
| Wheat | leaf growth stages (from stage 3 to 5) | 3.3–14.5% | No effect | [268] |
| Spring wheat | Second node detectable/third node visible | 1–29% | No effect | [269] |
| Winter wheat | Pseudo stem erection-first node visible | 6% | No effect | [270] |
| Winter wheat | Pseudo stem erection/first node visible | 3–15% | No effect | [7] |
| Spring wheat | Vegetative development | 20 cm | Smaller effect | [271] |
| Effect of Ethephon | ||||
| Spring wheat | 12 cm | No effect | [271] | |
| Spring wheat | Booting stage | 1–12% | Increase grain yield (5.4%) | [274] |
| Wheat | Before anthesis | 9.0% | Reduced grain yield (8.3%) | [249] |
| Effect of Trinexapac-ethyl | ||||
| Wheat | Stem elongation | Reduced plant height | Reduced wheat yield | [279,280] |
| Paclobutrazol | ||||
| Wheat | Stem elongation stage | 10–20% | Increased grain yield | [5] |
| Rice | First growing seedlings | 20–30% | Increased grain yield | [284] |
| Rice | 15–10 days before heading | 15–25% | Increased yield (15%) | [285] |
| Prohexadione-calcium (Pro-Ca) and trinexapac-ethyl (TNE) Effect | ||||
| Rice | 5–10 days before heading | 5–10% | No effects | [287] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, L.; Yahya, M.; Shah, S.M.A.; Nadeem, M.; Ali, A.; Ali, A.; Wang, J.; Riaz, M.W.; Rehman, S.; Wu, W.; et al. Improving Lodging Resistance: Using Wheat and Rice as Classical Examples. Int. J. Mol. Sci. 2019, 20, 4211. https://doi.org/10.3390/ijms20174211
Shah L, Yahya M, Shah SMA, Nadeem M, Ali A, Ali A, Wang J, Riaz MW, Rehman S, Wu W, et al. Improving Lodging Resistance: Using Wheat and Rice as Classical Examples. International Journal of Molecular Sciences. 2019; 20(17):4211. https://doi.org/10.3390/ijms20174211
Chicago/Turabian StyleShah, Liaqat, Muhammad Yahya, Syed Mehar Ali Shah, Muhammad Nadeem, Ahmad Ali, Asif Ali, Jing Wang, Muhammad Waheed Riaz, Shamsur Rehman, Weixun Wu, and et al. 2019. "Improving Lodging Resistance: Using Wheat and Rice as Classical Examples" International Journal of Molecular Sciences 20, no. 17: 4211. https://doi.org/10.3390/ijms20174211
APA StyleShah, L., Yahya, M., Shah, S. M. A., Nadeem, M., Ali, A., Ali, A., Wang, J., Riaz, M. W., Rehman, S., Wu, W., Khan, R. M., Abbas, A., Riaz, A., Anis, G. B., Si, H., Jiang, H., & Ma, C. (2019). Improving Lodging Resistance: Using Wheat and Rice as Classical Examples. International Journal of Molecular Sciences, 20(17), 4211. https://doi.org/10.3390/ijms20174211