1. Introduction
Social learning is the process in which animals gain new skills and information through interaction with or observation of other individuals, and it eliminates the need for individuals to pay the costs of acquiring information on their own. Social learning plays important roles in a wide range of contexts, including acquiring new foraging skills and food preferences. There is now strong experimental evidence in humans that the eating behavior of other people can strongly influence the amount of food that we eat [
1,
2]. The learning of foraging what, where and how to eat is the main topic in studies of behavioral innovations in birds [
3,
4].
Because of the complex foraging environments and the change of food items, seasonally or spatially, learning plays a role in the foraging flexibility of fish [
5]. Several studies have demonstrated that experience influences the foraging success, novel foraging behavior and profitability of different prey types. Groups of fish forage more efficiently than individuals, such as the goldfish (
Carassius auratus auratus), bluntnose minnows (
Pimephales notatus), Alaska pollock (
Theragra chalcogramma) and sticklebacks (
Gasterosteus aculeatus) [
6,
7,
8,
9]. The enhancement of foraging by social learning has also been reported in hatchery-reared juvenile chum salmon (
Oncorhynchus keta), Alaska pollock and rock bass (
Ambloplites rupestris) [
8,
10,
11]. In addition, there are well-documented effects of social learning on novel foraging behavior through the observation of conspecifics in fishes. Anthouard reported that European sea bass (
Dicentrarchus labrax) learned to press a lever to receive a food reward through the observation of the trained demonstrators [
12]. However, little is known about the potential role of social learning in acquiring new feeding habits in fish.
Mandarin fish (
Siniperca chuatsi) is an important aquaculture fish and highly valued due to its excellent flesh quality with an output value of more than 3 billion U.S. dollars (20 billion Yuan). Mandarin fish are aggressive piscivores; once their fry start feeding, they feed solely on live fry of other fish species and refuse zooplankton or formulated diets [
13]. The acceptance of dead prey fish or artificial diets represents a key challenge in the aquaculture of mandarin fish, similar in the Eurasiam perch [
14]. We designed a specific training paradigm to domesticate the feeding habits of mandarin fish, and we found that some individuals could accept dead prey fish successfully (feeders) [
15]. What is less clear is whether mandarin fish are aware of the influence of social learning from feeders on their feeding behavior of accepting dead prey fish as a novel feeding habit.
Observational learning, i.e., the learning of individual (observer, actor) by the direct observation of the behavior of an experienced individual (demonstrator, tutor) is the most frequent type of social learning [
16]. To investigate the effect of social learning on acquiring new feeding habits of mandarin fish, we examined the success rate of feeding habit domestication from live prey fish to dead prey fish, as well as the food intake of dead prey fish in mandarin fish with or without feeders of dead prey fish as demonstrator. In addition, the analysis of gene expressions and signaling pathways of learning through Western blotting, transcriptome sequencing, inhibitor treatment and a chromatin immunoprecipitation (ChIP) assay allowed us to gain insights into the molecular mechanism of social learning in acquiring new feeding habits in mandarin fish. These results can support aquaculture domestication programs and research in mandarin fish, and they can lay a solid genetic foundation to understand the social learning of feeding behavior.
3. Discussion
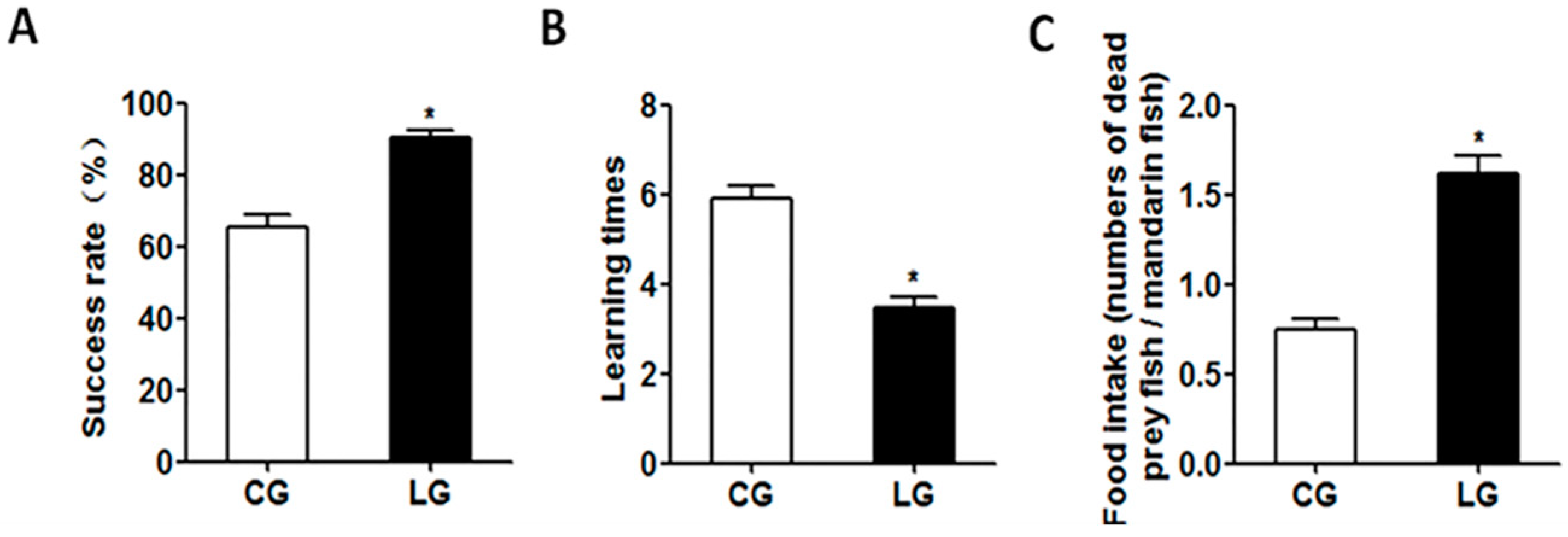
Most studies of social learning in a foraging context have been focused on the social transmission of new foraging skills [
17,
18,
19] and food preferences [
20,
21,
22]. To date, there have been few studies demonstrating learning processes in the feeding habit domestication of fish. Our study showed that mandarin fish were capable of social learning to acquire a new feeding habit. The success rates of feeding habit domestication from live prey fish to dead prey fish was significantly higher in mandarin fish with the positive demonstration fish than that with the negative demonstration fish. The food intake of dead prey fish was also increased in the learning group, and the learning times were decreased. Mandarin fish with the positive demonstration fish might be easier to be domesticated to feed on dead prey fish, suggesting that the learning ability of mandarin fish during the domestication of feeding habits. This is the first time that it has been shown that that social learning could enable mandarin fish with a naive feeding habit of live prey fish to acquire information of accepting dead prey fish from a knowledgeable one, thus avoiding the costs of learning solely by individual experience. Food preference resulting from the observation of the feeding behavior of the demonstrator has been found also in other fish. The observation of a trained conspecific Atlantic salmon
Salmo salar significantly increased the rate at which naive hatchery-reared fish accepted novel, live prey items [
23]. The young-of-the-year perch
Perca fluviatilis could finish a shift from natural food items to commercial dry feeds faster through the presence of conspecifics that are familiar with the dry feed than without the demonstrators [
24]. Previous studies have shown that changes due to experience increased the capture rate of a specific prey type and shortened the time to approach a novel food item in naive rainbow trout (
Oncorhynchus mykiss) [
25] and pink salmon (
Oncorhynchus gorbuscha) [
26]. Our results indicated that social learning could facilitate the acquisition of a novel feeding habit in mandarin fish and have a great effect on domestication and food intake. However, it is unclear how social learning could affect feeding habits, as is the molecular mechanism of social learning on food preference.
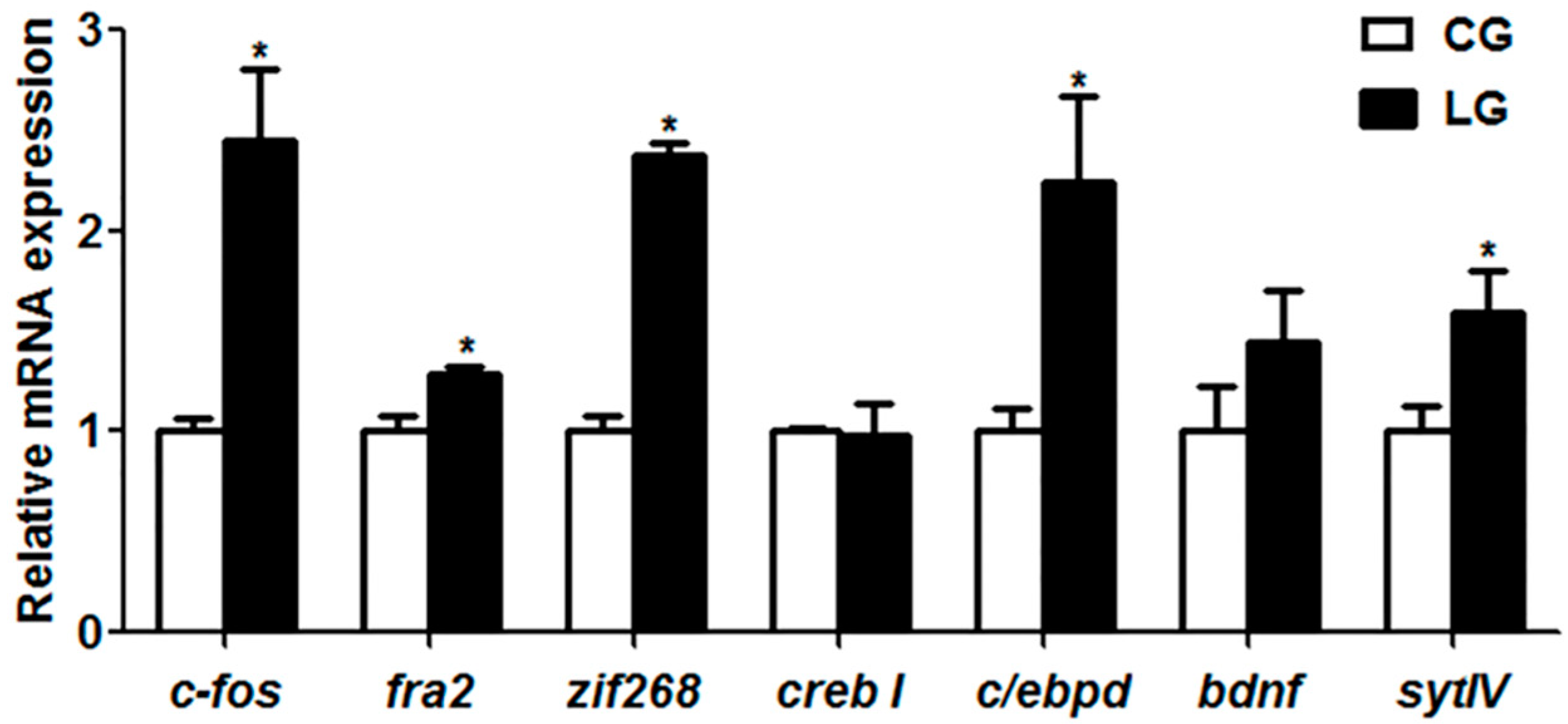
With regard to the molecular mechanism of social learning during the domestication of feeding habits in mandarin fish, we examined the mRNA expression of learning genes. In mandarin fish with the positive demonstration fish, the gene expressions of
c-fos,
fra2,
zif268,
c/ebpd and
sytIV were significantly increased, and the expression of the
creb I and
bdnf genes showed no difference between the two groups. The TORC1 (target of rapamycin complex 1)-mediated CREB regulation is a critical molecular step underlying synaptic plasticity and long-term memory [
27]. As members of immediate early gene (IEG) and the Fos family of transcription factors,
c-fos and
fra2 mRNA expressions are up-regulated in response to a variety of neuronal activation protocols, including long-term protentiation (LTP) [
28] and behavioral training [
29,
30]. The expression of the c-Fos protein is increased in the hippocampus of rats trained on the socially transmitted food preference (STFP) [
31]. The short-term memory impairment of ethanol-exposed rats is related to a decrease of
c-fos expression in the hippocampus [
32]. Besides
c-fos, another IEG gene,
zif268, is reported to be involved in learning and memory, and several studies have shown that
zif268 mRNA is upregulated during different forms of associative learning [
33,
34]. In the present study, compared with fish in the control group, the mRNA levels of
c-fos,
zif268 and
c/ebpd were dramatically increased in fish of the learning group, suggesting that the social learning to exploit new food sources in mandarin fish might be attributed to the higher expression of the
c-fos,
zif268 and
c/ebpd genes, potentially resulting in the long-term protentiation of a new feeding habit (feed on dead prey fish).
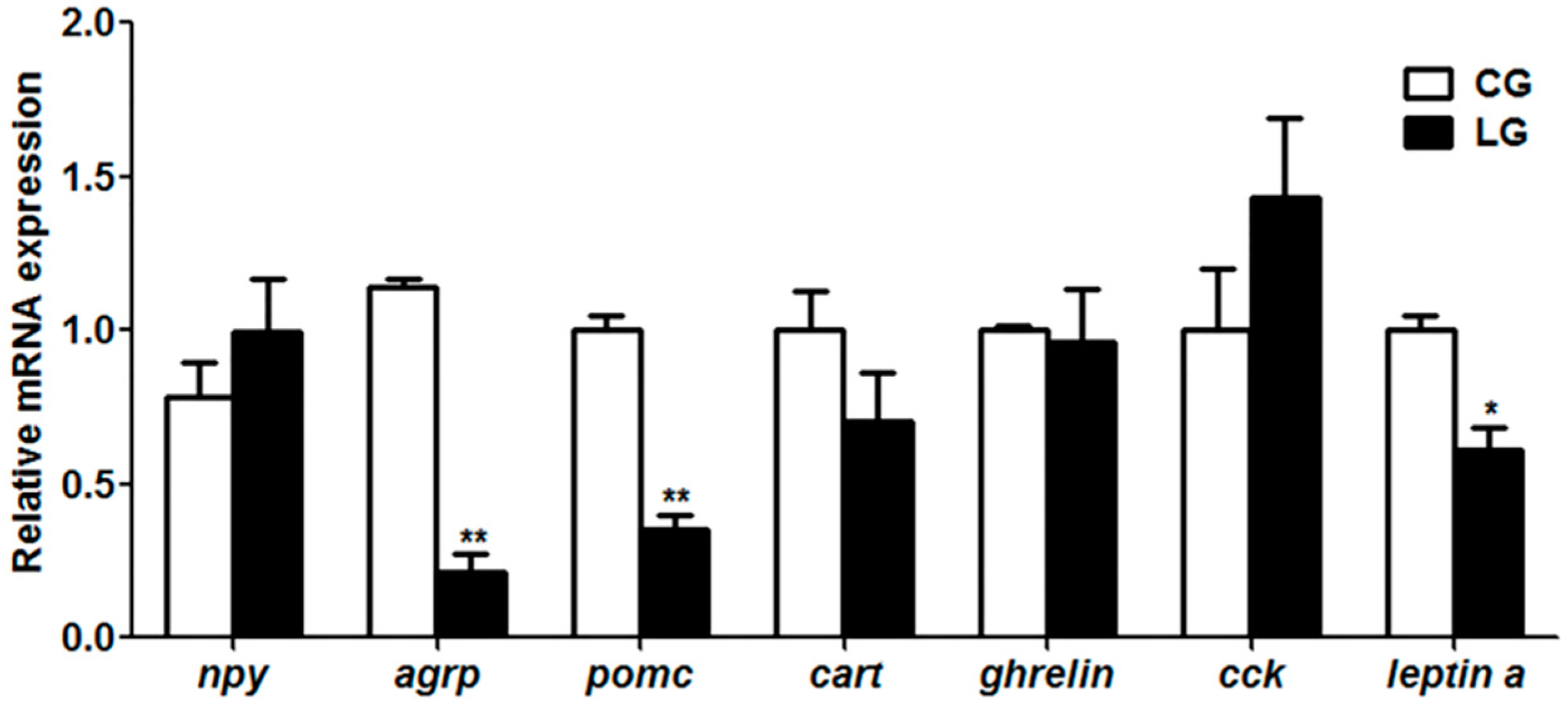
Consistent with the increase of food intake of dead prey fish, mandarin fish with the positive demonstration fish showed a decrease of anorexigenic
pomc and
leptin a. The neuropeptides
npy,
agrp,
pomc and
cart play essential roles in the regulation of food intake and energy homeostasis in mammals [
35]. Recent findings have demonstrated that peripherally-derived endocrine signals act on receptors in hippocampal neurons to regulate food intake through leptin or ghrelin and to learn food reward-driven responding [
36]. Our results suggested that the lower expression of the anorexigenic genes
pomc and
leptin a might facilitate the food intake of dead prey fish in the learning group. Furthermore, c-Fos has been considered as the learning molecule, whether the appetite control genes
pomc and
leptin a could be the downstream molecular targets of c-Fos is poorly understood.
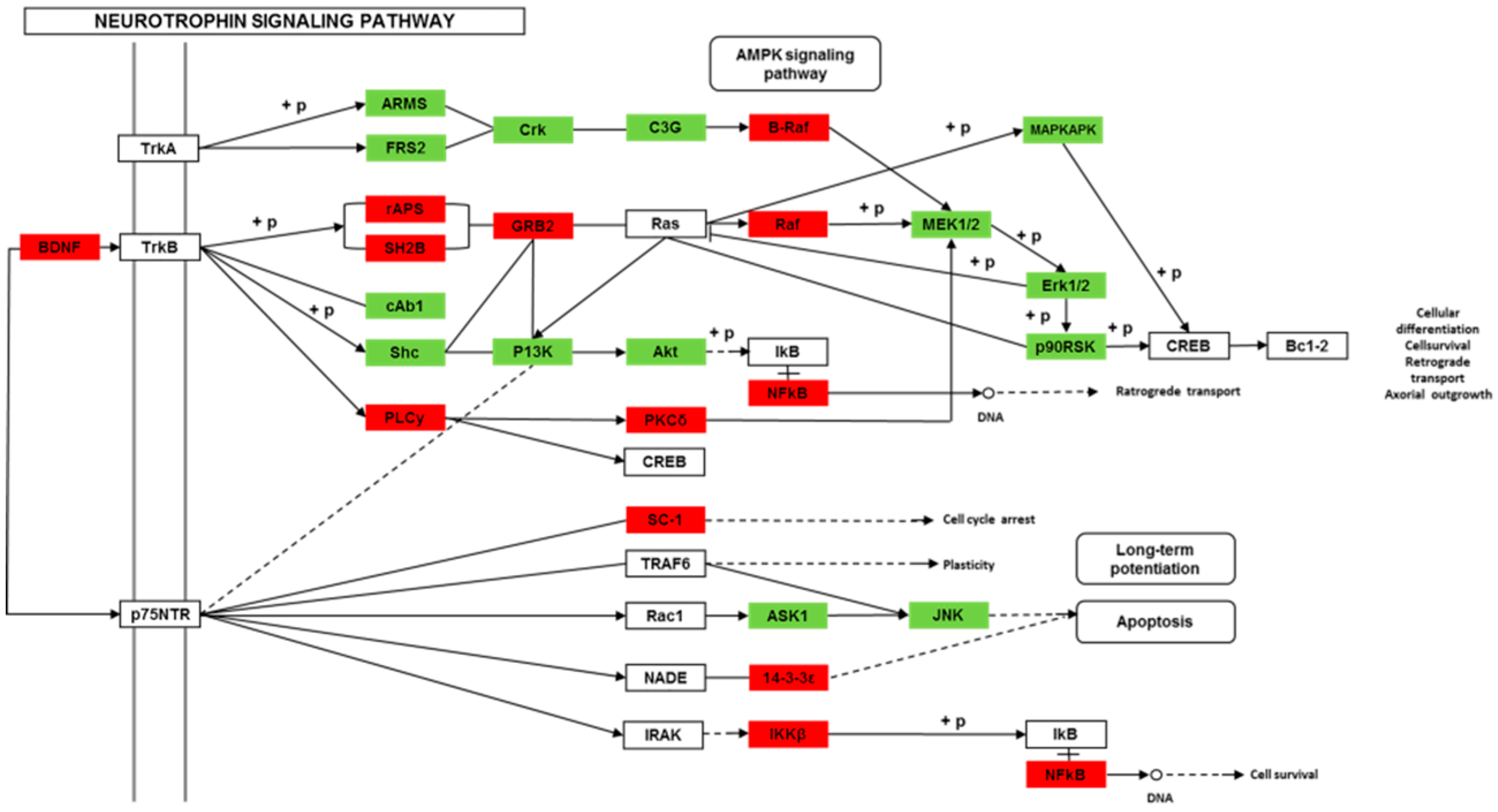
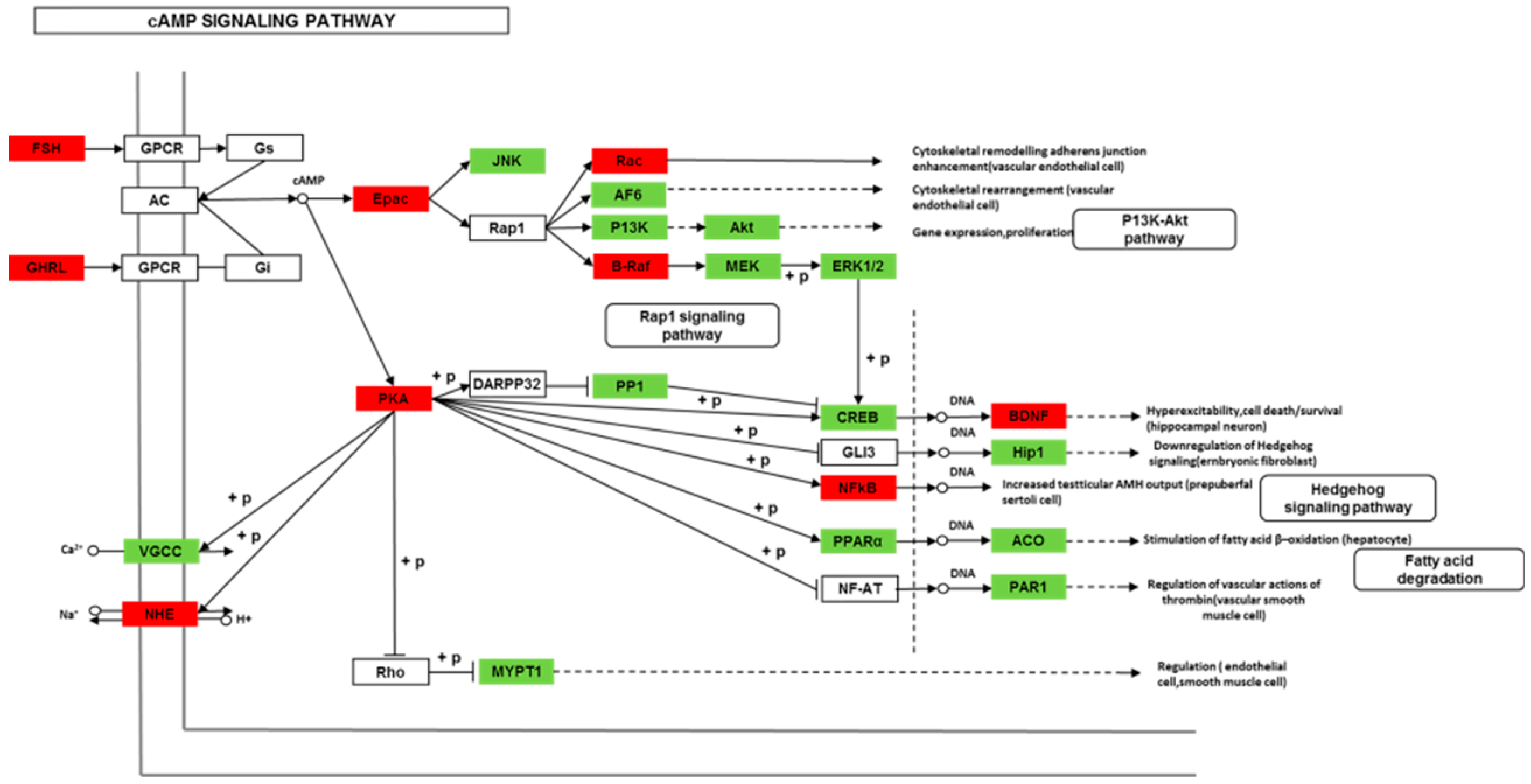
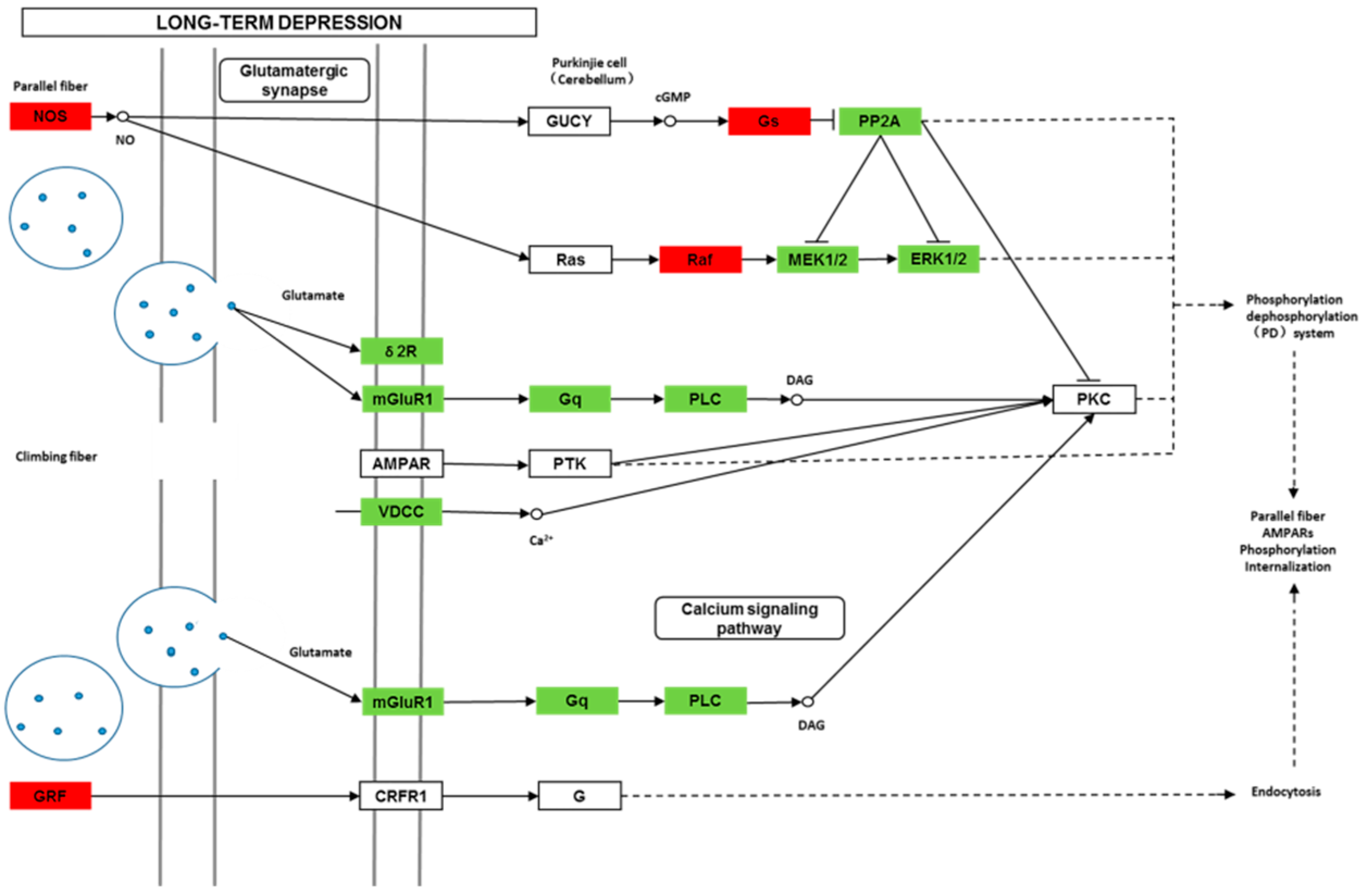
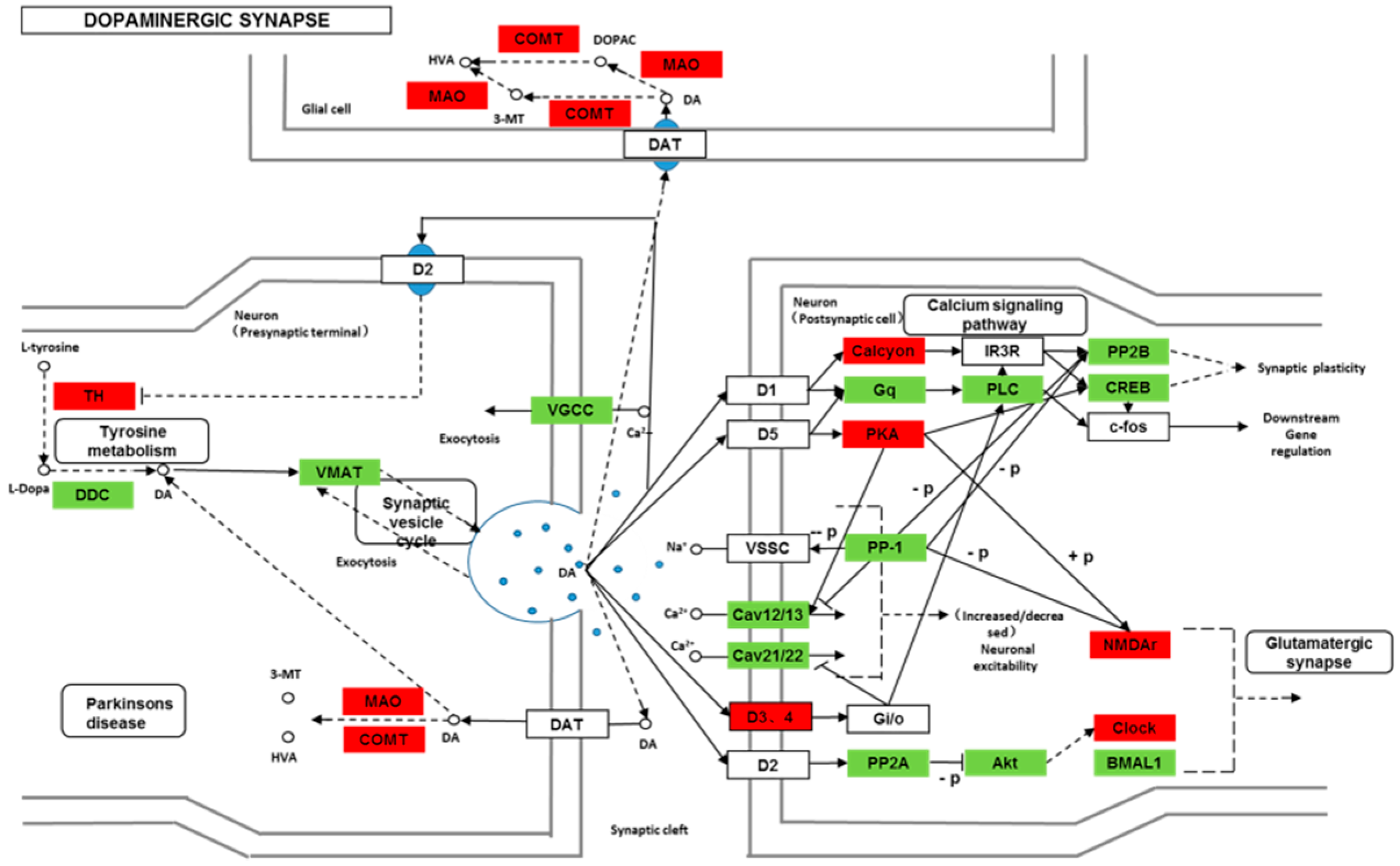
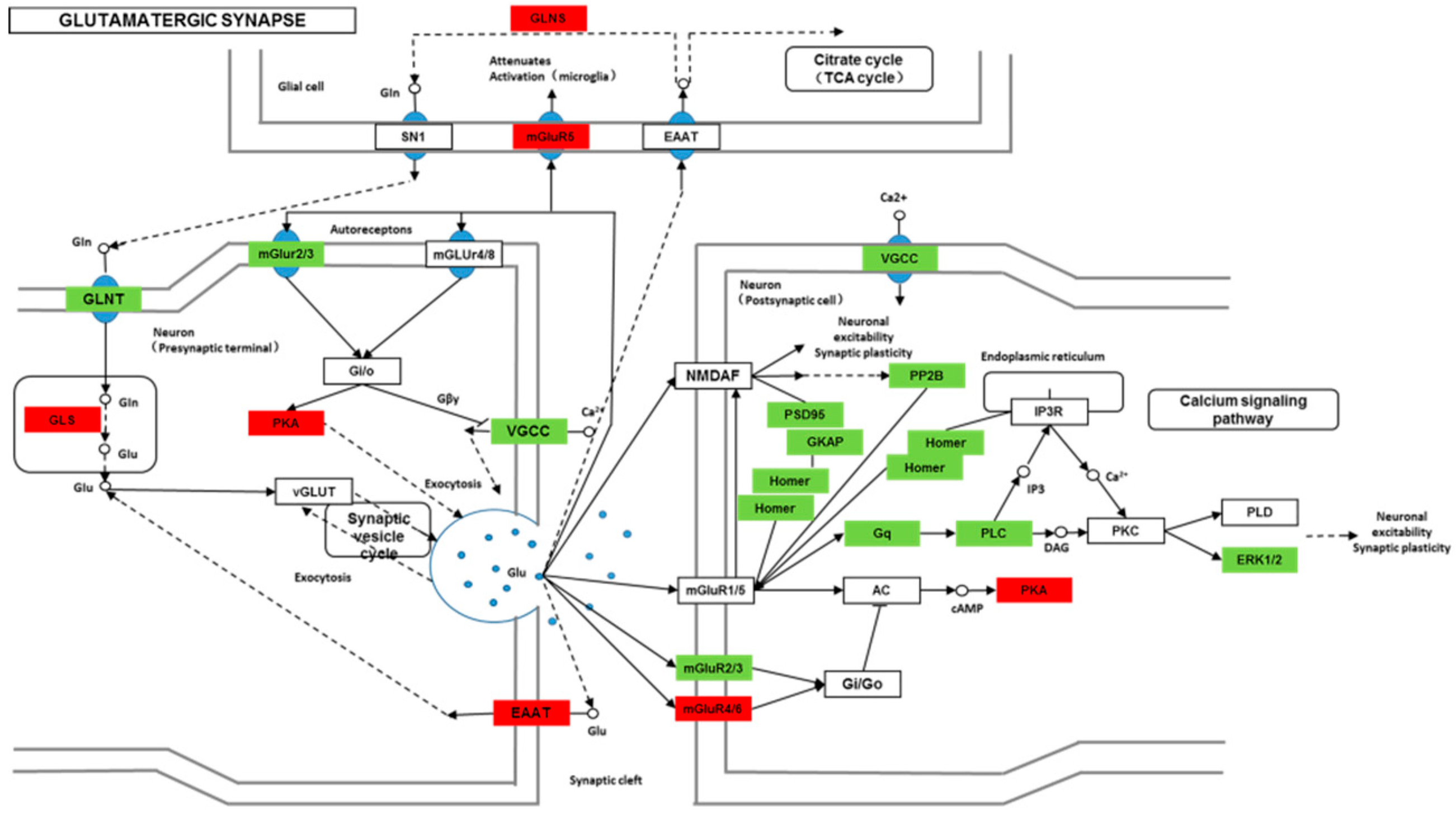
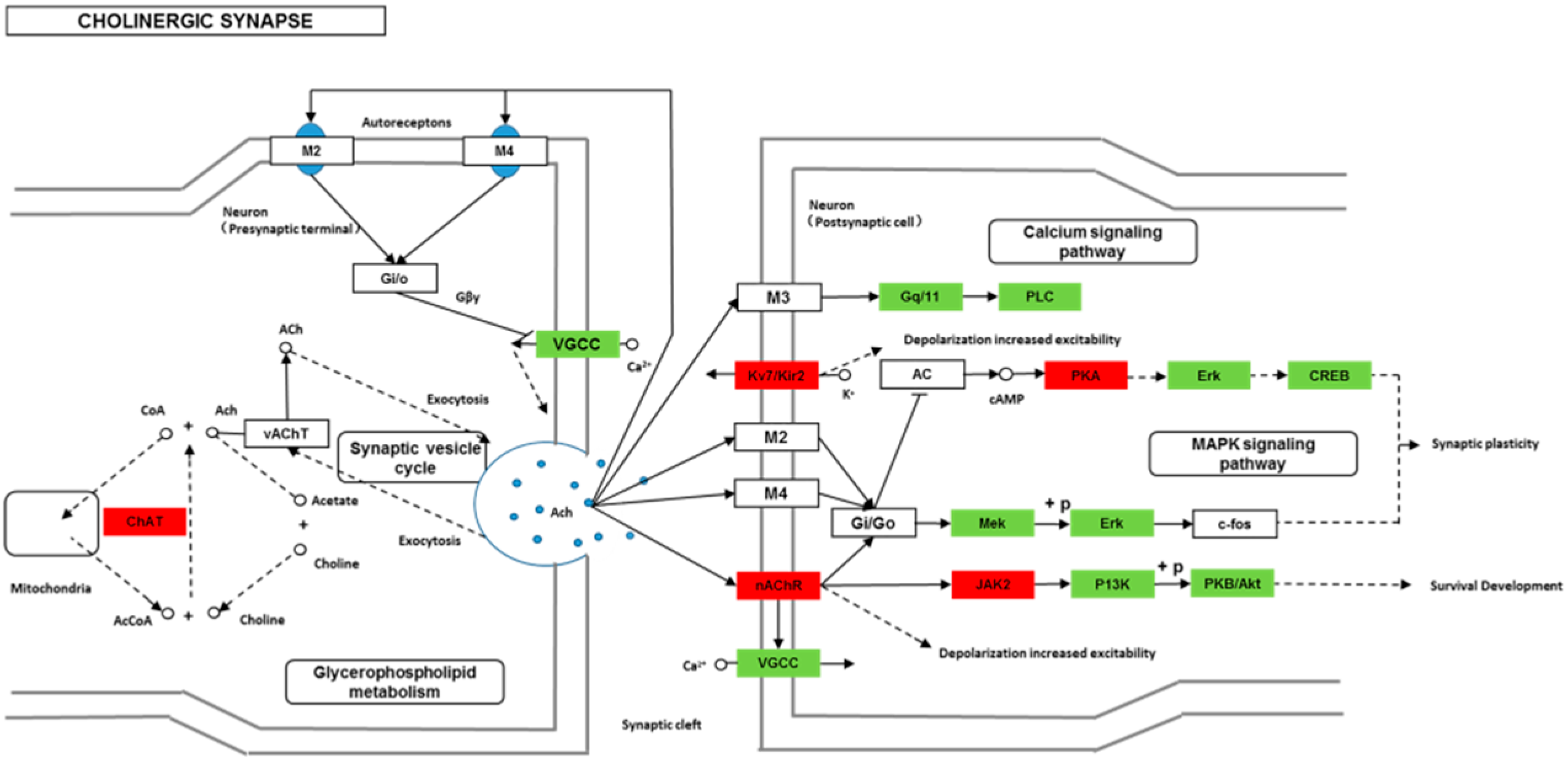
Though social learning is necessary for the domestication of feeding habits, the neural mechanisms are mostly unknown. To obtain an overview of gene expression profile in mandarin fish with or without learning, we found the differentially expressed genes between Groups CG and LG in several signaling pathways, such as the cAMP signaling, neurotrophin signaling, long-term potentiation, long-term depression, serotonergic synapse, dopaminergic synapse, glutamatergic synapse, and cholinergic synapse pathways. Learning and memory processes depend on electro-chemical signaling undertaken by amino acids, biogenic monoamines, acetylcholine, gasotransmitters, and peptides within the neuronal networks in the brain [
37]. It has been shown that dopamine is involved in the expression of activity-dependent synaptic plasticity as well as in behavioral learning and learning-associated immediate-early gene expression [
38]. In the present study, mandarin fish in the LG group showed a higher expression of D3-class receptors (D3/D4R), suggesting that the exposure to the novel feeding habit of dead prey fish might be known to release dopamine and facilitate dopamine-dependent synaptic plasticity induction via D3-class receptors. The efficacy of glutamate transmission can be persistently changed following the neuronal activities [
39] in LTP and long-term depression (LTD) [
40]. Our results also showed that the glutamatergic synapse signaling was involved in the social learning process of new feeding habits in mandarin fish through both long-term potentiation and long-term depression.
Here, to investigate the signaling pathways that lead to activation of the IEGs and the upstream targets of these genes will advance our understanding of how functional activation of
c-fos and
zif268 may be implicated in processing social learning. It remains unclear why the social learning of feeding habits in mandarin fish promoted the expression of learning related genes, such as
c-fos,
fra2,
zif268,
c/ebpd and
sytIV. The long-lasting synaptic plasticity, which is frequently investigated in LTP and in learning paradigms, is associated with several cellular events: Ca
2+ influx through the N-methyl-D-aspartate (NMDA) receptor, the generation of cAMP and the activation of PKA, the phosphorylation of mitogen-associated protein kinase (MAPK) and CREB, and the subsequent transcription of plasticity-associated genes [
28,
41]. CaMKII is also an essential component of the signaling cascade required for the regulation of synaptic plasticity and neuronal growth [
42]. The PKA signaling pathway has been proposed to participate in both the late phase of long term potentiation and protein synthesis-dependent phase of memory formation [
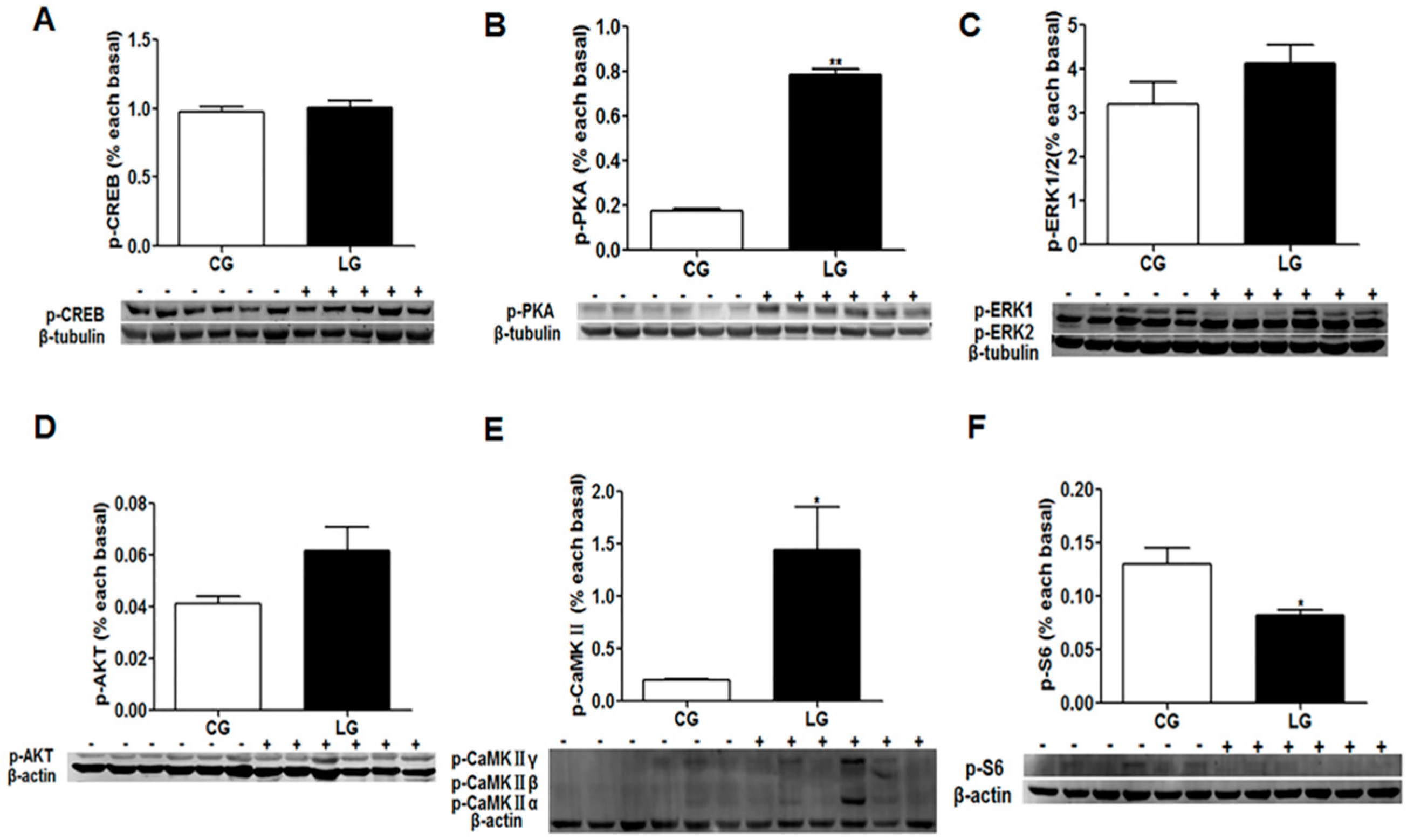
43]. Based on the current study, the increased success rate of domestication and food intake of dead prey fish could be attributed to the phosphorylation and activation of the PKA and CaMKII signaling pathway. The learning-induced phosphorylation of CREB co-occurs with the expression of the c-Fos protein [
31]. There strong evidence that the phosphorylation of CREB is important for the long term consolidation of memories and long-term memory formation [
44,
45], but not short-term memory for socially transmitted food preference [
31]. In the present study, p-CREB was measured in the brain of mandarin fish following the acquisition of a new feeding habit. The result indicated that the phosphorylation of CREB is not related to the short-term memory of new feeding habits in mandarin fish. In addition, the mechanistic target of rapamycin (mTOR) signaling is considered as an intracellular signaling for the regulation of food intake and energy homeostasis [
46]. We found that the phosphorylation level of the S6 ribosomal protein (p-S6) was significantly decreased in the learning group with the increased food intake of dead prey fish. The result was consistent the hypothesis that the inhibition of mTOR signaling might stimulate food intake through changing the translation levels of the anorexigenic Pomc protein in mandarin fish (data not published).
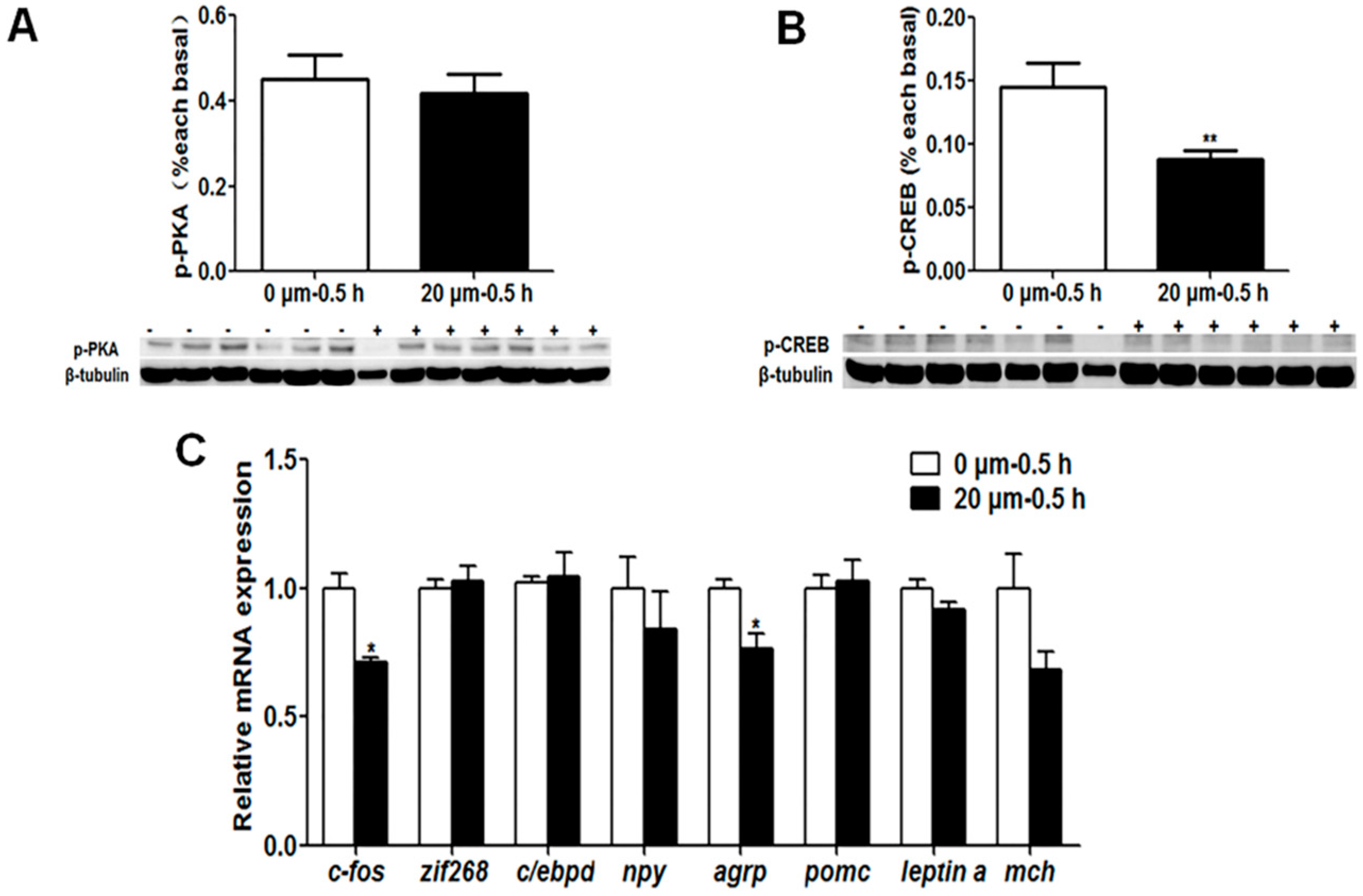
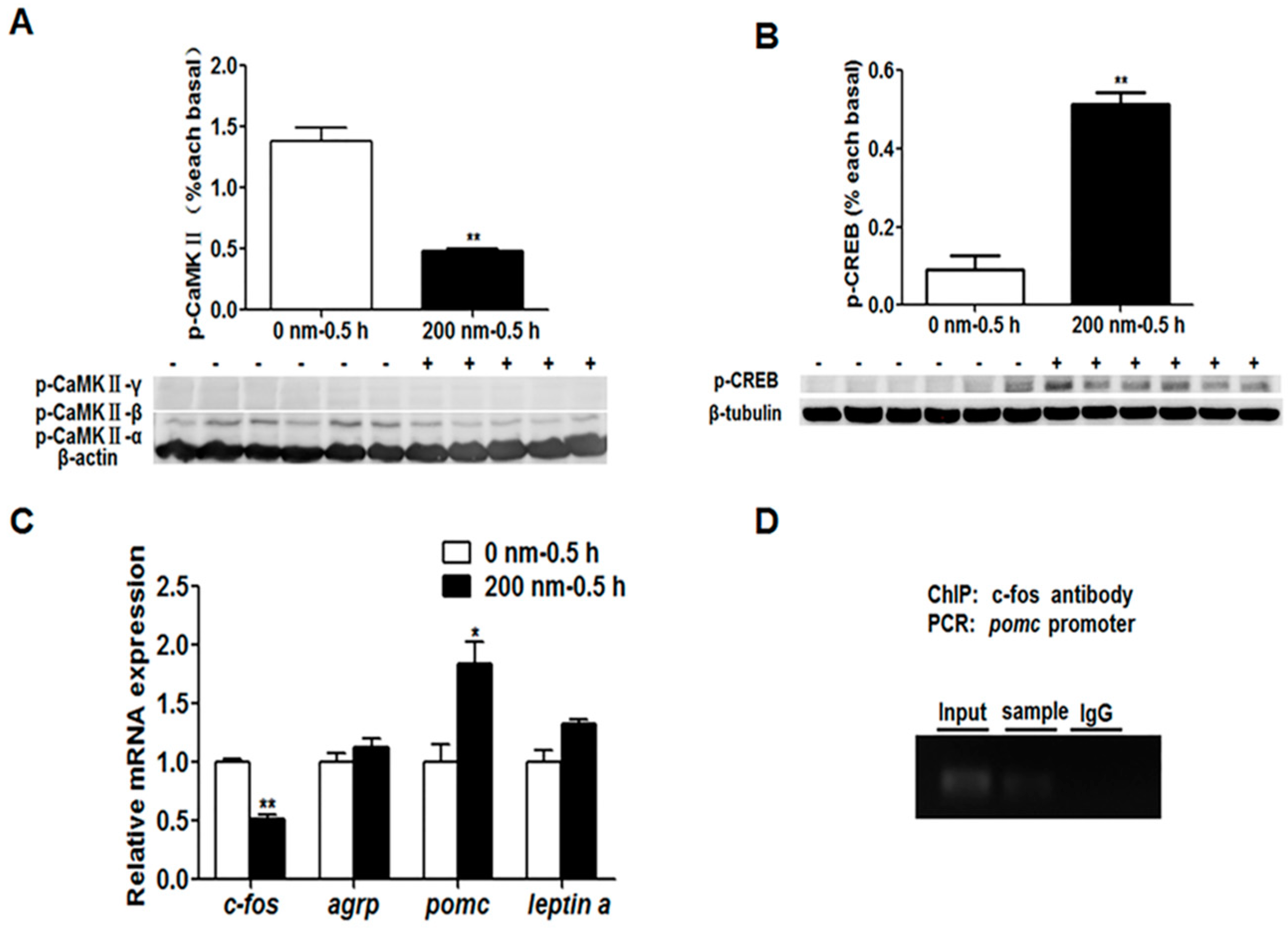
The interaction between learning genes and appetite control genes involved in the social learning of mandarin fish to accept dead prey fish through the PKA and CaMKII signaling pathways remains to be further explored. We used the inhibitors of the learning signaling pathway, H-89 for PKA and KN-62 for CaMKII, to confirm if the learning signaling pathway, especially the learning gene c-fos, could affect appetite control genes. The results showed that no significant change of the p-PKA level and mRNA expression of the zif268, c/ebpd, npy, pomc, leptin a and mch genes were observed upon the inhibitor H-89 treatment, but the p-CREB level, c-fos and agrp gene expressions were significantly decreased. Though the p-PKA level did not change with inhibitor treatment, the downstream signaling of the PKA pathway, the p-CREB level was decreased, suggesting that the PKA pathway might regulate c-fos expression through CREB signaling but not work on the anorexigenic gene expression. The p-CaMKII level and c-fos expression with KN-62 treatment were significantly lower than that without treatment, the p-CREB level and pomc expression were significantly increased, and the expression of the agrp and leptin a genes show no changes between groups. It is suggested that the CaMKII signaling pathway activated by social learning could stimulate the expression of the c-fos gene, and then c-fos might be an important transcriptional factor to inhibit the expression of the anorexigenic gene pomc, resulting the increase of food intake of dead prey fish in mandarin fish.
To determine whether c-Fos has any direct regulation on transcriptional activity of the
pomc gene, we examined the interaction between transcriptional factor c-Fos and the three potential AP-1 binding sites of the
pomc gene with a ChIP assay. We found significant enrichments at the Site 3 in c-Fos-immunoprecipitate, suggesting the binding of c-Fos to AP-1 binding sites in the regulatory region of
pomc. A previous study has shown that c-Fos functions as a proapoptotic agent by binding the c-FLIP(L) (FLICE (FADD-like IL-1β-converting enzyme) -like inhibitory protein) promoter and repressing the antiapoptotic molecule c-FLIP(L) transcriptional activity [
47]. The AP-1 family member protein c-Fos plays a crucial role in a variety of biological processes, and the downstream targets of c-Fos have been identified in a wide range of normal development, inflammation, and oncogenesis [
47]. In this report, we showed that c-Fos, in addition to its well-known oncogenic function, has a novel regulatory function on food intake by the
pomc gene. The regulation of c-Fos on
pomc might be a key pathway for feeding and food habit modification by learning, and the activator of the c-Fos/
pomc pathway could play important roles in animal culture with artificial diets, especially endangered species and economic animals, and in the formation of healthy food preferences for children, such as socially transmitted food preference.
In conclusion, we investigated the effect of social learning on the feeding habit domestication from live prey fish to dead prey fish in mandarin fish. Our results showed that with the positive demonstration fish that feed on the dead prey fish, mandarin fish also could accept the dead prey fish with a higher success rate and lower learning times than those with the negative demonstration fish. The social learning of acquiring new feeding habits in mandarin fish could be attributed to the expression of learning and appetite control genes, as well as the activation of the CaMKII signaling pathway. The CaMKII signaling pathway was activated by social learning and the stimulation of the expression of the c-fos gene. As such, the c-fos gene might be an important transcriptional factor to inhibit the expression of the anorexigenic gene pomc, resulting in an increase of the food intake of dead prey fish in mandarin fish. This offers a first insight into the ability and molecular mechanism of social learning on acquiring novel feeding habits in fish. As the ability to copy the feeding behavior from intraspecific tutors, our experiment encourages further studies on the potential roles of social learning in the context of feeding training in fish, especially for large-scale aquaculture purpose.
4. Materials and Methods
4.1. Reagents and Fish
Phospho-CREB (Ser133) (87G3) Rabbit mAb, Phospho-PKA C (Thr197) (D45D3) Rabbit mAb, Phospho-p44/42 MAPK (Erk1/2) (Thr202/Tyr204) (D13.14.4E) XP® Rabbit mAb, Phospho-CaMKII (Thr286) (D21E4) Rabbit mAb, Phospho-S6(Ser235/236) (D57.2.2E) XP® Rabbit mAb and H-89 (Dihydrochloride) were purchased from Cell Signaling Technology (Beverly, MA, USA). Phospho-AKT (Thr308) antibody, β-tubulin antibody and β-actin antibody were purchased from Bioss (Bioss Beijing, China). KN-62 was purchased from TOCRIS (Minneapolis, MN, USA). Mandarin fish S. chuatsi (3 months of age, total length 21.22 ± 1.35 cm) were obtained from the Chinese Perch Research Center of Huazhong Agricultural University (Wuhan, China). Fish were maintained in an aquarium (60 × 45 × 45 cm) with a continuous system of water filtration and aeration at constant temperature (25 ± 1 °C). Fish were acclimated to the condition for 2 weeks prior to experimentation. Fish were fed once daily at 5:30 pm with live India mrigal Cirrhinus mrigala juvenile. The animal protocol was approved by The Scientific Ethics Committee of Huazhong Agricultural University (Wuhan, China). The ethical code is HZAUFI-2019-013.
4.2. Learning Test
The experimental fish were randomly divided into two groups (
n = 12)—the control group and the learning group. The control group: Experimental fish were paired with the negative demonstration fish (fish without training). The learning group: Experimental fish were paired with the positive demonstration fish (fish had been pre-trained to accept dead prey fish using the method mentioned in the previous study [
15]). The live and frozen India mrigal were used as live prey fish and dead prey fish, respectively. During the test period, mandarin fish were fed with prey fish 10 times (2 min per time) at 5:30 pm each day over a 12 day period. The prey fish were placed 20 cm away from mandarin fish, and only one live or dead prey fish was allowed into each aquarium at any time. After 2 min, the live or dead prey fish that had not been eaten were removed from the aquarium. The testing process was recorded with a digital camera. The success rates, learning times of feeding dead prey fish, and food intake were counted from the videos. As long as one dead prey fish was successfully accepted, the experimental fish was marked as a successful transformer of feeding habits to dead prey fish. No mandarin fish died of natural causes during the testing.
4.3. Sample Collection
The experimental fish (n = 12) were anesthetized with MS-222 (Argent Chemical Laboratories, Redmond, WA, USA) (200 mg L−1), and euthanized by decapitation at 2 h after feeding. Brain tissues were immediately frozen in liquid nitrogen upon surgical resection and stored at −80 °C prior to RNA isolation and protein isolation.
4.4. RNA Isolation and Reverse Transcription
Total RNA was isolated with a Trizol reagent (TaKaRa, Tokyo, Japan) following the manufacturer’s protocol. The extracted RNA was re-suspended in 30 μL of RNase-free water and then quantified with a BioTek Synergy™2 Multi-detection Microplate Reader (BioTek Instruments, Winooski, VT, USA) and agarose gel electrophoresis. One microgram of total RNA was used for reverse transcription with Revert Aid™ Reverse Transcriptase (TaKaRa, Tokyo, Japan) according to the manufacturer’s instructions. The synthesized cDNA was stored at −20 °C.
4.5. Real-Time Quantitative PCR
Primers were designed using Primer 5.0 software (Premier, Canada) based on the sequences obtained from transcriptome sequencing data of mandarin fish, and they were synthesized by Sangon (Shanghai, China). The amplification information of the primers is listed in
Table 2. A set of housekeeping genes including β-actin,
rpl13a,
b2m,
ywha2,
hmbs and
sdha were selected according to the literature [
48]. The
Rpl13a gene was more stable and amplified as the internal control. Real-time quantitative PCR was carried out with a MyiQ™ 2 Two-Color Real-Time PCR Detection System (Bio-Rad, Hercules, CA USA). PCR was performed in a total volume of 20 μL containing 10 μL of AceQ® qPCR SYBR® Green Master Mix (Vazyme, Piscataway, NJ, USA), 0.4 M of each primer, and 20 ng of cDNA. The PCR cycling parameters were 95 °C for 5 min, followed by 40 cycles of 95 °C for 10 s, and annealing temperatures for 30 s. A melt curve analysis was performed from 65 to 95 °C, gradually increasing at 0.5 °C/6 s, to verify the specificity. Reactions for each sample were performed in triplicate. Gene expression levels were quantified relative to the expression of
rpl13a using the optimized comparative Ct (2
−ΔΔCt) value method [
49]. Data are presented as mean ± S.E.M. (
n = 6).
4.6. Transcriptome Sequencing
An equal amount of total RNA of each group was used to construct the libraries for the transcriptome analysis using MGIEasy RNA kit following manufacturer’s instructions (BGI, Wuhan, China). Poly(A) mRNA was purified from total RNA using oligo-dT-attached magnetic beads. Paired-end cDNA libraries were sequenced using the BGISEQ-500 system (BGI, Wuhan, China). Image deconvolution and base calling were performed with the SOAPnuke. Clean reads were obtained by removing adaptor reads and low quality reads (Q ≤ 10), on which all following analyses were based. Transcriptome assembly was carried out with short reads assembling program Trinity with k-mer length 25 bp. The reads were mapped back to assembled contigs. By using the paired-end information, it was able to detect contigs from the same transcript as well as the distances between these contigs. We connected the contigs using N to represent unknown sequences between each pair of contigs, and then scaffolds were made. Paired-end reads were used again for gap filling of scaffolds to obtain sequences with least Ns and could not be extended on either end. Such sequences were defined as unigenes. To annotate the transcriptome, we performed the BLAST alignment between unigene and databases such as the Kyoto Encyclopedia of Genes and Genomes (KEGG), Gene Ontology (GO), NR, NT, SwissProt, Pfam and KOG with Blast2GO, hmmscan and getorf software.
To estimate expression levels, the RNA-Seq reads generated were mapped to the unigenes using Bowtie2. Gene expression levels were measured through RSEM. We analyzed the differentially expressed genes used DEGseq method described before [
50], and false discovery rate (FDR) ≤ 0.001 and fold change ≥ 2.00 as the threshold to judge the significance of gene expression difference. A GO functional analysis and a KEGG pathway analysis were then carried out in differentially expressed genes.
4.7. Signaling Pathways Analysis
Brain tissues stored at −80 °C were solubilized in a lysis buffer, and lysates were separated on 10% SDS-PAGE gel. Proteins were then transferred onto a PVDF membrane. Phospho-cAMP-response element-binding protein (p-CREB), Phospho-protein kinase A (p-PKA), Phospho- extracellular regulated protein kinases (p-ERK1/2), Phospho-protein kinase B (p-AKT), Phospho-Ca2+/calmodulin-dependent protein kinase II (p-CaMKII), Phospho-S6 ribosomal protein (p-S6), β-tubulin and β-actin were detected by immunoblotting with the antibody (1:1000–1:4000). Blots were probed by goat anti-rabbit or goat anti-mouse second antibody with IR-Dye 680 or 800 cw labeled (1:2000–1:4000, Licor, Lincoln, NE, USA) at room temperature for 1 h. The membranes were then visualized using a LiCor Odyssey scanner (Licor, Lincoln, NE, USA) and quantified with ImageJ 1.44 software (National Institute of Health, Bethesda, MD). The phosphorylation levels of the cAMP-response element-binding protein (CREB), the cAMP-dependent protein kinase (PKA), the extracellular regulated protein kinases (ERK1/2), protein kinase B (AKT), Ca2+/calmodulin-dependent protein kinase II (CaMKII) and S6 ribosomal protein (S6) were normalized according to the loading of proteins by expressing the data as a ratio over β-actin or β-tubulin.
4.8. Effects of the Inhibitors for PKA and CaMKII Signaling Pathway
Brain cells were obtained from mandarin fish and maintained at 28 °C in L15 (leibovitz’s L15-medium) containing 20% newborn calf serum and 5% bio-antibiotics (Penicillin-Streptomycin). After three generations, cells were plated on 60 mm dishes and incubated at 28 °C without CO2. Four days later, the cells were with 80% coverage, were washed twice, and starved for 24 h with L15 media. The starved cells were treated for 0.5–3.0 h with either buffer alone or different concentration of the inhibitors (H-89 for the PKA signaling pathway, KN-62 for the CaMKII signaling pathway). The best time point and drug concentration were chosen based on pre-experiment. Total RNA or protein samples were collected with a lysis buffer or Trizol reagent. The gene expression levels of learning or appetite control related genes and the phospho-PKA, phospho-CaMKII and phospho-CREB were detected by real-time quantitative PCR and a Western blotting analysis, respectively.
4.9. ChIP Assay
A CHIP assay was performed using a SimpleChIP® Enzymatic ChIP kit (CST, USA) according to the manufacturer’s instruction. The brain tissue from KM mice was isolated, and the DNA sample was purified with a ChIP Kit following manufacturer’s instructions. Positive controls were 10% of each DNA sample, which did not include the immunoprecipitation step. The remainder of the samples were equally divided into two groups. The experimental group was immunoprecipitated with a specific mouse c-Fos antibody (Santa Cruz, USA; 1:500), whereas the negative control group was immunoprecipitated with a general mouse IgG antibody (proteintech, wuhan; 1:250). The isolated DNA was firstly purified then amplified by PCR using specific primers encompassing the AP-1 binding site of the anorexigenic neuropeptides
proopiomelanocortin (
pomc) gene. The putative transcription factor binding sites (TFBS) of the regulatory region at 2000 bp upstream from the transcriptional start site of the
pomc gene were predicted by AliBaba2.1 software (
http://gene-regulation.com/pub/programs/alibaba2) and PROMO using version 8.3 of TRANSFAC (
http://alggen.lsi.upc.es/cgi-bin/promo_v3/promo/promoinit.cgidirDB=TF_8.3). These putative binding sites of c-Fos protein are located at -1541 to -1534 bp (gtttcagt, site 1), -1128 to -1119 bp (tcactgaagg, site 2), and -383 to -374 bp (tttatagtga, site 3) on the upstream of the transcriptional start site. The primers are 5′-GCCAACCCAAACTTACCT-3′ (site 1, forward primer) and 5′-TTCAACCTCCCATCCACC-3′ (site 1, reverse primer), 5′-AACTGGGAGATGATGGGG-3′ (site 2, forward primer) and 5′-CGGAGTGACTTCCTGCTGT-3′ (site 2, reverse primer), 5′-CCCACTCCAAAAGGTAGC-3′ (site 3, forward primer) and 5′-TGAGGAAGGGGATTTGTTA-3′ (site 3, reverse primer).
4.10. Statistical Analysis
Statistical analyses were performed with SPSS 19.0 software (SPSS, Chicago, IL, USA). All data were tested for normality and homogeneity of variances using the Shapiro–Wilk’s test and Levene’s test, respectively. Significant differences were found using a one-way analysis of variance (ANOVA), followed by Fisher’s least significant difference post hoc test and Duncan’s multiple range tests, after confirming data normality and homogeneity of variances. Differences were considered to be significant if p < 0.05.

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}