Protective Effects of Novel Antioxidant Peptide Purified from Alcalase Hydrolysate of Velvet Antler Against Oxidative Stress in Chang Liver Cells In Vitro and in a Zebrafish Model In Vivo



Abstract
:1. Introduction
2. Results
2.1. Preparation of Enzymatic Hydrolysates from Velvet Antler and Their Peroxyl Radical Scavenging Activities
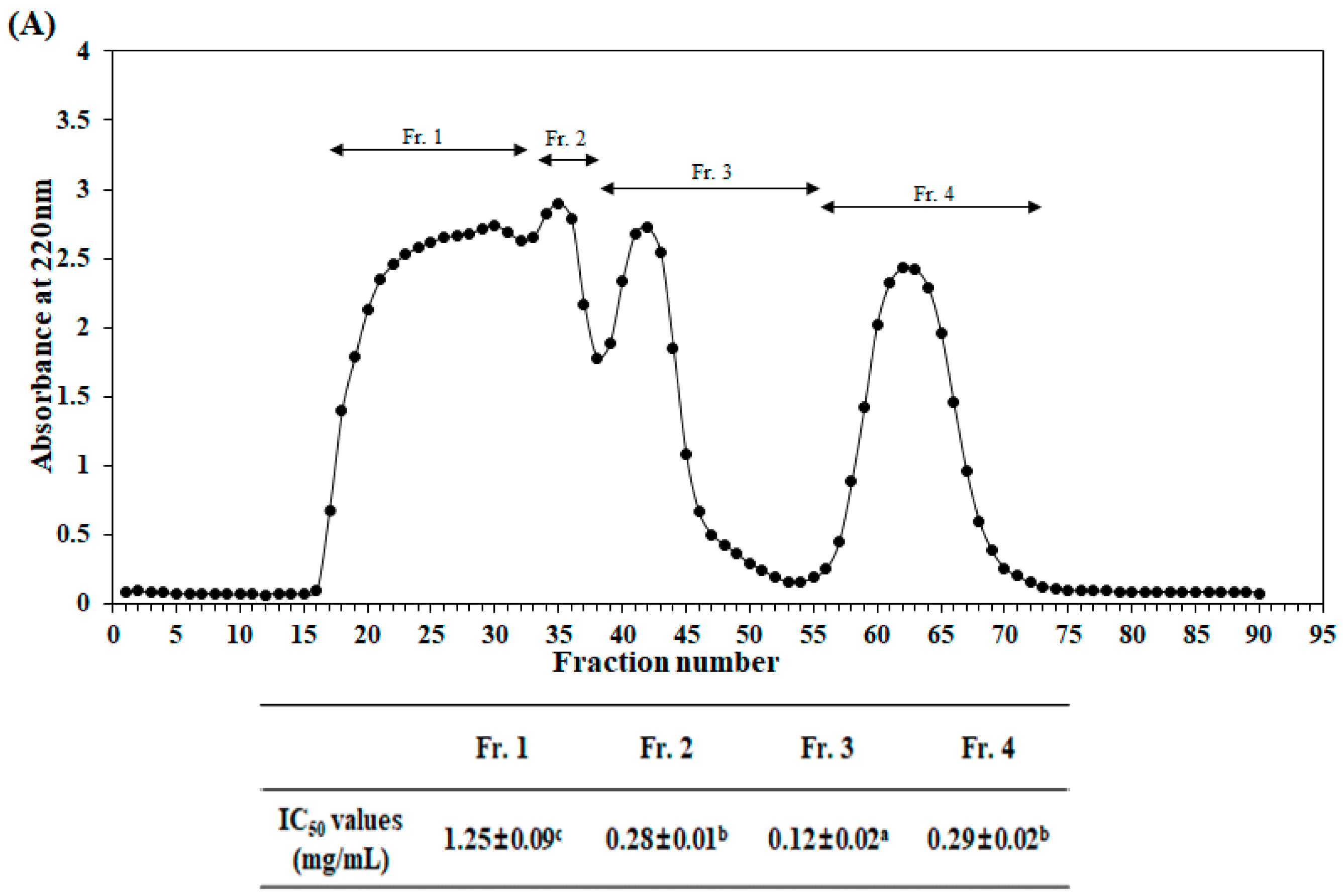
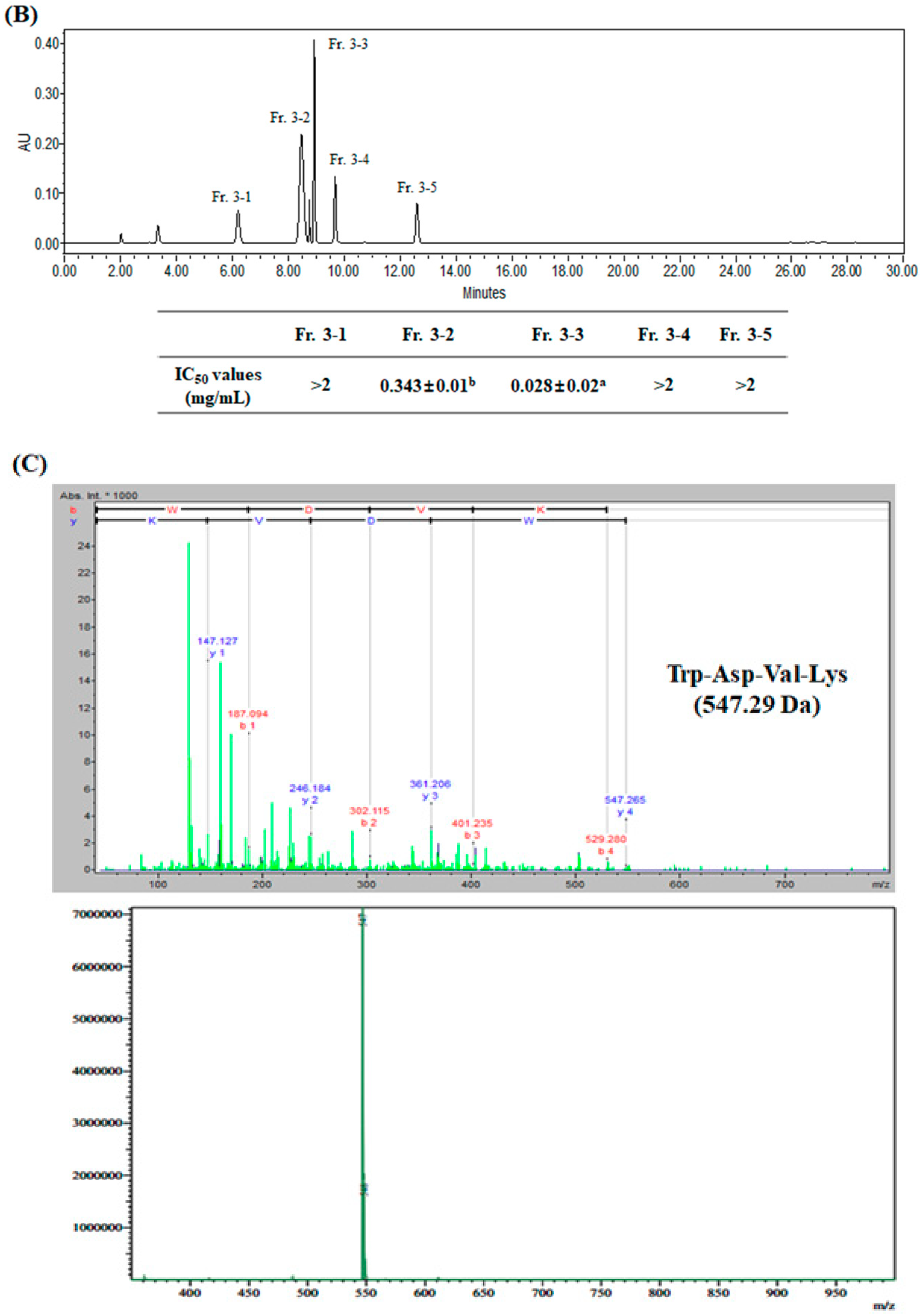
2.2. Purification and Identification of Antioxidant Peptide
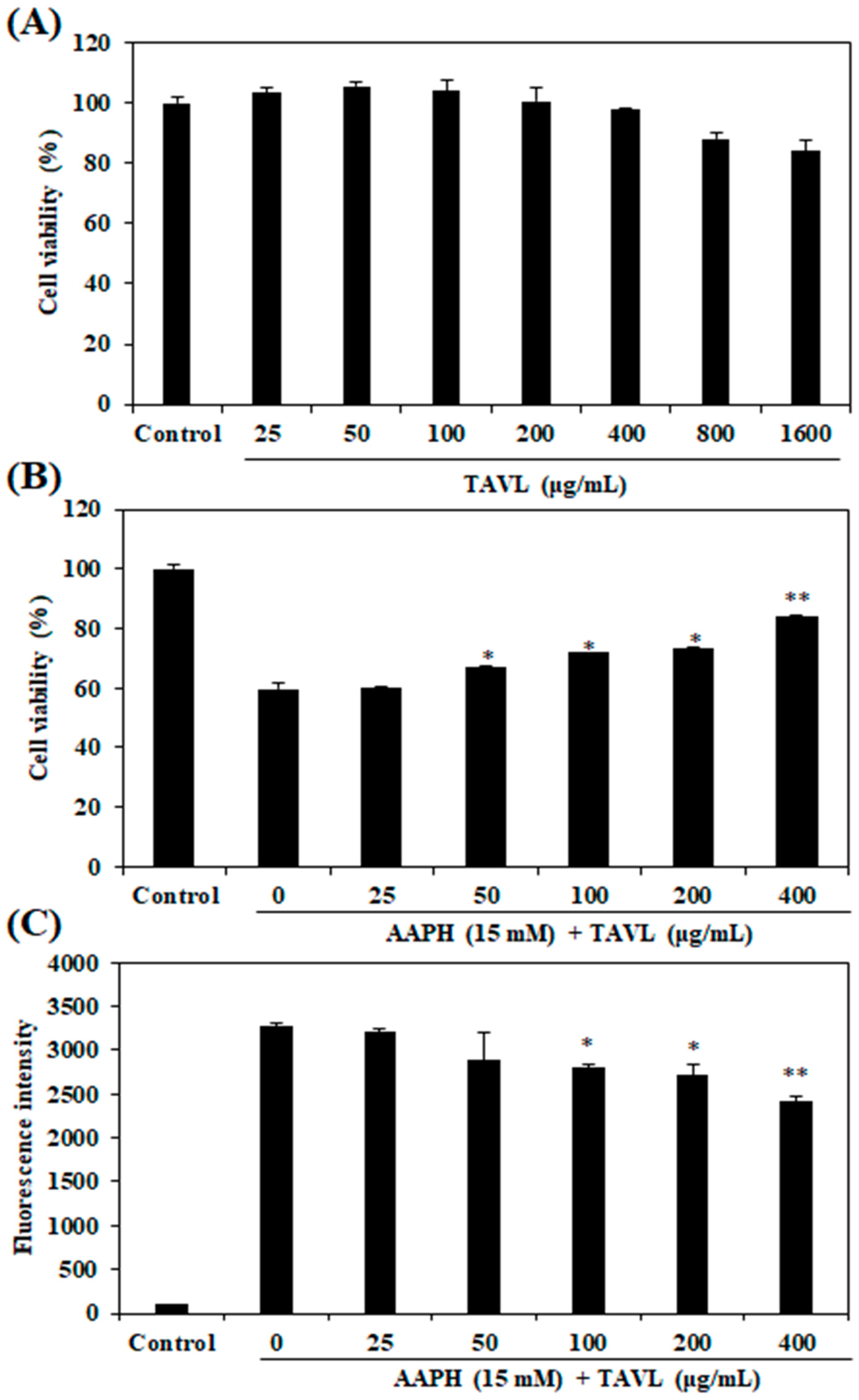
2.3. Intracellular Antioxidant Activities of the Purified Peptide
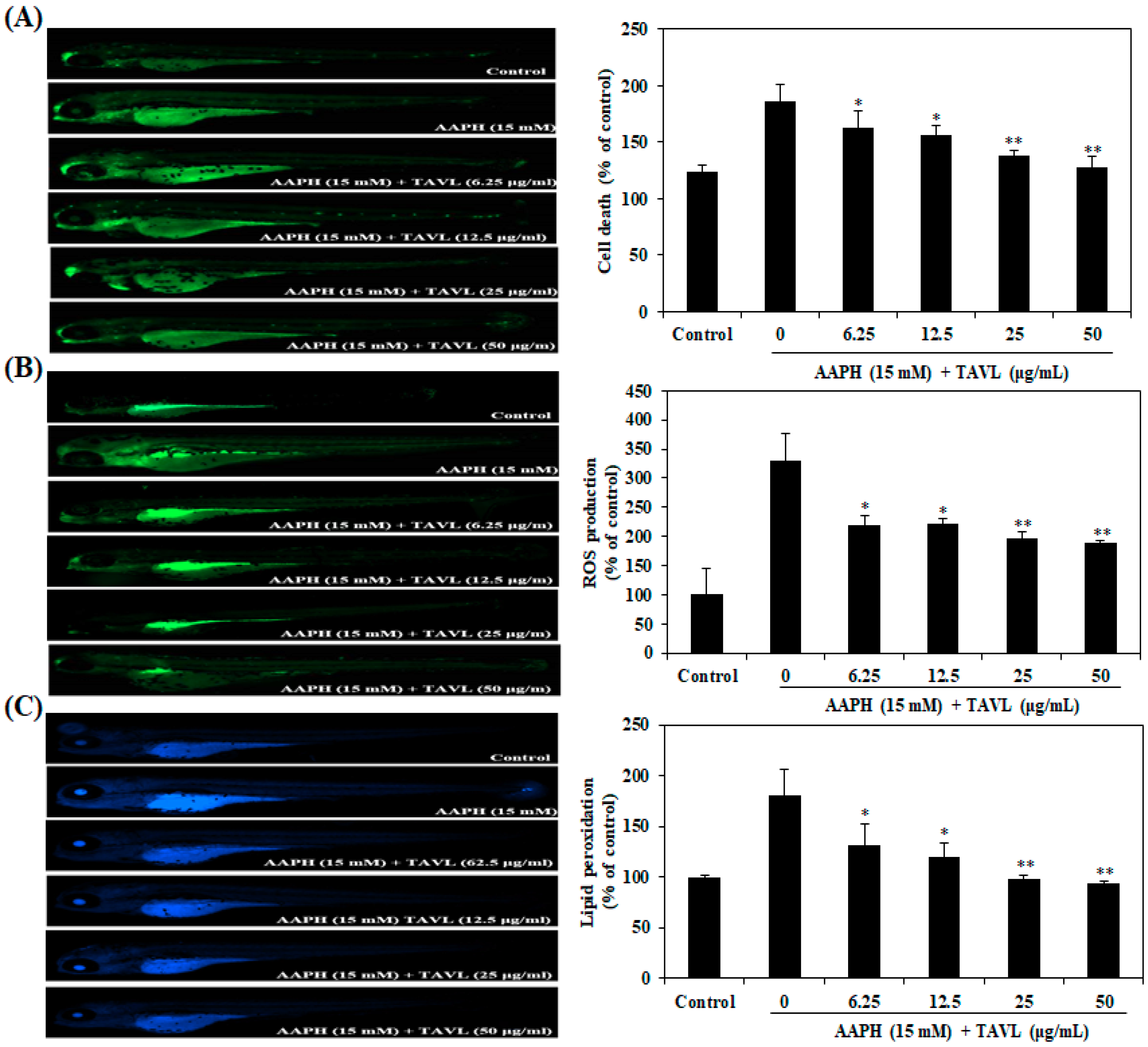
2.4. Protective Effects of Antioxidant Peptide (TAVL) against AAPH-Induced Oxidative Stress in a Zebrafish Model In Vivo
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Sample Preparation
4.3. Preparation of Enzymatic Hydrolysates from Velvet Antler
4.4. Peroxyl Radical Scavenging Activity
4.5. Isolation of Antioxidant Peptides from the Enzymatic Hydrolysate of Velvet Antler
4.5.1. Fractionation According to the Molecular Weight
4.5.2. Purification of Antioxidant Peptides
4.5.3. Identification of Purified Antioxidant Peptide
4.6. Experiments for Antioxidant Activity Assay Using Cells
4.6.1. Cell Culture
4.6.2. Measuring Cytoprotective Effect by MTT Assay
4.6.3. Intracellular ROS Measurement
4.7. In Vivo Zebrafish Model for Antioxidant Activity Assay
4.8. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Qian, Z.J.; Jung, W.K.; Kim, S.K. Free radical scavenging activity of a novel antioxidative peptide purified from hydrolysate of bullfrog skin, Rana catesbeiana Shaw. Bioresour. Technol. 2008, 99, 1690–1698. [Google Scholar] [CrossRef]
- Halliwell, B. Free radicals, antioxidants, and human disease: Curiosity, cause, or consequence. Lancet 1994, 344, 721–724. [Google Scholar] [CrossRef]
- Ito, N.; Hirose, M.; Fukushima, S.; Tsuda, H.; Shirai, T.; Tatematsu, M. Studies on antioxidants: Their carcinogenic and modifying effects on chemical carcinogenesis. Food Chem. Toxicol. 1986, 24, 1071–1082. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef]
- Byun, H.G.; Lee, J.K.; Park, H.G.; Jeon, J.K.; Kim, S.K. Antioxidant peptides isolated from the marine rotifer, Brachionus rotundiformis. Process Biochem. 2009, 44, 842–846. [Google Scholar] [CrossRef]
- Ko, S.C.; Kim, D.; Jeon, Y.J. Protective effect of a novel antioxidative peptide purified from a marine Chlorella ellipsoidea protein against free radical-induced oxidative stress. Food Chem. Toxicol. 2012, 50, 2294–2302. [Google Scholar] [CrossRef]
- Davalos, A.; Miguel, M.; Bartolome, B.; Lopez-Fandino, R. Antioxidant activity of peptides derived from egg white proteins by enzymatic hydrolysis. J. Food Prot. 2004, 67, 1939–1944. [Google Scholar] [CrossRef]
- Guo, H.; Kouzuma, Y.; Yonekura, M. Structures and properties of antioxidative peptides derived from royal jelly protein. Food Chem. 2009, 113, 238–245. [Google Scholar] [CrossRef]
- Lafarga, T.; Hayes, M. Bioactive peptides from meat muscle and by-products: Generation functionality and application as functional ingredients. Meat Sci. 2014, 98, 227–239. [Google Scholar] [CrossRef]
- Wu, F.; Li, H.; Jin, L.; Li, X.; Ma, Y.; You, J.; Xu, Y. Deer antler base as a traditional Chinese medicine: A review of its traditional uses, chemistry and pharmacology. J. Ethnopharmacol. 2013, 145, 403–415. [Google Scholar] [CrossRef]
- Sui, Z.; Zhang, L.; Huo, Y.; Zhang, Y. Bioactive components of velvet antlers and their pharmacological properties. J. Pharm. Biomed. Anal. 2014, 87, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Luo, Y.C.; Wang, C.T.; Ji, B.P. Antioxidant activity of protein hydrolysates from aqueous extract of velvet antler (Cervus elaphus) as influenced by molecular weight and enzymes. Nat. Prod. Commun. 2011, 6, 1683–1688. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Yang, H.W.; Ding, Y.; Wang, Y.; Jeon, Y.J.; Moon, S.H.; Jeon, B.T.; Sung, S.H. Anti-inflammatory effects of enzymatic hydrolysates of velvet antler in Raw 264.7 cells in vitro and zebrafish model. EXCLI J. 2015, 14, 1122–1132. [Google Scholar] [PubMed]
- Ding, Y.; Wang, Y.; Jeon, B.T.; Moon, S.H.; Lee, S.H. Enzymatic hydrolysate from velvet antler suppresses adipogenesis in 3T3-L1 cells and attenuates obesity in high-fat diet mice. EXCLI J. 2017, 16, 328–339. [Google Scholar] [PubMed]
- Zhao, L.; Wang, X.; Zhang, X.L.; Xie, Q.F. Purification and identification of anti-inflammatory peptides derived from simulated gastrointestinal digests of velvet antler protein (Cervus elaphus Linnaeus). J. Food Drug Anal. 2016, 24, 376–384. [Google Scholar] [CrossRef]
- Lee, J.K.; Hong, S.; Jeon, J.K.; Kim, S.K.; Byun, H.G. Purification and characterization of angiotensin I converting enzyme inhibitory peptides from the rotifer, brachionus rotundiformis. Bioresour. Technol. 2009, 100, 5255–5259. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, S.Y.; Fernando, I.P.S.; Sanjeewa, K.K.A.; Wang, L.; Lee, S.H.; Ko, S.C.; Kang, M.C.; Jayawardena, T.U.; Jeon, Y.J. Free radical scavenging activity of the peptide from the Alcalase hydrolysate of the edible aquacultural seahorse (Hippocampus abdominalis). J. Food Biochem. 2019, 43, e12833. [Google Scholar] [CrossRef]
- Saito, Y.; Wanezaki, K.; Kawato, A.; Imayasu, S. Structure and activity of angiotensin I converting enzyme inhibitory peptides from sake and sake lees. Biosci. Biotechnol. Biochem. 1994, 58, 1767–1771. [Google Scholar] [CrossRef]
- Sunwoo, H.H.; Nakano, T.; Hudson, R.J.; Sim, J.S. Chemical composition of antlers from wapiti (Cervus elaphus). J. Agric. Food Chem. 1995, 43, 2846–2849. [Google Scholar] [CrossRef]
- Li, G.H.; Wan, J.Z.; Le, G.W.; Shi, Y.H. Novel angiotensin I-converting enzyme inhibitory peptides isolated from alcalase hydrolysate of mung bean protein. J. Pept. Sci. 2006, 12, 509–514. [Google Scholar] [CrossRef]
- Wijesekara, I.; Qian, Z.J.; Ryu, B.M.; Ngo, D.H.; Kim, S.K. Purification and identification of antihypertensive peptides from seaweed pipefish (Syngnathus schlegeli) muscle protein hydrolysate. Food Res. Int. 2011, 44, 703–707. [Google Scholar] [CrossRef]
- Byun, H.G.; Kim, S.K. Purification and chracterization of angiotensin I converting enzyme (ACE) inhibitory peptides from Alaska pollack (Theragra chalogramma) skin. Process Biochem. 2001, 36, 1155–1162. [Google Scholar] [CrossRef]
- Sivakumar, R.; Hordur, G.K. ACE-inhibitory activity of tilapia protein hydrolysates. Food Chem. 2009, 117, 582–588. [Google Scholar]
- Ko, J.Y.; Lee, J.H.; Samarakoon, K.; Kim, J.S.; Jeon, Y.J. Purification and determination of two novel antioxidant peptides from flounder fish (Paralichthys olivaceus) using digestive proteases. Food Chem. Toxicol. 2013, 52, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Sheih, I.C.; Wu, T.K.; Fang, T.J. Antioxidant properties of a new antioxidative peptide from algae protein waste hydrolysate in different oxidation systems. Bioresour. Technol. 2009, 100, 3419–3425. [Google Scholar] [CrossRef]
- Rajapakse, N.; Mendis, E.; Byun, H.G.; Kim, S.K. Purification and in vitro antioxidative effects of giant squid muscle peptides on free radical-mediated oxidative systems. J. Nutr. Biochem. 2005, 16, 562–569. [Google Scholar] [CrossRef]
- Roberts, P.R.; Burney, J.D.; Black, K.W.; Zaloga, G.P. Effect of chain length on absorption of biologically active peptides from the gastrointestinal tract. Digestion 1999, 60, 332–337. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of aging. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Kim, E.A.; Lee, S.H.; Ko, C.I.; Cha, S.H.; Kang, M.C.; Kang, S.M.; Ko, S.C.; Lee, W.W.; Ko, J.Y.; Lee, J.H.; et al. Protective effect of fucoidan against AAPH-induced oxidative stress in zebrafish model. Carbohydr. Polym. 2014, 102, 185–191. [Google Scholar] [CrossRef]
- Kang, M.C.; Cha, S.H.; Wijesinghe, W.A.J.P.; Kang, S.M.; Lee, S.H.; Kim, E.A.; Jeon, Y.J. Protective effect of marine algae phlorotannins against AAPH-induced oxidative stress in zebrafish embryo. Food Chem. 2013, 138, 950–955. [Google Scholar] [CrossRef]
- Sevanian, A.; Hochstein, P. Mechanism and consequence of lipid peroxidation in biological systems. Annu. Rev. Nutr. 1985, 5, 365–390. [Google Scholar] [CrossRef] [PubMed]
- Hiramoto, K.; Johkoh, H.; Sako, K.I.; Kikugawa, K. DNA breaking activity of the carbon-centered radical generated from 2, 2′-azobis (2-amidinopropane) hydrochloride (AAPH). Free Radic. Res. Commun. 1993, 19, 323–332. [Google Scholar] [CrossRef]
- Rosenkranz, A.R.; Schmaldienst, S.; Stuhlmeier, K.M.; Chen, W.; Knapp, W.; Zlabinger, G.J. A microplate assay for the detection of oxidative products using 2′,7′-dichlorofluorescin-diacetate. J. Immunol. Methods 1992, 156, 39–45. [Google Scholar] [CrossRef]
- Lee, S.H.; Ko, C.I.; Jee, Y.; Jeong, Y.; Kim, M.; Kim, J.S.; Jeon, Y.J. Anti-inflammatory effect of fucoidan extracted from Ecklonia cava in zebrafish model. Carbohydr. Polym. 2013, 92, 84–89. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Extraction Yield (%) | Radical Scavenging Activity (%) 1) |
|---|---|---|
| Trypsin | 34.09 | 90.00 ± 1.69 |
| Pepsin | 12.39 | 78.58 ± 1.08 |
| α-Chymotrypsin | 38.96 | 89.54 ± 0.42 |
| Neutrase | 23.81 | 89.97 ± 1.05 |
| Alcalase | 29.75 | 93.78 ± 0.45 |
| Molecular Sizes | IC50 Value (mg/mL) |
|---|---|
| >10 kDa | 0.30 ± 0.01 b |
| 5–10 kDa | 0.35 ± 0.01 b |
| <5 kDa | 0.26 ± 0.02 a |
| IC50 Value (μM) | |
|---|---|
| TAVL | 51.16 ± 0.2 |
| Ascorbic acid | 19.26 ± 0.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Y.; Ko, S.-C.; Moon, S.-H.; Lee, S.-H. Protective Effects of Novel Antioxidant Peptide Purified from Alcalase Hydrolysate of Velvet Antler Against Oxidative Stress in Chang Liver Cells In Vitro and in a Zebrafish Model In Vivo. Int. J. Mol. Sci. 2019, 20, 5187. https://doi.org/10.3390/ijms20205187
Ding Y, Ko S-C, Moon S-H, Lee S-H. Protective Effects of Novel Antioxidant Peptide Purified from Alcalase Hydrolysate of Velvet Antler Against Oxidative Stress in Chang Liver Cells In Vitro and in a Zebrafish Model In Vivo. International Journal of Molecular Sciences. 2019; 20(20):5187. https://doi.org/10.3390/ijms20205187
Chicago/Turabian StyleDing, Yuling, Seok-Chun Ko, Sang-Ho Moon, and Seung-Hong Lee. 2019. "Protective Effects of Novel Antioxidant Peptide Purified from Alcalase Hydrolysate of Velvet Antler Against Oxidative Stress in Chang Liver Cells In Vitro and in a Zebrafish Model In Vivo" International Journal of Molecular Sciences 20, no. 20: 5187. https://doi.org/10.3390/ijms20205187
APA StyleDing, Y., Ko, S.-C., Moon, S.-H., & Lee, S.-H. (2019). Protective Effects of Novel Antioxidant Peptide Purified from Alcalase Hydrolysate of Velvet Antler Against Oxidative Stress in Chang Liver Cells In Vitro and in a Zebrafish Model In Vivo. International Journal of Molecular Sciences, 20(20), 5187. https://doi.org/10.3390/ijms20205187