Abstract
Ceramide and sphingosine display a unique profile during brain development, indicating their critical role in myelinogenesis. Employing advanced technology such as gas chromatography–mass spectrometry, high performance liquid chromatography, and immunocytochemistry, along with cell culture and molecular biology, we have found an accumulation of sphingosine in brain tissues of patients with multiple sclerosis (MS) and in the spinal cord of rats induced with experimental autoimmune encephalomyelitis. The elevated sphingosine leads to oligodendrocyte death and fosters demyelination. Ceramide elevation by serine palmitoyltransferse (SPT) activation was the primary source of the sphingosine elevation as myriocin, an inhibitor of SPT, prevented sphingosine elevation and protected oligodendrocytes. Supporting this view, fingolimod, a drug used for MS therapy, reduced ceramide generation, thus offering partial protection to oligodendrocytes. Sphingolipid synthesis and degradation in normal development is regulated by a series of microRNAs (miRNAs), and hence, accumulation of sphingosine in MS may be prevented by employing miRNA technology. This review will discuss the current knowledge of ceramide and sphingosine metabolism (synthesis and breakdown), and how their biosynthesis can be regulated by miRNA, which can be used as a therapeutic approach for MS.
1. Introduction
Sphingolipids are amphipathic molecules with diverse functions in cell development and death. Ceramide (Cer) and sphingosine (Sph) are two unique sphingolipids that comprise the core structural bases for all sphingolipids. The biological functions of sphingoids such as Sph, dihydrosphingosine (dhSph), and psychosine as well as of sphingolipids are regulated by their relative concentrations in cells and tissues. For example, Cer participates in a wide variety of biochemical events such as protein phosphorylation, modulation of protein kinase C (PKC) and phospholipase A2, but mediates signal transduction leading to cell death at a high concentration, while Sph and psychosine are potent inhibitors of PKC [1]. Hence, the precise concentration of sphingolipid metabolites is the key to normal tissue development and maintenance. Besides their important regulatory functions in brain development and myelinogenesis [1], they also actively contribute to cell death, including promotion of cell death in oligodendrocytes in multiple sclerosis (MS) [2].
MS is a devastating demyelinating human disease leading to extensive neurodegeneration and is characterized by immune mediated loss of myelin and axons [3]. The clinical manifestations and disease progression in MS are substantially variable and related to complex genetic, epigenetic, and environmental factors [4,5,6,7]. MS is generally characterized by the infiltration of lymphocytes into the central nervous system (CNS), leading to axonal demyelination and degeneration of neurons [8]. The development of modern genomics has successfully identified the various molecular mechanisms in MS pathogenesis [9]. The down- and up-regulation of specific microRNAs (miRNAs) may nowadays be used as biomarkers for predicting disease progression [10]. Identifying agents inhibiting the enzymes controlling epigenetic modifications, particularly DNA methyltransferases and histone deacetylases, may be introduced as promising tools for therapeutic interventions in MS [11].
Although multiple factors may have been implicated in the pathogenesis of MS, all reports indicate a severe change in the biochemical milieu in the CNS in progression of MS. The inflammatory microenvironment contains a variety of substances such as proteolytic enzymes, cytokines, oxidative products, and free radicals that can injure neurons and axons [12]. Studies including human and animal models of MS have indicated that inflammatory cytokines play a critical role in MS pathogenesis [13] by fostering inflammation and demyelination [10]. Cytokines such as tumor necrosis factor alpha (TNFα) and interferon gamma (IFNγ) mediate aberrant lipid metabolism with accumulation of toxic Sph in MS brains [2,14]. This, in turn, may further damage brain function due to degeneration of oligodendrocytes and neurons, the two key CNS cell types for myelin constituents [2]. Investigations have shown that an intermittent increase in Cer by TNFα and IFNγ stimulation of serine palmitoyltransferase (SPT) activity followed by Sph accumulation in spinal cords from Lewis rats with experimental autoimmune encephalitis (EAE) resulted in induction of apoptosis in the lumbar spinal cord [2]. TNFα and IFNγ stimulated Cer elevation in cultured human oligodendrocytes and Cer production was blocked by myriocin, an inhibitor of SPT, causing prevention of apoptosis in oligodendrocytes [2].
Medications are used in MS to modify the disease course and manage MS symptoms. Although there is no cure, the medications are apparently essential components that help people manage their MS and enhance their life. Such medications include oral administration of teriflunomide, fingolimod, dimethyl fumarate, injectable administrations (interferon beta 1a, glatirmer acetate, and daclizumab), and infused medications (alemtuzumab, mitoxantrone, and natalizumab) [15]. Symptomatic medications are also available to treat syndromes such as bladder problems, fatigue, emotional stress, etc. Although many clinical trials are being conducted for MS, to the best of our knowledge, there is no primarily sphingolipid-targeted therapy available, with the exception of fingolimod, which acts as a ligand for the Sph-1-P receptor and inhibits Cer synthase [2]. Hence, Sph toxicity via Cer generation may be a potential target in both EAE and MS as an alternate therapy by blocking Cer synthesis.
In this article, we introduce the concept of miRNA therapy to protect the oligodendrocytes from apoptosis in order to prevent demyelination in MS. Expression of several miRNAs such as miR-155 and miR-326 are increased in MS brains and these may have potential roles in MS pathogenesis [3]. Several factors that contribute to MS pathogenesis foster aberrant lipid metabolism leading to accumulation of Sph, a toxic sphingolipid, that has been demonstrated to mediate apoptosis in human gastric cancer cells [16], hepatoma cells [17], and rhabdomyosarcoma cells [18]. Our recent study demonstrated an up-regulation of Sph in MS tissues causing demyelination via oligodendrocyte apoptosis [2].
There are many therapeutic strategies available for MS based on the patient’s condition. Although fingolimod is a lipid inhibitor, its efficacy in inhibiting Cer generation is much less than that of myriocin [2]. Myriocin itself displays toxicity in humans and hence is unsuitable for human therapy. Other natural or synthetic inhibitors of Cer synthesis such as fumonisin B1 may display severe adverse effects in humans [19]. Hence, an alternate therapy that involves the use of miRNA may show high potential. At present multiple effects of miRNAs are well known, and they are considered to be nontoxic (or less toxic) because of their natural synthesis in humans. This review describes the basic rationale of miRNA therapy to regulate Cer/Sph accumulation in MS as a novel therapeutic approach for MS.
2. Diversity in Sphingolipid Functions
2.1. Ceramide (Cer) and Sphingosine (Sph) Stimulate Myelinogenesis
To determine the regulation of sphingolipids in myelinogenesis, we quantified the concentrations of Cer, Sph/dhSph, and monoglycosylceramides (MGCs) during rat brain development using our advanced methodology [20] and showed that all three components are critical for myelination. An increasing concentration of Cer during development, with an optimum concentration at postnatal day (P) 21, clearly signifies its direct involvement during myelination. However, its precise mechanism (i.e., interactions with other myelin components) has not yet been explored. Cer, being the core structure, may stimulate the synthesis of other myelin-specific sphingolipids such as GalCer, GM1, etc. that are pertinent to the interactive composition of myelin. Interestingly, the Cer/dihydro-(dh) Cer ratio determined using gas chromatography–mass spectrometry (GC–MS) and high performance liquid chromatography (HPLC) indicated a constant value between 4.0/1 and 4.5/1 during normal brain development. A higher ratio has been observed in MS brains where an aberrant sphingolipid metabolic activity has recently been confirmed [2].
For MGC, GlcCer is the only component in embryonic and early postnatal brains (P2 to P5). GalCer synthesis initiates during subsequent developmental stages (P8–P10) with increasing concentration, reaches an optimum at P21, and then stabilizes. The activities of two synthesis enzymes, Cer:glucosyltransferase and Cer:galactosyltransferase were in correspondence with the GlcCer and GalCer concentrations [21]. The GalCer concentration in MS brain tissue has been greatly reduced as a direct manifestation of extensive demyelination [22]. A schematic presentation of sphingolipid-mediated myelinogenesis is shown (Figure 1).
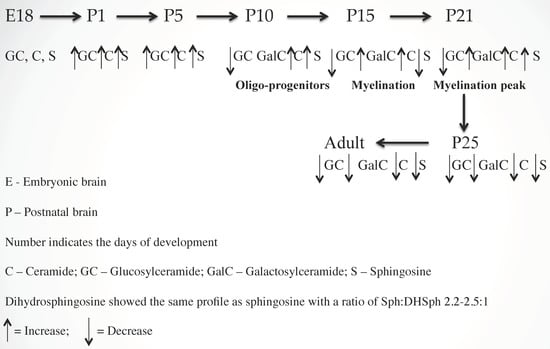
Figure 1.
Sphingolipid regulated vertebrate brain development and myelinogenesis.
Similarly, Sph/dhSph concentrations are gradually elevated until P10 during the stage of (oligodendrocyte) development and then they decrease. A secondary short peak is seen during myelinogenesis at P21 with a steady ratio (2.2–2.5/1) of Sph/dhSph, which varies greatly in MS brains [1]. Cer is produced from dhCer (by desaturase or DES), which in turn is hydrolyzed to Sph by ceramidase (Figure 2). The accumulation of Sph may be triggered by excess Cer biosynthesis (as a breakdown product of ceramidase) as shown in the diagram (Figure 2). Sph plays an unknown but critical role in oligodendrocyte synthesis and maturation during myelination by activating the melastatin-like transient receptor potential protein 3 (TRMP3) [23] which facilitates spontaneous Ca2+ entry. Hence, the appearance of the peak at P10 and then a short peak at P21 [1] correlates with the activation of TRMP3, as TRPM3 participates as a Ca2+-permeable and Sph-activated channel in oligodendrocyte differentiation (P10) and CNS myelination [23]. This corresponds with our observation that a lower Sph concentration stimulates human oligodendrocyte proliferation, while a higher concentration leads to cell death [2,24]. However, the detailed mechanism by which Cer and Sph aid myelinogenesis needs further investigation.
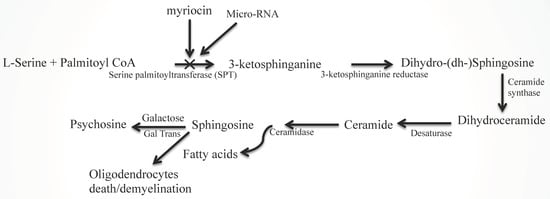
Figure 2.
Ceramide and sphingosine generation via SPT activation and its inhibition by myriocin and miRNA. The biosynthesis of sphingolipids (dihydrosphingosine, ceramides, and sphingosine) in the de novo pathway begins with the condensation of palmitoyl-CoA and L-serine by serine palmitoyltransferase (SPT), producing 3-ketosphinganine or 3-ketodihydrosphingosine. The rate-limiting enzyme of the sphingolipid biosynthesis pathway is SPT, which can be pharmacologically inhibited by myriocin. Then, 3-ketosphinganine reductase (3-ketodihydrosphingosine reductase) reduces 3-ketosphinganine to sphinganine (dihydrosphingosine). Ceramide synthase or fatty acyltransferase converts sphinganine (dihydrosphingosine) to dihydroceramide. Dihydroceramide is dehydrogenated (desaturated) at C3–C4 of the sphinganine of dihydroceramide by desaturase to generate ceramide. The action of ceramidase converts ceramide into sphingosine via releasing fatty acids. The enzyme galactosyltransferase catalyzes the reaction between sphingosine and UDP-galactose to produce psychosine (galactosylsphingosine).
We have recently described the chromatographic purification and characterization of phytoCer in vertebrate brains and other tissues [25], indicating that a more careful evaluation of Cer biosynthesis is warranted as no such naturally-occurring Cer species has been reported in vertebrates. We postulate that phytoCer may be biosynthesized by addition of a fatty acyl group to phytoSph by a phytoCer synthase, and this enzyme has not yet been reported [1]. Moreover, a huge accumulation of Sph at an initial developmental stage (P10) without any major peaks for Cer leads us to predict that a DES may possibly exist that directly converts dhSph into Sph [1].
2.2. Sphingolipid Metabolic Disorders
Sphingolipid metabolic diseases are designated as inborn errors of metabolism, which include Gauscher’s and Krabbe’s disease [26,27,28], GM1 gangliosidosis [29], Tay-sach’s disease [30], etc. In these diseases, there is an accumulation of a specific sphingolipid in a particular tissue(s)/organ(s) due to the deficiency of a specific metabolic enzyme. However, no such disease correlating Cer or Sph accumulation has yet been reported. Cer has been at the center of extensive study for its role in various cell death mechanisms in nervous system disorders [31,32,33,34]. We have recently reported Sph elevation preceeded by a transient accumulation of Cer in MS brains [2]. This mechanism is due to cytokine stimulated SPT activation, which is an initial step of the pathway leading to Cer biosynthesis (Figure 2), rather than sphingomyelin degradation (salvage pathway). Although Sph has been shown to trigger oligodendrocyte cell death, the precise mechanism remains uninvestigated.
2.3. Aberrant Lipid Metabolism in MS Brains
Demyelination in MS may proceed via different mechanisms in different MS patients due to its stimulation by a variety of factors such as proteolytic enzymes, cytokines, oxidative products, and free radicals [18]. It is evident that inflammatory cytokines play a critical role in pathogenesis in MS lesions in humans and animals [35,36] by triggering inflammation [12,36], although the mechanism of cytokine action remains unknown. Immune cell filtration accompanying chronic inflammation signifies that MS is an autoimmune demyelinating disease, even though the precise etiology of MS has not yet been resolved [37]. Moreover, many major issues such as the primary cause of inflammation, primary target antigen, stimulation of autoimmunity, etc. have not yet been answered [38,39,40].
The CNS, more specifically, myelin, is enriched with sphingolipids and lysosphingolipids such as GalCer and Cer, sphingoids, and psychosine [20]. An increasing concentration of these sphingolipids during brain development and myelinogenesis implies that (lyso)sphingolipids may participate in cell growth, differentiation, myelinogenesis, and maintenance of the structural and functional integrity of myelin [1,20]. However, their concentration in white matter of a normal brain, in normal appearing white matter (NAWM) and in a plaque of a MS brain significantly varies [2]. An accumulation of psychosine and increasing trend of Sph has been reported in NAWM, while MS plaque shows a decrease in Cer concentration but an accumulation of Sph. The breakdown of Cer by ceramidase is the prime source of Sph production, while cytokine-mediated activation of SPT plays the major role in Cer synthesis in vivo (Figure 2). Although there are few studies involving the sphingolipid/lipid metabolism in MS [41,42,43], our recent study [2] has shown the detailed changes in sphingolipid profile with a dramatic accumulation of Sph in patient brains and EAE animal spinal cords, and this is in agreement with a recent publication [44].
Briefly, we have shown that in addition to Galcer, both Cer and Sph are necessary components for myelinogenesis, maintaining a defined ratio of Cer/dhCer and Sph/dhSph during the rat brain development and a higher ratio is observed in MS due to accumulation of Cer and Sph that lead to oligodendrocyte death and thereby fostering demyelination.
2.4. Expression of miRNAs
The small non-coding RNAs, which have an average length of 22 nucleotides, are known as miRNAs. All miRNAs are evolutionarily conserved and have been detected only in eukaryotic cells. The first miRNA was discovered in Caenorhabditis elegans. It showed complementarity to the 3’ untranslated region (3’UTR) of a target messenger RNA (mRNA) for a RNA–RNA interaction and its degradation (silencing) [45]. This discovery triggered many other studies, which concluded that all miRNAs were single stranded RNA molecules (ssRNA). They have a stem-loop structure. All miRNAs are highly stable and are partially complementary to the 3’UTR of the target mRNAs. Moreover, all miRNAs can act on multiple target mRNAs. Studies have shown that each miRNA may degrade almost 100 different target mRNAs. All miRNAs form a network of complex regulation. A specific mRNA may contain multiple binding sites for many miRNAs. Intergenic regions and introns of protein-coding genes most frequently contain miRNA genes. It now appears that so called ‘junk DNA’ locations act as the origin of most of the miRNAs. Studies have revealed that biogenesis and maturation of miRNAs occur in four steps: transcription, nuclear processing, nuclear exportation, and cytoplasmic processing [45]. It is now well-established that all miRNAs are highly conserved molecules and play highly regulatory roles in activation or inhibition of many cell-signalling pathways during normal development. Deregulation of miRNAs causes diseases.
2.4.1. miRNAs in Oligodendrocyte Development and Ceramide Metabolism
Although significant progress has been made in examining lipid metabolism, little effort is made to investigate the role of miRNAs in sphingolipid metabolism [46]. Oligodendrocyte (oligo) progenitors express A2B5 antigen, while GalC is the considered as the matured oligo marker. The miRNA expression profile in oligo lineage cells is known as “microRNAome” that contains 43 miRNAs [47] and their expression changes from oligo progenitor cells (A2B5) to pre-myelinating oligo (GalC+) cells that include a target bias for a class of miRNA (including miR-9). MiR-9 expression is down-regulated during differentiation of oligos and its expression inversely correlates with the expression of its target which is the peripheral myelin protein 22 (PMP22) [47]. By interacting with the PMP22, miR-9 down-regulates the expression of PMP22 [47]. SPT is the first limiting enzyme of de novo Cer synthesis (Figure 2) and its precise regulation is still obscure. A recent study examining the regulation of SPT in Alzheimer’s disease identified the loss of miR-137, miR-181c, miR-9, and miR-29a/b-1, which stimulated SPT and Aβ levels [48]. Moreover, FTY720 increases miR-376, miR-30, miR-128, miR-126, miR-7, and miR-9, interfering with Cer synthesis [49]. Hence, SPT down-regulation by miRNA technology may be considered as a novel approach to protect the oligos from Cer/Sph toxicity.
2.4.2. miRNAs in MS and Neurodegenerative Disorders
Current studies strongly indicate alterations in expression of neurodegenerative and neuroprotective miRNAs in neurodegenerative injuries or diseases including MS [50,51,52]. Levels of neurodegenerative miRNAs are increased, while levels of neuroprotective miRNAs are down-regulated, contributing to progressive neurodegeneration. Fumonisin B1, a naturally occurring myotoxin in maize and an inhibitor of Cer synthesis significantly down-regulated expression of miR-27 in HepG2 cells, while human cytochrome P450 mRNA and protein expression were up-regulated in HepG2 cells [53]. Therefore, levels of specific miRNAs can serve as important biomarkers for diagnosis, treatment, and prognosis in a CNS injury or disease [54]. Recently, a radical miRNA-based therapy has been designed to encompass neuroprotection and neuronal restoration [55,56].
2.4.3. Role of miRNA in Normal and Pathological Functions of Rodent and Human CNS
EAE is a rodent model that can reproduce typical MS lesions that occur due to inflammation, demyelination, and axonal damage [57]. While MS is generally characterized by loss of myelin and axons leading to progressive neurological deterioration [3,58], there are no specific tests for MS, its diagnosis often relies on ruling out other conditions that might produce similar signs and symptoms, known as a differential diagnosis. These include blood tests, magnetic resonance imaging (MRI), lumber puncture, and evoked potential tests such as visual stimuli, or electrical stimuli [59].
Numerous studies, aiming for a diagnosis of MS by examining the miRNA expressions, have detected specific miRNA ‘signatures’ for MS. For example, in peripheral blood mononuclear cells, a distinct profile of miRNA has been identified between relapse and remission states in MS and EAE, a rodent model for MS [57,60,61]. Accumulating evidence shows that the miRNAs in the CNS may play crucial roles in abnormal and normal functions in CNS. For example, overexpression of miR-134 is associated with epilepsy in experimental rodent models and temporal lobe epilepsy in humans [60]. Inhibition of miR-134 expression in the brain using antisense oligonucleotides can attenuate status epilepticus in rodents [58]. Many miRNAs are involved in immune system development and also regulate the immune system; for example, miR-150 is critical for B cell differentiation [61], miR-155 promotes autoimmune inflammation [62], and miR-146a regulates the T-cell mediated T helper-1 (Th-1) response [63] and is expressed in interleukin-17 (IL-17)-producing Th-17 cells in rheumatoid arthritis [64]. IL-17-producing Th-17 cells plays a key role in MS and other autoimmune diseases [64,65,66,67] and a recent study showed a correlation between miR-326 expression and the disease severity in MS patients as well as in EAE rats [68]. All continuing efforts to search for advanced detection technique(s) in identifying the early onset of MS may lead to a prompt and efficient therapy.
3. MS Therapy with Conventional, Complementary, and Alternative Medicines and the Perspective of miRNA Targeting
Treatment for MS typically focuses on gaining some recovery after attacks, slowing the progression of the disease, and managing MS symptoms. Conventional therapy varies from drugs to diet. Corticosteroids and plasmapheresis, used as a common treatment for MS, have been found through clinical trials to reduce the number of relapses and inflammation, and limit new disease with a short span of activity. Ocrelizumab (infusion administration) has been administered to reduce the number of relapses, delay progression of disability (in some patients), and limit new disease activity (seen on MRI). Other medications such as beta interferons, fingolimod, dimethyl fumerate and other drugs are used to prevent or slow down the MS progression but they have minimal effects in delaying disability progression [15]. Symptomatic medications such as muscle relaxants, fatigue medication, and antidepressants are also used to treat the specific symptoms. To maintain body fitness, typical alternative treatment strategies such as Tai Chi, chiropractor therapy, meditations, and yoga are also recommended for relieving stress [15]. A list of the therapeutic drugs and other alternative therapies for MS patients is provided in Table 1.

Table 1.
A short list of common medications and other therapies for MS patients*.
Although some miRNAs have been shown to promote inflammation and autoimmunity via Th cells, their precise roles in regulation of the inflammatory diseases have not yet been clearly defined. Recently, they have been used as a new therapeutic approach in the treatment and prevention of autoimmune disorders. Alteration in specific miRNA expression may produce desirable therapeutic outcomes in MS and other autoimmune diseases (Table 2). A therapeutic strategy targeting miRNA in MS and EAE relies on manipulation of endogenous miRNA levels by delivering the oligonucleotides mimicking or inhibiting the specific mRNA sequences [67]. MS and EAE are associated with overexpression of miR-155, knockdown of which results in low Th1 and Th17 cells and mild EAE [67,68]. MiR-155-3p up-regulates the Th17 population by inhibiting two heat shock protein 40 genes and this contributes to the development of EAE [69]. As miRNAs target multiple regulatory gene expressions, they may have an advantage over the concept of targeting multiple genes individually. At the same time, potential target of multiple genes is also a concern. However, these side effects could be modified and addressed by utilizing the effective methods of delivery. The miRNA therapeutic approach technology has recently been examined in different cancer models with minimum or no toxicity [70,71]. Obliteration of miR-338 aggravates the miR-219 mutant hypomyelination phenotype but miR-219 mimics enhance myelin restoration in EAE animals, indicating a therapeutic role for miR-219 in myelin repair [72].
Table 2.
Emerging therapeutic strategies for alteration in levels of specific miRNAs in the treatment of MS and EAE.
The miRNA-mediated regulation of sphingolipid metabolism is of current scientific interest. Down-regulation of miR-101 leading to Cer synthase elevation and upregulation of Cer synthesis has been reported in metastasis-prone lung cancer cells [74]. A change in the sphingolipid compositions, along with a sub-acute change in plasma miRNA, in traumatic brain injury has also been reported [75]. A remarkable conservation of the majority of miRNA and the total lipid content indicates that they can potentially be used as biomarkers for diagnosis of diseases [76]. Despite all the recent advanced knowledge about immunopathogenesis and demyelination in MS, there is no efficacious therapy available for this devastating disease. Because our study indicates an elevation of ceramide via SPT activation leading to ceramide/sphingosine accumulation in MS [2], we propose that a therapy to block the SPT1 gene using miRNA technology will prevent the ceramide generation and this may be considered as an alternate therapeutic approach. Fingolimod (FTY720) is phosphorylated in vivo to form fingolimod-phosphate. Fingolimod-phosphate initially activates lymphocyte S1P1 via high-affinity receptor binding, yet subsequently induces S1P1 down regulation so as to prevent lymphocyte egress from lymphoid tissues, thereby reducing auto-aggressive lymphocyte infiltration into the CNS and further preventing demyelination in MS [73]. Circulating miR-15b, miR-23a and miR-223 levels were significantly down regulated in serum samples from relapsing remitting MS patients, but levels of these circulating miRNAs recovered in the MS patients following fingolimod treatment for 6 months, indicating that pharmacological manipulation of levels of miRNAs could be an emerging therapeutic strategy in reducing the frequency of exacerbations in MS patients [77]. However, our study indicates that fingolimod has much less potential in blocking Cer elevation compared to myriocin [2]. These results are important as they indicate that targeting Cer biosynthetic pathways for alteration in levels of specific miRNAs may provide significant therapeutic benefits in relapsing remitting MS patients. A detailed overview of the experimental design and findings of studies investigating miRNAs as potential biomarkers of MS have recently been published [78], which may help in designing efficient miRNAs in future MS therapy. An approach of MS therapy can be initiated by targeting SPT regulation by selecting one or more from a group of miRNAs such as miR-376, miR-30, miR-128, miR-126, miR-7, and miR-9, as these miRNAs are involved in interfering with Cer synthesis [49].
4. Conclusions
In summary, we have explored the dual role of Cer and Sph in mammalian brain development and diseases, specifically in MS. It is worth mentioning that Cer is a major component of sphingolipid (g/g) while Sph is a minor component (ng/g), and they play a significant role in myelinogenesis. Their roles in apoptosis in MS leading to demyelination has recently been published [2]. Sph is the base structure of a sphingolipid and displays toxicity with a minor change in concentration. Measuring the Sph/dhSph ratio in the blood of a MS patient may be utilized to predict the onset of MS, but this needs a careful and thorough investigation.
MS pathology and demyelination are still an open field for research exploring the precise cause of MS. The diagnosis of MS includes multiple procedures such as MRI, spinal fluid examination (for abnormal immune response), vision and memory assessments, nerve conduction, etc., all of which confirm the onset and progression of MS. There is a great demand for evaluating an early assessment and diagnosis of MS in order to initiate therapy at a preliminary stage of the disease. It is noteworthy that there is no single therapeutic approach that can help control and/or improve the patient’s health and, of course, there is no comprehension for cure. Most therapies are based on the patient’s symptoms and include a wide variety of medications and conventional therapies. One such difficult hurdle is our lack of knowledge regarding the precise biochemical mechanisms, the complex interactions and regulation, of oligodendrocytes, neurons, and astrocytes during myelin formation, and the participation of biomolecules such as lipids and proteins in protecting the myelin sheath. It is now certain that there are dramatic changes in the sphingolipid profile in MS brains and such changes have also been reflected in NAWM as a clear indication of the disease progression. These changes are mediated by primary biochemical alterations and/or a series of cytokines that are auto-released, either offering immunoprotection against some unidentified invasive agents that are yet to be characterized. Although current therapeutic strategies are unable to offer an absolute cure, they can help patients towards a better life by protecting the oligodendrocytes/neurons from lipid-mediated cell death. One such option includes treatment with synthetic enzyme inhibitors, but most, if not all of them, may be toxic to our system and may exert serious side effects. Alternatively, we can use miRNAs to inhibit enzyme-mRNA expression with lesser side effects using innovative technology for drug delivery. With other conventional therapeutic methods, one must develop sphingolipid miRNA technology not only for MS but also for other diseases (such as cancers) where bioactive lipids also play a devastating role.
Acknowledgments
This work was supported in part by these grants: NINDS-NS-31355 (SD), NIAAA-11865 (SD), SC State Appropriation # CR22 (SD), NCI-CA-91460 (SKR), NINDS-NS-057811 (SKR), and SC SCIRF-2015-I-0 (SKR).
Conflicts of Interest
The authors have no conflict of interests to declare.
References
- Dasgupta, S.; Ray, S.K. Diverse biological functions of sphingolipids in the CNS: Ceramide and sphingosine regulate myelination in developing brain but stimulate demyelination during pathogenesis of multiple sclerosis. J. Neurol. Psycol. 2017, 5, 1–7. [Google Scholar]
- Miller, L.G., Jr.; Young, J.A.; Ray, S.K.; Wang, G.; Purohit, S.; Banik, N.L. Dasgupta S. Sphingosine toxicity in EAE and MS: Evidence for ceramide generation via serine-palmitoyltransferase activation. Neurochem. Res. 2017, 42, 2755–2768. [Google Scholar] [CrossRef] [PubMed]
- Kücükali, C.I.; Kürtüncii, M.; Çoban, M.; Tüzün, E. Epigenetics of multiple sclerosis: An updated review. Neuromol. Med. 2015, 17, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Urdinguino, R.G.; Sanchez-Mut, J.V.; Esteller, M. Epigenetic mechanisms in neurological diseases; genes, syndrome, and therapies. Lancet. Neurol. 2009, 8, 1056–1072. [Google Scholar] [CrossRef]
- Lauer, K. Environmental risk factorsin multiple sclerosis. Expert. Rev. Neurthera. 2010, 10, 421–440. [Google Scholar] [CrossRef]
- Oskenburg, J.R.; Baranzini, S.E. Multiple sclerosis genetics-Is the glass half full or half empty? Nat. Rev. Neurol. 2010, 6, 429–437. [Google Scholar]
- Hunter, S.F. Overview and diagnosis of multiple sclerosis. Am. J. Manag. Care 2016, 22, s141–s150. [Google Scholar]
- Compston, A.; Cole, A. Multiple sclerosis. Lancet 2008, 372, 1502–1517. [Google Scholar] [CrossRef]
- Lindberg, R.L.; Hoffman, F.; Mehling, M.; Kuhle, J.; Kappos, L. Altered expression of miR17-p in CD+ lymphocytes of relapsing-remitting multiple sclerosis patients. Eur. J. Immunol. 2010, 40, 888–898. [Google Scholar] [CrossRef]
- Thamilarsan, M.; Koczan, D.; Hecker, M.; Brigitte, P.; Zettl, U. MicroRNAs in multiple sclerosis and experimental autoimmune encephalomyeilitis. Autimmune. Rev. 2012, 11, 174–179. [Google Scholar] [CrossRef]
- Aslani, S.; Jafari, N.; Reza, M.; Javan, R.; Karami, J.; Ahmadi, M.; Jafarnejad, M. Epigenetic modifications and therapy in multiple sclerosis. Neuromol. Med. 2017, 19, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Martino, G.; Poliani, P.L.; Furlan, R.; Marconi, P.; Glorioso, J.C.; Adorini, L.; Comi, G. Cytokine therapy in immune-mediated demyelinating diseases of the central nervous system: a novel gene therapy approach. J. Neuroimmunol. 2000, 107, 184–190. [Google Scholar] [CrossRef]
- Navikas, V.; Link, H. Review: cytokines and the pathogenesis of multiple sclerosis. J. Neurosci. Res. 1996, 45, 322–333. [Google Scholar] [CrossRef]
- Dasgupta, S.; Ray, S.K. Insights into abnormal sphingolipid metabolism in multiple sclerosis: targeting ceramide biosynthesis as a unique therapeutic strategy. Ther. Targets Neurol. Dis. 2017, 4, e1598. [Google Scholar] [CrossRef] [PubMed]
- Foster, V.; Macfarlane, E.B. Clear Thinking about Alternate Therapies, Staying Well; National Multiple Sclerosis Society: New York, NY, USA, 2011; pp. 1–22. Available online: www.nationalMSsociety.org.
- Kanno, T.; Nishimoto, T.; Fujita, Y.; Goto, A.; Nakano, T.; Nishizaki, T. Sphingosine induces apoptosis in MKN-28 human gastric cancer cells in an SDK-dependent manner. Cell Physiol. Biochem. 2012, 30, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Ullio, C.; Casas, J.; Brunk, U.T.; Sala, G.; Fabrias, G.; Ghidoni, R.; Bonelli, G.; Baccino, F.M.; Autelli, R. Sphingosine mediates TNFα-induced lysosomal membrane permeabilization and ensuing programmed cell death in hepatoma cells. J. Lipid Res. 2012, 53, 1134–1143. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.C.; Martin, S.; Doyle, B.T.; Houghton, J.A. Sphingosine-induced apoptosis in rhabdomyosarcoma cell lines is dependent on pre-mitochondrial Bax activation and post-mitochondrial caspases. Cancer Res. 2007, 67, 756–764. [Google Scholar] [CrossRef]
- Stockman-Juvala, H.; Savolainen, K. A review of the toxic effects and mechanism of action of fumonisin B1. Hum. Exp. Toxicol. 2008, 27, 799–809. [Google Scholar] [CrossRef]
- Dasgupta, S.; Hogan, E.L. Chromatographic resolution and quantitative assay of CNS tissue sphingoids and sphingolipids. J. Lipid Res. 2001, 42, 301–308. [Google Scholar]
- Dasgupta, S.; Everhart, M.; Bhat, N.; Hogan, E.L. Neutral monoglycosylceramides in rat brain: occurrence, molecular expression and developmental variation. Dev. Neurosci. 1997, 19, 152–161. [Google Scholar] [CrossRef]
- Dasgupta, S.; Levery, S.B.; Hogan, E.L. 3-O-acetyl-sphingosine-series myelin glycolipids: characterization of novel 3-O-acetyl-sphingosine galactosylceramide. J. Lipid Res. 2002, 43, 751–761. [Google Scholar] [PubMed]
- Hoffmann, A.; Grimm, C.; Kraft, R.; Goldbaum, O.; Wrede, A.; Nolte, C.; Hanisch, U.K.; Richter-Landsberg, C.; Brück, W.; Kettenmann, H.; et al. TRPM3 is expressed in sphingosine-responsive myelinating oligodendrocytes. J. Neurochem. 2010, 114, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Bell, R.M. The sphingomyelin cycle and the second messenger function of ceramide. Science 1989, 243, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Kong, J.; Bieberich, E. Phytoceramide in vertebrate tissues: one step chromatography separation for molecular characterization of ceramide species. PLoS One 2013, 8, e80841. [Google Scholar] [CrossRef] [PubMed]
- Raymond, G.V. Leukodystrophy: Basic and Clinical. Adv. Neurobiol. 2017, 15, 365–382. [Google Scholar]
- Pavuluri, P.; Vadakedath, S.; Gundu, R.; Uppulety, S.; Kandi, V. Krabbe disease: Report of a rare lipid storage and neurodegenerative disorder. Cureus 2017, 9, e949. [Google Scholar] [CrossRef]
- Potter, G.B.; Petryniak, M.A. Neuroimmune mechanisms in Krabbe’s disease. J. Neurosci. Res. 2016, 94, 1341–1348. [Google Scholar] [CrossRef]
- Saffari, A.; Kölker, S.; Hoffmann, G.F.; Ebrahimi-Fakhari, D. Linking mitochondrial dysfunction to neurodegeneration in lysosomal storage diseases. J. Inherit. Metab. Dis. 2017, 5. [Google Scholar] [CrossRef]
- Lai, M.K.; Chew, W.S.; Torta, F.; Rao, A.; Harris, G.L.; Chun, J.; Herr, D.R. Biological Effects of Naturally Occurring Sphingolipids, Uncommon Variants, and Their Analogs. Neuromolecular Med. 2016, 18, 396–414. [Google Scholar] [CrossRef]
- Chakrabarti, S.S.; Bir, A.; Poddar, J.; Sinha, M.; Ganguly, A.; Chakrabarti, S. Ceramide and Sphingosine-1-Phosphate in Cell Death Pathways: Relevance to the Pathogenesis of Alzheimer Disease. Curr. Alzheimer. Res. 2016, 3, 1232–1248. [Google Scholar] [CrossRef]
- Chaurasia, B.; Summers, S.A. Ceramides - Lipotoxic Inducers of Metabolic Disorders. Trends Endocrinol. Metab. 2015, 26, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Coant, N.; Sakamoto, W.; Mao, C.; Hannun, Y.A. Ceramidases, roles in sphingolipid metabolism and in health and disease. Adv. Biol. Regul. 2017, 63, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Sandhoff, K. Neuronal sphingolipidoses: Membrane lipids and sphingolipid activator proteins regulate lysosomal sphingolipid catabolism. Biochimie 2016, 130, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Söderström, M. Clues to the immunopathogenesis of multiple sclerosis by investigating untreated patients during the very early stage of disease. Neurol. Sci. 2001, 22, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Martino, G.; Furlan, R.; Brambilla, E.; Bergami, A.; Ruffini, F.; Gironi, M.; Poliani, P.L.; Grimaldi, L.M.; Comi, G. Cytokines and immunity in multiple sclerosis: The dual signal hypothesis. J. Neuroimmunol. 2000, 109, 3–9. [Google Scholar] [CrossRef]
- Deckx, N.; Lee, W.P.; Berneman, Z.N.; Cools, N. Neuroendocrine immunoregulation in multiple sclerosis. Clin. Dev. Immunol. 2013, 2013, 705232. [Google Scholar] [CrossRef] [PubMed]
- Genain, C.P.; Cannella, B.; Hauser, S.L.; Raine, C.S. Identification of autoantibodies associated with myelin damage in multiple sclerosis. Nat. Med. 1999, 5, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M. Mechanisms of inflammation in MS tissue: Adhesion molecules and chemokines. J. Neuroimmunol. 1999, 98, 57–68. [Google Scholar] [CrossRef]
- Kieseier, B.C.; Storch, M.K.; Archelos, J.J.; Martino, G.; Hartung, H.P. Effector pathways in immune mediated central nervous system demyelination. Curr. Opin. Neurol. 1999, 12, 323–336. [Google Scholar] [CrossRef]
- Checa, A.; Khademi, M.; Sar, D.G.; Haeggstrom, J.Z.; Lundberg, J.O.; Piehl, F.; Olsson, T.; Wheelock, C.E. Hexosylceramides as intrathecal markers of worsening disability in multiple sclerosis. Mult. Scler. 2015, 21, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.; Bandaru, V.V.; Calabresi, P.A.; Nath, A.; Haughey, N.J. A defect of sphingolipid metabolism modifies properties of normal appearing whote matter in multiple sclerosis. Brain 2008, 131, 3092–3102. [Google Scholar] [CrossRef] [PubMed]
- Vidaurre, O.G.; Haines, J.D.; Katz, S.I.; Adula, K.P.; Huynh, J.L.; McGraw, C.A.; Zhang, F.; Varghese, M.; Sotirchos, E.; Bhargava, P.; et al. Cerebrospinal fluid ceramides from patients with multiple sclerosis impairs neuronal bioenergetics. Brain 2014, 137, 2271–2286. [Google Scholar] [CrossRef] [PubMed]
- Gakyung, L.; Hasan, H.; Kwon, O.-S.; Jung, B.H. Identification of altered metabolic pathways during disease progression in EAE mice via metabolics and lipidomics. Neuroscience 2019, 416, 74–87. [Google Scholar]
- Slezak-Prochazka, I.; Durmus, S.; Kroesen, B.J.; van den Berg, A. MicroRNAs, macrocontrol: regulation of miRNA processing. RNA 2010, 16, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Jahangir, J.; Bakillah, A.; Iqbal, J. Regulation of sphingolipid metabolism by micro-RNAs: A potential approach to alleviate aetherosclerosis. Diseases 2018, 6, 82. [Google Scholar] [CrossRef] [PubMed]
- Lau, P.; Verrier, J.D.; Nielsen, J.A.; Johnson, K.R.; Notterpek, L.; Hudson, L.D. Identification of dynamically regulated microRNA and mRNA networks in developing oligodendrocytes. J. Neurosci. 2008, 28, 11720–11730. [Google Scholar] [CrossRef] [PubMed]
- Geekiyanage, H.; Chan, C. Micro-RNA137/181c regulates serine palmitoyltransferase and in turn amyloid beta, novel targets in sporadic Alzheimer’s disease. J. Neurosci. 2011, 31, 14820–14830. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Medrano, J.; Yang, B.; Garza, N.T.; Segura-Ulate, I.; Perez, R.Z. Up-regulation of protective neuronal microRNAs by FTY720 and novel FTY720-derivatives. Neurosci. Lett. 2019, 690, 178–190. [Google Scholar] [CrossRef]
- Wang, C.; Ji, B.; Cheng, B.; Bai, B. Neuroprtection of microRNA in Neurological disorders (review). Biomed. Rep. 2014, 2, 611–619. [Google Scholar] [CrossRef]
- Quinlan, S.; Kenny, A.; Medina, M.; Engel, T.; Jimenez-Mateos, E.M. MicroRNAs in neurodegenerative diseases. Int. Rev. Cell Mol. Biol. 2017, 334, 309–343. [Google Scholar]
- Brenan, S.; Keon, M.; Liu, B.; Su, Z.; Saksena, M.K. Panoramic visualization of circulating microRNAs across neurodegenerative diseases in humans. Mol. Neurobiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Chuturgoon, A.A.; Phulukdaree, A.; Moodley, D. Fumonisin B1 modulates expression of human cytochrome P450 1b1 in human hepatoma (Hepg2) cells by repressing mir-27b. Toxicol. Lett. 2014, 227, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.K.; Xu, X.M. MicroRNA in central nervous system trauma and degenerative disorders. Physiol. Genomics 2011, 43, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Nampoothiri, S.S.; Rajanikant, G.K. miR-9 upregulation integrates post-ischemic neuronal survival and regeneration in vitro. Cell Mol. Neurobiol. 2019, 39, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Schratt, G.M.; Tuebing, F.; Nigh, E.A.; Kane, C.G.; Sabatini, M.E.; Kiebler, M.; Greenberg, M.E. A brain-specific micro RNA regulates dendritic spine development. Nature 2006, 439, 283–289. [Google Scholar] [CrossRef]
- Mix, E.; Meyer-Rienecker, H.; Hartung, H.P.; Zettl, U.K. Animal models of multiple sclerosis-potentials and limitations. Prog. Neurobiol. 2010, 92, 386–404. [Google Scholar] [CrossRef] [PubMed]
- Lucchinetti, C.; Bruck, W.; Parisi, J.; Scheithauer, B.; Rodriguez, M.; Lassmann, H. Heterogeneity of multiple sclerosis lesions: Implications for the pathogenesis of demyelination. Ann. Neurol. 2000, 47, 707–717. [Google Scholar] [CrossRef]
- Comabella, M.; Sastre-Garriga, J.; Montalban, X. Precision medicine in multiple sclerosis: biomarkers for diagnosis, prognosis, and treatment response. Curr. Opin. Neurol. 2016, 29, 254–262. [Google Scholar] [CrossRef]
- Morris, G.; Reschke, C.R.; Henshall, D.C. Targeting microRNA-134 for seizure control and disease modification in epilepsy. EBioMedicine 2019, 45, 646–654. [Google Scholar] [CrossRef]
- Zhou, B.; Wang, S.; Mayr, C.; Bartel, D.P.; Lodish, H.F. Mir-150, a microRNA expressed in mature B and T cells, blocks early B cell develpoment when expressed prematurely. Proc. Natl. Acad. Sci. USA 2007, 107, 7080–7085. [Google Scholar] [CrossRef]
- O’Connell, R.M.; Kahn, D.; Gibson, W.S.; Round, J.L.; Scholz, A.L.R.l.; Cahudhuri, A.A.; Kahn, M.E.; Rao, D.S.; Baltimore, D. MicrRNA-155 promotes autoimmune inflammation by enhancing inflammatory T-cell development. Immunity 2010, 33, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.F.; Boldin, M.P.; Chaudhury, A.; Lin, L.L.; Taganov, K.D.; Hanada, T.; Yoshimura, A.; Baltimore, D.; Rudensky, A.Y. Function of miR-146a in controlling Treg cell-mediated regulation of Th 1 response. Cell 2010, 142, 914–929. [Google Scholar] [CrossRef] [PubMed]
- Niimoto, T.; Nakasa, T.; Ishikawa, M.; Okuhara, A.; Izumi, B.; Deie, M.; Suzuki, O.; Adachi, N.; Ochi, M. Micro-RNA-146a expresses in interleukin-17 producing T-cells in rheumatoid arthritis patients. BMC Muscuoskelet Disord 2010, 11, 209. [Google Scholar]
- Octaegui, D.; Mostafavi, S.; Bernard, C.C.; Lopez de Munain, A.; Mousavi, P.; Oksenberg, J.R.; Baranzini, S.E. Increased transcriptional activity of milk-related genes following the active phase of experimental autoimmune encephalomyelitis and multiple sclerosis. J. Immunol. 2007, 179, 4074–4082. [Google Scholar] [CrossRef] [PubMed]
- Garo, L.P.; Murugaiyan, G. Contribution of microRNAs to autoimmune diseases. Cell Mol. Life Sci. 2016, 73, 2041–2051. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cheng, Y.; Cui, W.; Li, M.; Li, B.; Guo, L. MicroRNA-155 modulates Th1 and Th17 cell differentiation and is associated with multiple sclerosis and experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2014, 266, 56–63. [Google Scholar] [CrossRef]
- Du, C.; Liu, C.; Kang, J.; Zhao, G.; Ye, Z.; Huang, S.; Li, Z.; Wu, Z.; Pei, G. MicroRNA miR-326 regulates TH-17 differentiation and is associated with the pathogenesis of multiple sclerosis. Nat. Immunol. 2009, 10, 1252–1259. [Google Scholar] [CrossRef]
- Mycko, M.P.; Cichalewska, M.; Cwiklinska, H.; Selmaj, K.W. miR-155-3p Drives the development of autoimmune demyelination by regulation of heat shock protein 40. J. Neurosci. 2015, 35, 16504–16515. [Google Scholar] [CrossRef]
- Huang, X.; Schwind, S.; Yu, B.; Santhanam, R.; Wang, H.; Hoeller-bauer, P.; Mims, A.; Klisovic, R.; Walker, A.R.; Chan, K.K.; et al. Targeted delivery of microRNA-29b by transferrin-conjugated anionic lipopolyplex nanoparticles: a novel therapeutic strategy in acute myeloid leukemia. Clin. Cancer Res. 2013, 19, 2355–2367. [Google Scholar] [CrossRef]
- Babar, I.A.; Cheng, C.J.; Booth, C.J.; Liang, X.; Weidhass, J.B.; Saltzman, W.M.; Slack, F.J. Nano-particles based therapy in an in vivo microRNA-155 (miR-155) dependent mouse model of lymphoma. Proc. Natl. Acad. Sci. USA 2009, 109, E1695–E1704. [Google Scholar] [CrossRef]
- Wang, H.; Moyano, A.L.; Ma, Z.; Deng, Y.; Lin, Y.; Zhao, C.; Zhang, L.; Jiang, M.; He, X.; Ma, Z.; et al. miR-219 Cooperates with miR-338 in myelination and promotes myelin repair in the CNS. Dev. Cell 2017, 40, 566–582. [Google Scholar] [CrossRef]
- Chun, J.; Hartung, H.P. Mechanism of action of oral fingolimod (FTY720) in multiple sclerosis. Clin. Neropharmacol. 2010, 33, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Cao, K.; Kato, S.; Komizu, Y.; Mizutani, N.; Tanaka, K.; Arima, C.; Tai, M.C.; Yanagisawa, K.; Togawa, N.; et al. Targeting ceramide synthase 6-dependent metastasis-prone phenotype in lung cancer cells. J. Clinic. Invest. 2016, 126, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Sajja, V.S.S.; Jablonska, A.; Haughey, N.; Bulte, J.W.M.; Stevens, R.D.; Long, J.B.; Walczak, P.; Janowski, M. Sphingolipids and micrRNA changes in blood following blast traumatic brain injury. J. Neurotrauma. 2018, 35, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Piket, E.; Zheleznyakova, G.Y.; Kular, L.; Jagodic, M. Small non-coding RNAsas important players, biomarkers and therapeutic targets in multiple sclerosis: A comprehensive overview. J. Autoimmun. 2019, 101, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Fenoglio, C.; De Riz, M.; Pietroboni, A.M.; Calvi, A.; Serpente, M.; Cioffi, S.M.; Arcaro, M.; Oldoni, E.; Scarpini, E.; Galimberti, D. Effect of fingolimod treatment on circulating miR-15b, miR23a and miR-223 levels in patients with multiple sclerosis. J. Neuroimmunol. 2016, 299, 81–83. [Google Scholar] [CrossRef] [PubMed]
- Eftekharian, M.M.; Komaki, A.; Mazdeh, M.; Arsang-Jang, S.; Taheri, M.; Ghafouri-Fard, S. Expression profile of selected microRNAs in the peripheral blood of Multiple Sclerosis patients: a multivariate statistical analysis with ROC curve to find new biomarkers for fingolimod. J. Mol. Neursci. 2019, 68, 153–161. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).