Progress in Research on the Role of Flavonoids in Lung Cancer

 ,
,  ,
, 
 ,
, 
Abstract
1. Introduction
2. Anticancer Effects of Flavonoids in Lung Cancer
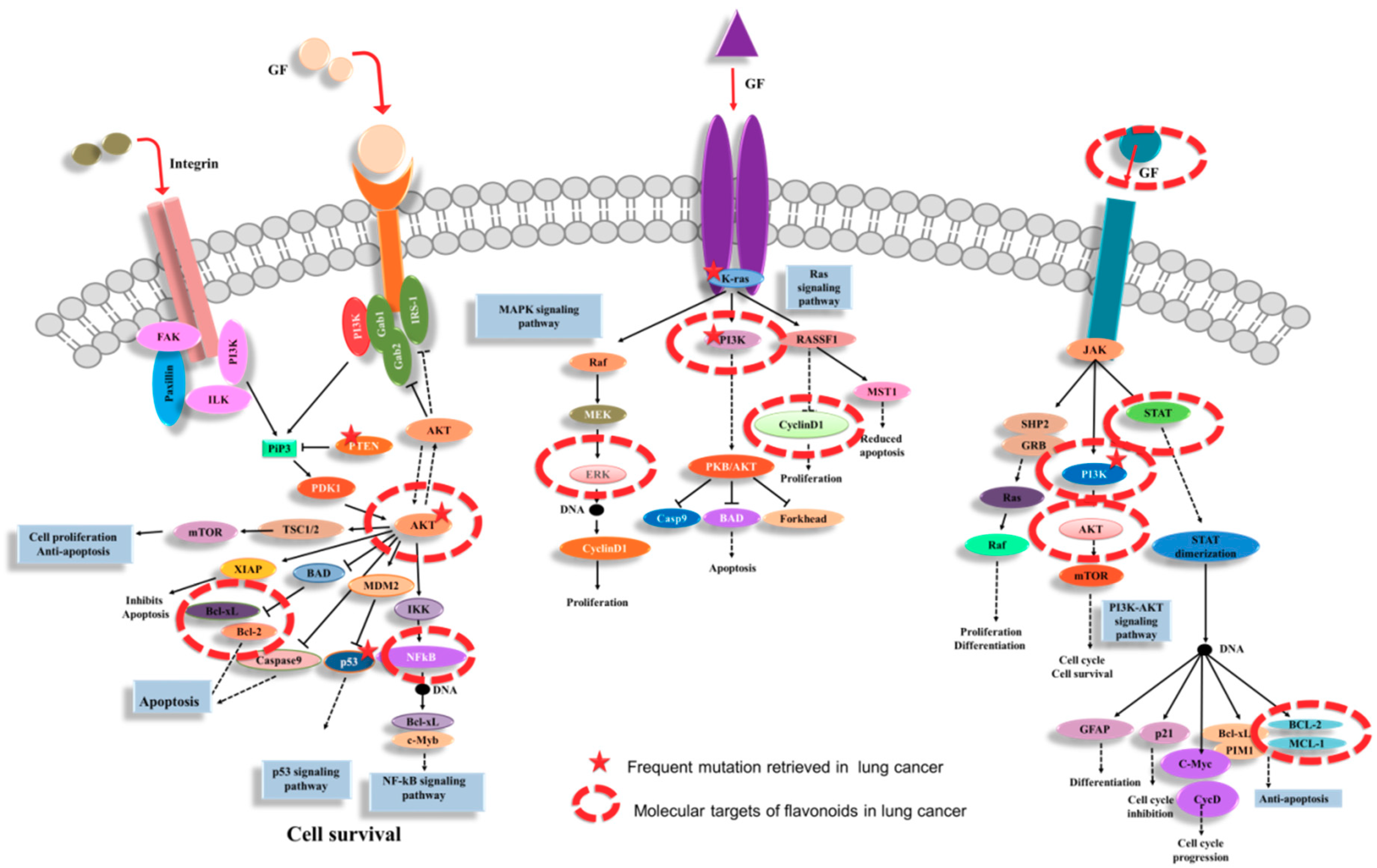
3. Flavonoids as Key Gene Expression Regulators in Lung Cancer
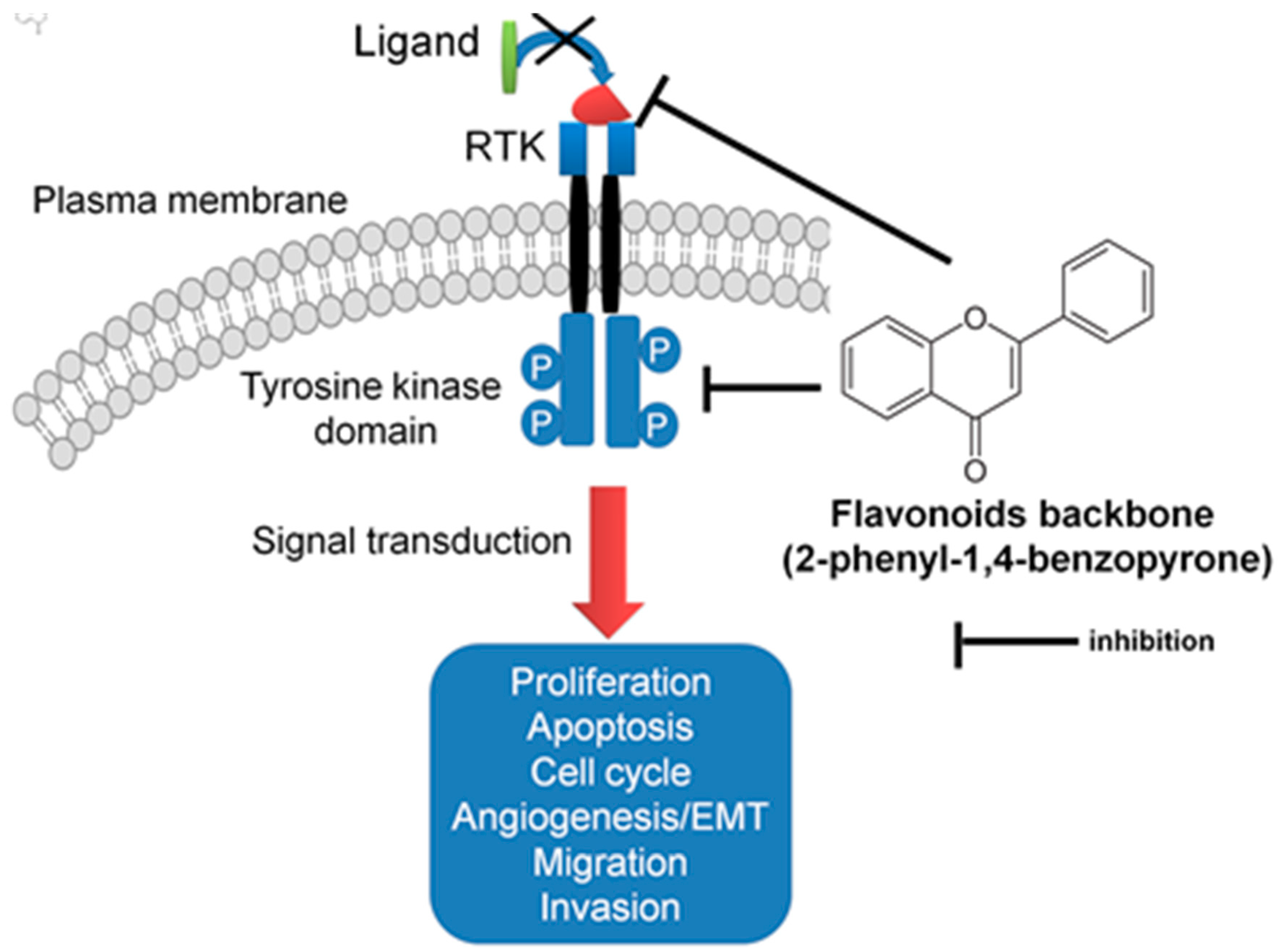
3.1. Flavonoids Interfere with Receptor Tyrosine Kinases Cascade in Lung Cancer
3.2. Flavomoids Affects Cell Proliferation, Apoptosis and Autophagy
3.3. Flavonoids as Cell Cycle Modulators
3.4. Flavonoids Regulate Invasion and Metastasis
4. Flavonoids as miRNA Modulators in Lung Cancer
5. Flavonoids in Combination with Chemotherapeutic and Radiotherapy Treatment in Lung Cancer
6. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Allemani, C.; Weir, H.K.; Carreira, H.; Harewood, R.; Spika, D.; Wang, X.S.; Bannon, F.; Ahn, J.V.; Johnson, C.J.; Bonaventure, A.; et al. Global surveillance of cancer survival 1995–2009: Analysis of individual data for 25,676,887 patients from 279 population-based registries in 67 countries (concord-2). Lancet 2015, 385, 977–1010. [Google Scholar] [CrossRef]
- Chae, D.K.; Ban, E.; Yoo, Y.S.; Kim, E.E.; Baik, J.H.; Song, E.J. Mir-27a regulates the tgf-beta signaling pathway by targeting smad2 and smad4 in lung cancer. Mol. Carcinog. 2017, 56, 1992–1998. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Altorki, N.K.; Markowitz, G.J.; Gao, D.; Port, J.L.; Saxena, A.; Stiles, B.; McGraw, T.; Mittal, V. The lung microenvironment: An important regulator of tumour growth and metastasis. Nat. Rev. Cancer 2019, 19, 9–31. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Tateishi, A.; Bychkov, A.; Fukuoka, J. Remarkable alteration of pd-l1 expression after immune checkpoint therapy in patients with non-small-cell lung cancer: Two autopsy case reports. Int. J. Mol. Sci. 2019, 20, 2578. [Google Scholar] [CrossRef] [PubMed]
- Vigneswaran, J.; Tan, Y.H.; Murgu, S.D.; Won, B.M.; Patton, K.A.; Villaflor, V.M.; Hoffman, P.C.; Hensing, T.; Hogarth, D.K.; Malik, R.; et al. Comprehensive genetic testing identifies targetable genomic alterations in most patients with non-small cell lung cancer, specifically adenocarcinoma, single institute investigation. Oncotarget 2016, 7, 18876–18886. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; More, S.; Murtuza, A.; Woodward, B.D.; Husain, H. New targets in non–small cell lung cancer. Hematol. Oncol. Clin. N. Am. 2017, 31, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Snyder-Talkington, B.N.; Dong, C.; Singh, S.; Raese, R.; Qian, Y.; Porter, D.W.; Wolfarth, M.G.; Guo, N.L. Multi-walled carbon nanotube-induced gene expression biomarkers for medical and occupational surveillance. Int. J. Mol. Sci. 2019, 20, 2635. [Google Scholar] [CrossRef] [PubMed]
- Shivappa, N.; Wang, R.; Hebert, J.R.; Jin, A.; Koh, W.P.; Yuan, J.M. Association between inflammatory potential of diet and risk of lung cancer among smokers in a prospective study in singapore. Eur. J. Nutr. 2018. [Google Scholar] [CrossRef] [PubMed]
- Krusinska, B.; Hawrysz, I.; Wadolowska, L.; Slowinska, M.A.; Biernacki, M.; Czerwinska, A.; Golota, J.J. Associations of mediterranean diet and a posteriori derived dietary patterns with breast and lung cancer risk: A case-control study. Nutrients 2018, 10, 470. [Google Scholar] [CrossRef] [PubMed]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P. Polyphenols and human health: Prevention of disease and mechanisms of action. Nutrients 2010, 2, 1106–1131. [Google Scholar] [CrossRef] [PubMed]
- Budisan, L.; Gulei, D.; Zanoaga, O.M.; Irimie, A.I.; Sergiu, C.; Braicu, C.; Gherman, C.D.; Berindan-Neagoe, I. Dietary intervention by phytochemicals and their role in modulating coding and non-coding genes in cancer. Int. J. Mol. Sci. 2017, 18, 1178. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.; Yen, H.; Hsiao, H.Y.; Su, S.C. Phytochemicals in skin cancer prevention and treatment: An updated review. Int. J. Mol. Sci. 2018, 19, 941. [Google Scholar] [CrossRef] [PubMed]
- Cojocneanu Petric, R.; Braicu, C.; Raduly, L.; Zanoaga, O.; Dragos, N.; Monroig, P.; Dumitrascu, D.; Berindan-Neagoe, I. Phytochemicals modulate carcinogenic signaling pathways in breast and hormone-related cancers. Oncotargets Ther. 2015, 8, 2053–2066. [Google Scholar] [CrossRef] [PubMed]
- Chahar, M.K.; Sharma, N.; Dobhal, M.P.; Joshi, Y.C. Flavonoids: A versatile source of anticancer drugs. Pharmacogn. Rev. 2011, 5, 1–12. [Google Scholar] [PubMed]
- Braicu, C.; Pilecki, V.; Balacescu, O.; Irimie, A.; Neagoe, I.B. The relationships between biological activities and structure of flavan-3-ols. Int. J. Mol. Sci. 2011, 12, 9342–9353. [Google Scholar] [CrossRef] [PubMed]
- Batra, P.; Sharma, A.K. Anti-cancer potential of flavonoids: Recent trends and future perspectives. 3 Biotech 2013, 3, 439–459. [Google Scholar] [CrossRef] [PubMed]
- Budisan, L.; Gulei, D.; Jurj, A.; Braicu, C.; Zanoaga, O.; Cojocneanu, R.; Pop, L.; Raduly, L.; Barbat, A.; Moldovan, A.; et al. Inhibitory effect of cape and kaempferol in colon cancer cell lines-possible implications in new therapeutic strategies. Int. J. Mol. Sci. 2019, 20, 1199. [Google Scholar] [CrossRef]
- Qiu, T.; Wu, D.; Yang, L.; Ye, H.; Wang, Q.; Cao, Z.; Tang, K. Exploring the mechanism of flavonoids through systematic bioinformatics analysis. Front. Pharmacol. 2018, 9, 918. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, T.; Chen, G.Y. Flavonoids and colorectal cancer prevention. Antioxidant 2018, 7, 187. [Google Scholar] [CrossRef]
- Shin, S.Y.; Lee, Y.; Kim, B.S.; Lee, J.; Ahn, S.; Koh, D.; Lim, Y.; Lee, Y.H. Inhibitory effect of synthetic flavone derivatives on pan-aurora kinases: Induction of g2/m cell-cycle arrest and apoptosis in hct116 human colon cancer cells. Int. J. Mol. Sci. 2018, 19, 4086. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, R.; Datt, M.; Liu, X.; Gupta, S. Dietary flavones as dual inhibitors of DNA methyltransferases and histone methyltransferases. PLoS ONE 2016, 11, e0162956. [Google Scholar]
- Xingyu, Z.; Peijie, M.; Dan, P.; Youg, W.; Daojun, W.; Xinzheng, C.; Xijun, Z.; Yangrong, S. Quercetin suppresses lung cancer growth by targeting aurora b kinase. Cancer Med. 2016, 5, 3156–3165. [Google Scholar] [CrossRef] [PubMed]
- Sonoki, H.; Tanimae, A.; Endo, S.; Matsunaga, T.; Furuta, T.; Ichihara, K.; Ikari, A. Kaempherol and luteolin decrease claudin-2 expression mediated by inhibition of stat3 in lung adenocarcinoma a549 cells. Nutrients 2017, 9, 597. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, G.; Zhang, X.; Zhang, Q.; Zheng, S.; Wang, G.; Chen, Q.-H. A new class of flavonol-based anti-prostate cancer agents: Design, synthesis, and evaluation in cell models. Bioorganic Med. Chem. Lett. 2016, 26, 4241–4245. [Google Scholar] [CrossRef] [PubMed]
- Chanet, A.; Milenkovic, D.; Manach, C.; Mazur, A.; Morand, C. Citrus flavanones: What is their role in cardiovascular protection? J. Agric. Food Chem. 2012, 60, 8809–8822. [Google Scholar] [CrossRef] [PubMed]
- Woo, Y.; Shin, S.Y.; Hyun, J.; Lee, S.D.; Lee, Y.H.; Lim, Y. Flavanones inhibit the clonogenicity of hct116 cololectal cancer cells. Int. J. Mol. Med. 2012, 29, 403–408. [Google Scholar] [PubMed]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in cancer and apoptosis. Cancers 2018, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.-J.; Hwang, K.-A.; Choi, K.-C. Antitumor effect of various phytochemicals on diverse types of thyroid cancers. Nutrients 2019, 11, 125. [Google Scholar] [CrossRef] [PubMed]
- De Sousa Moraes, L.F.; Sun, X.; Peluzio, M.; Zhu, M.J. Anthocyanins/anthocyanidins and colorectal cancer: What is behind the scenes? Crit. Rev. Food Sci. Nutr. 2019, 59, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Tang, M.; Liu, Y.; Zhang, Z.; Lu, R.; Lu, J. Apigenin inhibits cell proliferation, migration, and invasion by targeting akt in the a549 human lung cancer cell line. Anti-Cancer Drugs 2017, 28, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.H.; Cheng, C.W.; Yang, Y.C.; Chen, W.S.; Hung, W.Y.; Chow, J.M.; Chen, P.S.; Hsiao, M.; Lee, W.J.; Chien, M.H. Downregulating cd26/dppiv by apigenin modulates the interplay between akt and snail/slug signaling to restrain metastasis of lung cancer with multiple egfr statuses. J. Exp. Clin. Cancer Res. 2018, 37, 199. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Ahn, K.C.; Choi, J.Y.; Hwang, S.G.; Kim, W.J.; Um, H.D.; Park, J.K. Luteolin acts as a radiosensitizer in nonsmall cell lung cancer cells by enhancing apoptotic cell death through activation of a p38/ros/caspase cascade. Int. J. Oncol. 2015, 46, 1149–1158. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liu, C.F.; Gao, N.; Zhao, J.; Xu, J. Kaempferol suppresses proliferation but increases apoptosis and autophagy by up-regulating microrna-340 in human lung cancer cells. Biomed. Pharmacother. 2018, 108, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Xia, R.; Sheng, X.; Xu, X.; Yu, C.; Lu, H. Hesperidin induces apoptosis and g0/g1 arrest in human non-small cell lung cancer a549 cells. Int. J. Mol. Med. 2018, 41, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.C.; Tsao, T.C.; Hsu, S.R.; Wang, H.C.; Tsai, T.C.; Kao, J.Y.; Way, T.D. Egcg inhibits transforming growth factor-beta-mediated epithelial-to-mesenchymal transition via the inhibition of smad2 and erk1/2 signaling pathways in nonsmall cell lung cancer cells. J. Agric. Food Chem. 2012, 60, 9863–9873. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Li, J.; Li, B.; Wang, Y.; Li, M.; Ma, D.; Wang, X. Genistein exhibits anti-cancer effects via down-regulating foxm1 in h446 small-cell lung cancer cells. Tumour Biol. 2014, 35, 4137–4145. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Mukhtar, H. Dietary agents for prevention and treatment of lung cancer. Cancer Lett. 2015, 359, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Mehterov, N.; Vladimirov, B.; Sarafian, V.; Nabavi, S.M.; Atanasov, A.G.; Berindan-Neagoe, I. Nutrigenomics in cancer: Revisiting the effects of natural compounds. Semin. Cancer Biol. 2017, 46, 84–106. [Google Scholar] [CrossRef]
- Hou, D.X.; Kumamoto, T. Flavonoids as protein kinase inhibitors for cancer chemoprevention: Direct binding and molecular modeling. Antioxid. Redox Signal. 2010, 13, 691–719. [Google Scholar] [CrossRef]
- Regad, T. Targeting rtk signaling pathways in cancer. Cancers 2015, 7, 1758–1784. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.L.; Zhao, Y.N.; Shi, Z.Z.; Cong, D.; Bai, Y.S. Lutein inhibits cell growth and activates apoptosis via the pi3k/akt/mtor signaling pathway in a549 human non-small-cell lung cancer cells. J. Environ. Pathol. Toxicol. Oncol. 2018, 37, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Lee, J.H.; Chai, E.Z.P.; Kanchi, M.M.; Kar, S.; Arfuso, F.; Dharmarajan, A.; Kumar, A.P.; Ramar, P.S.; Looi, C.Y.; et al. Cancer prevention and therapy through the modulation of transcription factors by bioactive natural compounds. Semin. Cancer Biol. 2016, 40–41, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.-H.; Chu, J.-H.; Kwan, H.-Y.; Su, T.; Yu, H.; Cheng, C.-Y.; Fu, X.-Q.; Guo, H.; Li, T.; Tse, A.K.-W.; et al. Inhibition of the stat3 signaling pathway contributes to apigenin-mediated anti-metastatic effect in melanoma. Sci. Rep. 2016, 6, 21731. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.C.; Li, C.; Gao, F.; Xu, Y.; Jiang, Z.B.; Liu, J.X.; Jin, L.Y. Epigallocatechin gallate inhibits the growth of human lung cancer by directly targeting the egfr signaling pathway. Oncol. Rep. 2014, 31, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, S. Fisetin inhibits the growth and migration in the a549 human lung cancer cell line via the erk1/2 pathway. Exp. Ther. Med. 2018, 15, 2667–2673. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Wang, Z.; Liu, X.; Wang, D. New development of inhibitors targeting the pi3k/akt/mtor pathway in personalized treatment of non-small-cell lung cancer. Anti-Cancer Drugs 2015, 26, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.L.; Chang, Y.M.; Lai, S.C.; Chen, K.M.; Wang, K.C.; Chiu, T.T.; Chang, F.H.; Hsu, L.S. Naringenin inhibits migration of lung cancer cells via the inhibition of matrix metalloproteinases-2 and -9. Exp. Ther. Med. 2017, 13, 739–744. [Google Scholar] [CrossRef]
- Chen, Z.; Miao, H.; Zhu, Z.; Zhang, H.; Huang, H. Daidzein induces apoptosis of non-small cell lung cancer cells by restoring STK 4/YAP 1 signaling. Int. J. Clin. Exp. Med. 2017, 10, 15205–15212. [Google Scholar]
- Cincin, Z.B.; Unlu, M.; Kiran, B.; Bireller, E.S.; Baran, Y.; Cakmakoglu, B. Molecular mechanisms of quercitrin-induced apoptosis in non-small cell lung cancer. Arch. Med. Res. 2014, 45, 445–454. [Google Scholar] [CrossRef]
- Irimie, A.I.; Braicu, C.; Pileczki, V.; Petrushev, B.; Soritau, O.; Campian, R.S.; Berindan-Neagoe, I. Knocking down of p53 triggers apoptosis and autophagy, concomitantly with inhibition of migration on ssc-4 oral squamous carcinoma cells. Mol. Cell. Biochem. 2016, 419, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Irimie, A.I.; Braicu, C.; Zanoaga, O.; Pileczki, V.; Gherman, C.; Berindan-Neagoe, I.; Campian, R.S. Epigallocatechin-3-gallate suppresses cell proliferation and promotes apoptosis and autophagy in oral cancer ssc-4 cells. Oncotargets Ther. 2015, 8, 461–470. [Google Scholar]
- Merkel, O.; Taylor, N.; Prutsch, N.; Staber, P.B.; Moriggl, R.; Turner, S.D.; Kenner, L. When the guardian sleeps: Reactivation of the p53 pathway in cancer. Mutat. Res. 2017, 773, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Vucic, D.; Dixit, V.M.; Wertz, I.E. Ubiquitylation in apoptosis: A post-translational modification at the edge of life and death. Nat. Rev. Mol. Cell Biol. 2011, 12, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Ji, P.; Liu, B.; Qiao, H.; Wang, X.; Zhou, L.; Deng, T.; Ba, Y. Apigenin enhances the cisplatin cytotoxic effect through p53-modulated apoptosis. Oncol. Lett. 2017, 13, 1024–1030. [Google Scholar] [CrossRef]
- Park, K.-I.; Park, H.-S.; Kim, M.-K.; Hong, G.-E.; Nagappan, A.; Lee, H.-J.; Yumnam, S.; Lee, W.-S.; Won, C.-K.; Shin, S.-C.; et al. Flavonoids identified from korean citrus aurantium l. Inhibit non-small cell lung cancer growth in vivo and in vitro. J. Funct. Foods 2014, 7, 287–297. [Google Scholar] [CrossRef]
- Chen, M.; Wang, X.; Zha, D.; Cai, F.; Zhang, W.; He, Y.; Huang, Q.; Zhuang, H.; Hua, Z.C. Apigenin potentiates trail therapy of non-small cell lung cancer via upregulating dr4/dr5 expression in a p53-dependent manner. Sci. Rep. 2016, 6, 35468. [Google Scholar] [CrossRef]
- Kuo, P.C.; Liu, H.F.; Chao, J.I. Survivin and p53 modulate quercetin-induced cell growth inhibition and apoptosis in human lung carcinoma cells. J. Biol. Chem. 2004, 279, 55875–55885. [Google Scholar] [CrossRef]
- Tsui, K.C.; Chiang, T.H.; Wang, J.S.; Lin, L.J.; Chao, W.C.; Chen, B.H.; Lu, J.F. Flavonoids from gynostemma pentaphyllum exhibit differential induction of cell cycle arrest in h460 and a549 cancer cells. Molecules 2014, 19, 17663–17681. [Google Scholar] [CrossRef]
- Elango, R.; Athinarayanan, J.; Subbarayan, V.P.; Lei, D.K.Y.; Alshatwi, A.A. Hesperetin induces an apoptosis-triggered extrinsic pathway and a p53- independent pathway in human lung cancer h522 cells. J. Asian Nat. Prod. Res. 2018, 20, 559–569. [Google Scholar] [CrossRef]
- Cai, X.; Ye, T.; Liu, C.; Lu, W.; Lu, M.; Zhang, J.; Wang, M.; Cao, P. Luteolin induced g2 phase cell cycle arrest and apoptosis on non-small cell lung cancer cells. Toxicol. Vitr. Int. J. Publ. Assoc. Bibra 2011, 25, 1385–1391. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.Q.; Li, M.H.; Qin, Y.M.; Jiang, H.Y.; Zhang, X.; Wu, M.H. Luteolin inhibits tumorigenesis and induces apoptosis of non-small cell lung cancer cells via regulation of microrna-34a-5p. Int. J. Mol. Sci. 2018, 19, 447. [Google Scholar] [CrossRef]
- Ma, L.; Peng, H.; Li, K.; Zhao, R.; Li, L.; Yu, Y.; Wang, X.; Han, Z. Luteolin exerts an anticancer effect on nci-h460 human non-small cell lung cancer cells through the induction of sirt1-mediated apoptosis. Mol. Med. Rep. 2015, 12, 4196–4202. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.Y.; Park, C.; Hwang, H.J.; Kim, G.Y.; Choi, B.T.; Kim, W.J.; Choi, Y.H. Naringenin up-regulates the expression of death receptor 5 and enhances trail-induced apoptosis in human lung cancer a549 cells. Mol. Nutr. Food Res. 2011, 55, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.J.; Qiao, K.S.; Sun, P.; Chen, P.; Li, Q. Study of egcg induced apoptosis in lung cancer cells by inhibiting pi3k/akt signaling pathway. Eur. Rev. Med Pharmacol. Sci. 2018, 22, 4557–4563. [Google Scholar] [PubMed]
- Zhang, L.; Ma, X.; Dong, Y. Effect of genistein on apoptosis of lung adenocarcinoma a549 cells and expression of apoptosis factors. J. B.U. Off. J. Balk. Union Oncol. 2018, 23, 641–646. [Google Scholar]
- Ravishankar, D.; Rajora, A.K.; Greco, F.; Osborn, H.M. Flavonoids as prospective compounds for anti-cancer therapy. Int. J. Biochem. Cell Biol. 2013, 45, 2821–2831. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.; Gao, W.; Zhang, L.; Pan, Y.; Zhang, S.; Wang, Y. Insights on structural characteristics and ligand binding mechanisms of cdk2. Int. J. Mol. Sci. 2015, 16, 9314–9340. [Google Scholar] [CrossRef] [PubMed]
- Ramteke, P.; Yadav, U.C.S. Hesperetin, a Citrus bioflavonoid, prevents IL-1β-induced inflammation and cell proliferation in lung epithelial A549 cells. Indian J. Exp. Biol. 2019, 57, 7–14. Available online: http://nopr.niscair.res.in/handle/123456789/45568 (accessed on 24 June 2019).
- Kim, M.H.; Jeong, Y.J.; Cho, H.J.; Hoe, H.S.; Park, K.K.; Park, Y.Y.; Choi, Y.H.; Kim, C.H.; Chang, H.W.; Park, Y.J.; et al. Delphinidin inhibits angiogenesis through the suppression of hif-1alpha and vegf expression in a549 lung cancer cells. Oncol. Rep. 2017, 37, 777–784. [Google Scholar] [CrossRef]
- Pal, H.C.; Sharma, S.; Strickland, L.R.; Agarwal, J.; Athar, M.; Elmets, C.A.; Afaq, F. Delphinidin reduces cell proliferation and induces apoptosis of non-small-cell lung cancer cells by targeting egfr/vegfr2 signaling pathways. PLoS ONE 2013, 8, e77270. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Chen, H.; Sun, X. Baicalein suppresses non small cell lung cancer cell proliferation, invasion and notch signaling pathway. Cancer Biomark. 2018, 22, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.; So, Y.; Jeon, H.; Jeong, M.H.; Choi, H.K.; Ryu, S.H.; Lee, S.W.; Yoon, H.G.; Choi, K.C. Tgf-beta1-induced epithelial-mesenchymal transition and acetylation of smad2 and smad3 are negatively regulated by egcg in human a549 lung cancer cells. Cancer Lett. 2013, 335, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Irimie, A.I.; Braicu, C.; Sonea, L.; Zimta, A.A.; Cojocneanu-Petric, R.; Tonchev, K.; Mehterov, N.; Diudea, D.; Buduru, S.; Berindan-Neagoe, I. A looking-glass of non-coding rnas in oral cancer. Int. J. Mol. Sci. 2017, 18, 2620. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Zimta, A.A.; Gulei, D.; Olariu, A.; Berindan-Neagoe, I. Comprehensive analysis of circular rnas in pathological states: Biogenesis, cellular regulation, and therapeutic relevance. Cell. Mol. Life Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Catana, C.; Calin, G.A.; Berindan-Neagoe, I. Ncrna combined therapy as future treatment option for cancer. Curr. Pharm. Des. 2014, 20, 6565–6574. [Google Scholar] [CrossRef] [PubMed]
- Berindan-Neagoe, I.; Calin, G.A. Molecular pathways: Micrornas, cancer cells, and microenvironment. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2014, 20, 6247–6253. [Google Scholar] [CrossRef]
- Braicu, C.; Pileczki, V.; Irimie, A.; Berindan-Neagoe, I. P53sirna therapy reduces cell proliferation, migration and induces apoptosis in triple negative breast cancer cells. Mol. Cell. Biochem. 2013, 381, 61–68. [Google Scholar] [CrossRef]
- Seles, M.; Hutterer, G.C.; Kiesslich, T.; Pummer, K.; Berindan-Neagoe, I.; Perakis, S.; Schwarzenbacher, D.; Stotz, M.; Gerger, A.; Pichler, M. Current insights into long non-coding rnas in renal cell carcinoma. Int. J. Mol. Sci. 2016, 17, 573. [Google Scholar] [CrossRef]
- De la Parra, C.; Castillo-Pichardo, L.; Cruz-Collazo, A.; Cubano, L.; Redis, R.; Calin, G.A.; Dharmawardhane, S. Soy isoflavone genistein-mediated downregulation of mir-155 contributes to the anticancer effects of genistein. Nutr. Cancer 2016, 68, 154–164. [Google Scholar] [CrossRef]
- Bhardwaj, V.; Mandal, A.K.A. Next-generation sequencing reveals the role of epigallocatechin-3-gallate in regulating putative novel and known micrornas which target the mapk pathway in non-small-cell lung cancer a549 cells. Molecules 2019, 24, 368. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhang, X.; Shi, J. Mir-98 inhibits cell proliferation and invasion of non-small cell carcinoma lung cancer by targeting pak1. Int. J. Clin. Exp. Med. 2015, 8, 20135–20145. [Google Scholar] [PubMed]
- Zhou, D.H.; Wang, X.; Feng, Q. Egcg enhances the efficacy of cisplatin by downregulating hsa-mir-98-5p in nsclc a549 cells. Nutr. Cancer 2014, 66, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Bian, S.; Yang, C.S. Green tea polyphenol egcg suppresses lung cancer cell growth through upregulating mir-210 expression caused by stabilizing hif-1alpha. Carcinogenesis 2011, 32, 1881–1889. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zang, A.; Jia, Y.; Shang, Y.; Zhang, Z.; Ge, K.; Zhang, J.; Fan, W.; Wang, B. Genistein inhibits a549 human lung cancer cell proliferation via mir-27a and met signaling. Oncol. Lett. 2016, 12, 2189–2193. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Diaz, T.; Gallardo, E.; Vinolas, N.; Marrades, R.M.; Gel, B.; Campayo, M.; Quera, A.; Bandres, E.; Garcia-Foncillas, J.; et al. Prognostic implications of mir-16 expression levels in resected non-small-cell lung cancer. J. Surg. Oncol. 2011, 103, 411–415. [Google Scholar] [CrossRef]
- Andriani, F.; Majorini, M.T.; Mano, M.; Landoni, E.; Miceli, R.; Facchinetti, F.; Mensah, M.; Fontanella, E.; Dugo, M.; Giacca, M.; et al. Mir-16 regulates the pro-tumorigenic potential of lung fibroblasts through the inhibition of hgf production in an fgfr-1- and mek1-dependent manner. J. Hematol. Oncol. 2018, 11, 45. [Google Scholar] [CrossRef]
- Sonoki, H.; Sato, T.; Endo, S.; Matsunaga, T.; Yamaguchi, M.; Yamazaki, Y.; Sugatani, J.; Ikari, A. Quercetin decreases claudin-2 expression mediated by up-regulation of microrna mir-16 in lung adenocarcinoma a549 cells. Nutrients 2015, 7, 4578–4592. [Google Scholar] [CrossRef]
- Fernandez, S.; Risolino, M.; Mandia, N.; Talotta, F.; Soini, Y.; Incoronato, M.; Condorelli, G.; Banfi, S.; Verde, P. Mir-340 inhibits tumor cell proliferation and induces apoptosis by targeting multiple negative regulators of p27 in non-small cell lung cancer. Oncogene 2015, 34, 3240–3250. [Google Scholar] [CrossRef]
- Brito, A.F.; Ribeiro, M.; Abrantes, A.M.; Pires, A.S.; Teixo, R.J.; Tralhao, J.G.; Botelho, M.F. Quercetin in cancer treatment, alone or in combination with conventional therapeutics? Curr. Med. Chem. 2015, 22, 3025–3039. [Google Scholar] [CrossRef]
- Petrushev, B.; Tomuleasa, C.; Soritau, O.; Aldea, M.; Pop, T.; Susman, S.; Kacso, G.; Berindan, I.; Irimie, A.; Cristea, V. Metformin plus piaf combination chemotherapy for hepatocellular carcinoma. Exp. Oncol. 2012, 34, 17–24. [Google Scholar] [PubMed]
- Grozav, A.; Balacescu, O.; Balacescu, L.; Cheminel, T.; Berindan-Neagoe, I.; Therrien, B. Synthesis, anticancer activity, and genome profiling of thiazolo arene ruthenium complexes. J. Med. Chem. 2015, 58, 8475–8490. [Google Scholar] [CrossRef]
- Jiang, P.; Wu, X.; Wang, X.; Huang, W.; Feng, Q. Neat1 upregulates egcg-induced ctr1 to enhance cisplatin sensitivity in lung cancer cells. Oncotarget 2016, 7, 43337–43351. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Jiao, Y.; Xue, J.; Zhang, Q.; Yang, H.; Xing, L.; Chen, G.; Wu, J.; Zhang, S.; Zhu, W.; et al. Metformin sensitizes non-small cell lung cancer cells to an epigallocatechin-3-gallate (egcg) treatment by suppressing the nrf2/ho-1 signaling pathway. Int. J. Biol. Sci. 2017, 13, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lee, E.J.; Min, K.H.; Hur, G.Y.; Lee, S.H.; Lee, S.Y.; Kim, J.H.; Shin, C.; Shim, J.J.; In, K.H.; et al. Quercetin enhances chemosensitivity to gemcitabine in lung cancer cells by inhibiting heat shock protein 70 expression. Clin. Lung Cancer 2015, 16, e235–e243. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Hwang, I.S.; Lee, Y.J.; Lee, C.H.; Kim, S.H.; Nam, H.S.; Choi, Y.J.; Lee, S.H. Knockdown of bcl-xl enhances growth-inhibiting and apoptosis-inducing effects of resveratrol and clofarabine in malignant mesothelioma h-2452 cells. J. Korean Med. Sci. 2014, 29, 1464–1472. [Google Scholar] [CrossRef]
- Nie, P.; Hu, W.; Zhang, T.; Yang, Y.; Hou, B.; Zou, Z. Synergistic induction of erlotinib-mediated apoptosis by resveratrol in human non-small-cell lung cancer cells by down-regulating survivin and up-regulating puma. Cell. Physiol. Biochem. 2015, 35, 2255–2271. [Google Scholar] [CrossRef]
- Dai, X.; Li, R.Z.; Jiang, Z.B.; Wei, C.L.; Luo, L.X.; Yao, X.J.; Li, G.P.; Leung, E.L. Honokiol inhibits proliferation, invasion and induces apoptosis through targeting lyn kinase in human lung adenocarcinoma cells. Front. Pharmacol. 2018, 9, 558. [Google Scholar] [CrossRef]
- Klimaszewska-Wisniewska, A.; Halas-Wisniewska, M.; Tadrowski, T.; Gagat, M.; Grzanka, D.; Grzanka, A. Paclitaxel and the dietary flavonoid fisetin: A synergistic combination that induces mitotic catastrophe and autophagic cell death in a549 non-small cell lung cancer cells. Cancer Cell Int. 2016, 16, 10. [Google Scholar] [CrossRef]
- You, S.; Li, R.; Park, D.; Xie, M.; Sica, G.L.; Cao, Y.; Xiao, Z.Q.; Deng, X. Disruption of stat3 by niclosamide reverses radioresistance of human lung cancer. Mol. Cancer Ther. 2014, 13, 606–616. [Google Scholar] [CrossRef]
- Zhang, Z.; Jin, F.; Lian, X.; Li, M.; Wang, G.; Lan, B.; He, H.; Liu, G.D.; Wu, Y.; Sun, G.; et al. Genistein promotes ionizing radiation-induced cell death by reducing cytoplasmic bcl-xl levels in non-small cell lung cancer. Sci. Rep. 2018, 8, 328. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Qi, B.; Xiaoxiang, W.; Xu, J.; Liu, X. Baicalein increases cisplatin sensitivity of a549 lung adenocarcinoma cells via pi3k/akt/nf-kappab pathway. Biomed. Pharmacother. 2017, 90, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wu, Q.; Chen, Y.; Zhang, J.; Li, H.; Yang, Z.; Yang, Y.; Deng, Y.; Zhang, L.; Liu, B. Diosmetin induces apoptosis and enhances the chemotherapeutic efficacy of paclitaxel in non-small cell lung cancer cells via nrf2 inhibition. Br. J. Pharmacol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, H.S.; Koo, T.H. Heavy metal concentrations in three shorebird species from okgu mudflat, gunsan, korea. Ecotoxicology 2009, 18, 61–68. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
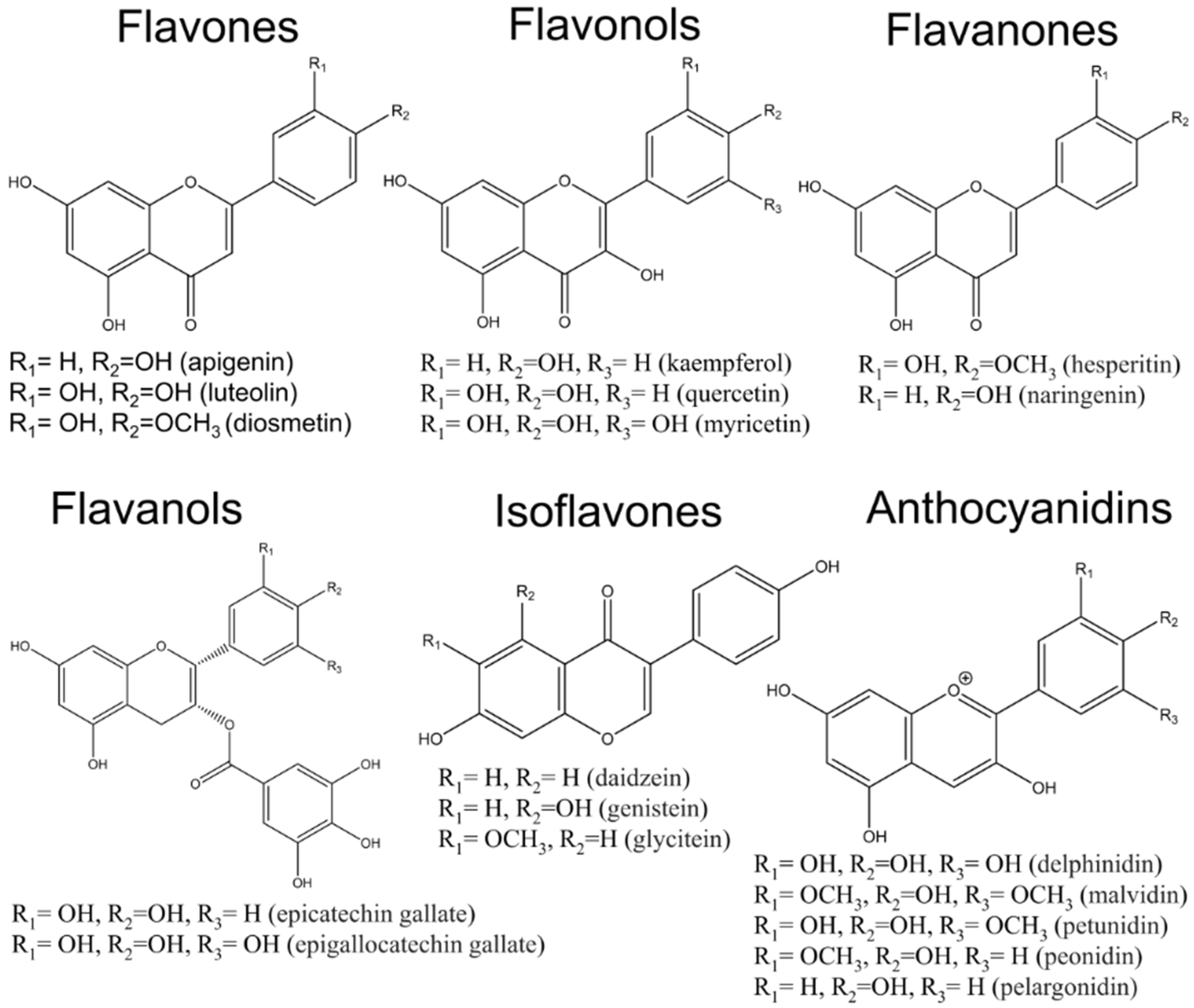
| Class of Flavonois | Representants | Source | Effect on Cells | References |
|---|---|---|---|---|
| Flavones | Apigenin, luteolin and diosmetin | Parsley, celery leaf, pepper, broccoli | Anti-inflammatory, activation of apoptosis, anti-proliferation, anti-migration, and anti-invasion effects | [21,31,32,33] |
| Flavonols | kaempherol, quercetin and myricetin | Brussel sprouts, apples, onion, leek and beans | Anti-inflammatory, antiproliferative effect, activation of apoptosis and autophagy cell adhesion, invasion and metastasis | [23,24,25,34] |
| Flavanons | hesperitin, naringenin | Citrus | Anti-inflammatory effects, Inhibition of cell proliferation, activation of apoptosis | [26,35] |
| Flavanols | catechin, epicatechin gallate, epigallocatechin, epigallocatechin gallate | apples, pears, grapes, berries green tea and cocoa | suppressing proliferation, inducing apoptosis, inhibition of EMT | [36] |
| Isoflavons | daidzein, genistein, glycitein | Soy products | Inducing apoptosis, cell cycle arrest, targeting tyrosine kinase inhibitors | [29,37] |
| Anthocyanidins | delphinidin, malvidin, petunidin, peonidin, pelargonidin | black berries, black currant and blue berries | Anti-inflammatory effect, Inhibition of proliferation, activation of apoptosis | [30] |
| Phycochemical Class | Phytochemicals | Dose | In Vitro Models | End-Point | Effects | Molecular Target | Reference |
|---|---|---|---|---|---|---|---|
| Flavones | Lutein | 20–160 μM | NCI-H460, HEK-293T cell line | Apoptosis assay, western blotting, RT-PCR | Apoptosis activation | Bad↓, Bcl-2↓, Bax↓, caspase-3↓ and Sirt1↓ | [63] |
| 0–100 μM | H1299 and -H460 cells | Immunoblot analysis, PI assay | Apoptosis activation | p38/ROS/caspase cascade↑ | [33] | ||
| Apigenin | 0–100 μM | A549 cells | MTT, colony assay, Transwell assay, western blot | anti-proliferation, anti-migration, and anti-invasion effects | ↓Akt affecting PI3K signaling | [31] | |
| 5–80 μM | A549, H1975, and HCC827 NSCLC cell lines | Transwell migration and invasion assays, RT-PCR | Inhibition of the migration/invasion of NSCLC cells | Akt and Snail/Slug ↑ | [32] | ||
| Baicalein | 0–100 μM | A549 and H1299 cells | Western blot, QRT-PCR | Inhibition of cell proliferation, down-regulation of Notch1 and hes-1 expression | Cyclin D1 and CDK1↓ | [72] | |
| Flavonols | Quercertin | 0–200 µM | JB6 Cl41 cells and A549 | Anchorage-independent transformation assay, Microscale thermophoresis | Suppression of cells proliferation | Aurora B kinase↓ | [23] |
| 0–200 µM | NCI-H358 and A549 cells. | Apoptosis, microarray | antiproliferative effect | Caspase-3↑ | [50] | ||
| Kaempferol | 1–50 μM | A549 cell line | MTT test, Transfection, PCR | inhibition of cell proliferation | STAT3↓, claudin-2↓ | [24] | |
| Fisetin | 0–40 μM | A549 cells | MTT test, RT-qPCR, flow-cytometry | Apoptosis activation | ERK1/2↓ | [46] | |
| Flavnones | Hesperetin | 0–100 μM | A549 cells | RT-PCR, western blot | inhibition of cell proliferation | ERK1/2↓, HFKb-p65↓ | [69] |
| 50–125 µg/mL | A549 cells | inhibition of cell proliferation and cell cycle progression | [35] | ||||
| Naringenin | 0–300 µM | A 549 cells | RT-PCR, western blot | suppression of Akt activity and the downregulation of MMP-2 and -9 | Akt↓ | [48] | |
| 0–500 µM | A 549 cells | Apoptosis assay, western blot | Cells apoptosis | Bid and DR5↑ | [64] | ||
| Flavanols | EGCG | 0–40 µM | H1299 cells | Cell proliferation, apoptosis assay, western blot | suppressing proliferation, inducing apoptosis | PI3K/Akt↓ | [65] |
| 0–40 µM | A549 cells | Cell culture and transfection, Western blot, Flow cytometry | Decreased EGF-induced EGFR, Akt and ERK1/2 activation. | EGFR↓ | [45] | ||
| EGCG | 0–40 µM | A549 and NCI-H1299 cells | Scattering assay, wound healing assay, in vitro invasion assay, qRT-PCR, Western blot, confocal microscopy | Cell proliferation, EMT | TGFβ↓, Smad2↓ and Erk1/2↓ | [36] | |
| EGCG | 0–100 µM | A549 cells | HAT activity assays, Immunoprecipitation and western blot analysis, RT-PCR | TGF-β1-induced EMT inhibition | TGFβ1↓, Smad2↓, Smad3↓ | [73] | |
| Isoflavones | Daidzein | 0–60 µM | NSCLC cells | TUNEL assay. Real-time PCR and western blotting | Inducing apoptosis | STK3↓, STK4↓, YAP1↓, caspase3↓ | [49] |
| Genistein | 0–100 µM | H446 cells | Apoptosis assay, colony assay, RT-PCR, western blot | apoptosis and G2/M phase cell cycle arrest | Cdc25B↓, cyclin B1↓, survivin↓ | [37] | |
| 0–100 µM | A549 cells | Apoptosis assay, qRT-PCR, Western blot | Inhibition of cell proliferation, cells apoptosis | Bax↑, Bcl-2↓ | [66] | ||
| Anthocyanidins | Delphinidin | 0–80 µM | A549 cells | Cell viability assay, Western blot, RT-PCR, Matrigel plug assay | suppression of the ERK, mTOR, and p70S6K pathways | HIF-1α↓ VEGF↓ | [70] |
| 5–60 µM | NCI-H441, SK-MES-1 and A549 | Western blot, Cell viability assay | Apoptosis and angiogenesis | ↑caspase-3/9, ↓ anti-apoptotic proteins (Bcl2, Bcl-xL and Mcl-1), ↑pro-apoptotic proteins (Bax and Bak);↓EGFR and VEGFR2 | [71] |
| Natural Compound | Preclinical Model | miRNA Targeted | Gene Targeted | Target Mechanism | Technology Approach | References |
|---|---|---|---|---|---|---|
| EGCG (0–50 µM) | CL13 cells, H1299, H460 and A549 | miR-210 (↑) | HIF-1α (↓) | reduction of cell growth, hypoxia | Microarray, RT-PCR | [84] |
| A549 cells | miR-212 (↓) miR-155 (↑) | MAPK | inhibition of proliferation and migration | NGS | [81] | |
| Quercetin | A549 cells | miR-16 (↑) | Claudin-2 (↓) | Decrease of tight junction mechanisms | qRT-PCR | [88] |
| Genistein | A549 cells | miR-27a (↓) | MET (↑) | apoptosis and promotes caspase-3/9 activation | Apoptosis assay, western blotting | [85] |
| Kaempferol | A549 | miR-340 (↑) | Cyclin D1 (↓), PTEN (↑) | Cell apoptosis, inhibition of proliferation | Apoptosis assay, qRT-PCR, western blotting | [34] |
| Phytochemicals | Chemotherapeutic | Biological System | Biological Effect | References |
|---|---|---|---|---|
| EGCG | Cisplatin | A549, H460 and H1299 cells | enhances cisplatin sensitivity, NEAT1 upregulates EGCG-induced CTR1 | [93] |
| Metformin | A549, H1299 and H460 human NSCLC cell lines | suppressing the Nrf2/HO-1 signaling pathway | [94] | |
| Cisplatin | H1299 and Lu99 cells | downregulation of AXK and TYRO3 receptor tyrosine kinases | [104] | |
| Quercetin | Gemcitabine | A549 and H460 cells | Inhibition of cancer cell growth and sensitized cancer cells to gemcitabine by quercetin, apoptosis via inhibiting HSP70 expression. | [95] |
| Fisetin | Paclitaxel | A549 cells | Synergic effect of combination treatment | [99] |
| Genistein | Radiotherapy | A549 cells | Stimulation of apoptosis, reducing plasmic Bcl-xL levels | [101] |
| Baicalein | Cisplatin | A549/ /CDDP cells | Induction of apoptosis via PI3K/Akt/NFκB pathway | [102] |
| Diosmetin | Paclitaxel | A549, H1299, H460, SPC-A1, H441, H1650, Calu-3 | Cells apoptosis, increasing paclitaxel efficacy, ROS accumulation, PI3K/Akt/GSK-3β/Nrf2 pathway disruption | [103] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zanoaga, O.; Braicu, C.; Jurj, A.; Rusu, A.; Buiga, R.; Berindan-Neagoe, I. Progress in Research on the Role of Flavonoids in Lung Cancer. Int. J. Mol. Sci. 2019, 20, 4291. https://doi.org/10.3390/ijms20174291
Zanoaga O, Braicu C, Jurj A, Rusu A, Buiga R, Berindan-Neagoe I. Progress in Research on the Role of Flavonoids in Lung Cancer. International Journal of Molecular Sciences. 2019; 20(17):4291. https://doi.org/10.3390/ijms20174291
Chicago/Turabian StyleZanoaga, Oana, Cornelia Braicu, Ancuta Jurj, Alexandru Rusu, Rares Buiga, and Ioana Berindan-Neagoe. 2019. "Progress in Research on the Role of Flavonoids in Lung Cancer" International Journal of Molecular Sciences 20, no. 17: 4291. https://doi.org/10.3390/ijms20174291
APA StyleZanoaga, O., Braicu, C., Jurj, A., Rusu, A., Buiga, R., & Berindan-Neagoe, I. (2019). Progress in Research on the Role of Flavonoids in Lung Cancer. International Journal of Molecular Sciences, 20(17), 4291. https://doi.org/10.3390/ijms20174291