Immune Complexes Impaired Glomerular Endothelial Cell Functions in Lupus Nephritis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
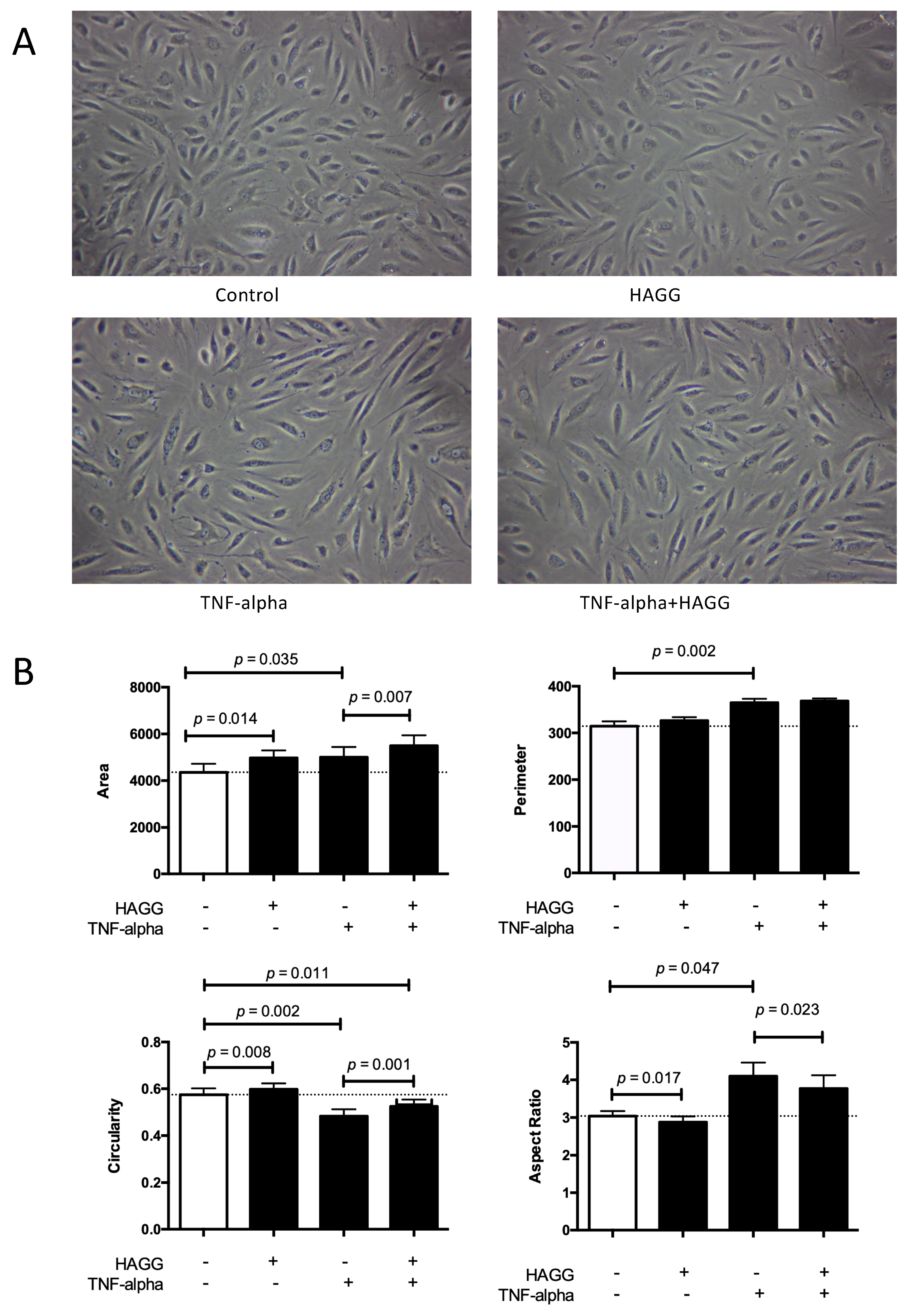
2.1. HAGG-induced GEC Morphology Changes
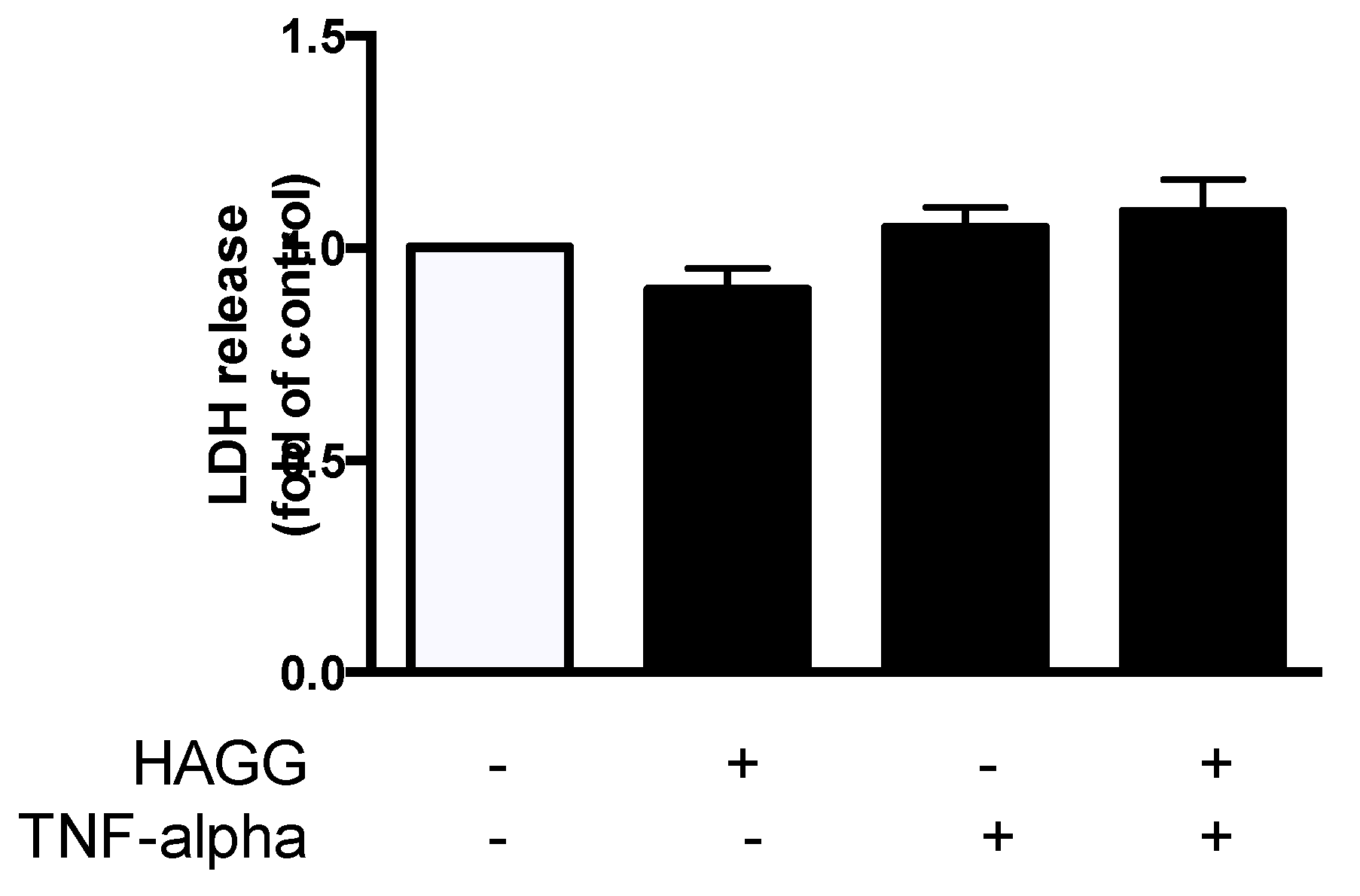
2.2. HAGG Did Not Induce Necrosis in GECs
2.3. HAGG Upregulated Intracellular Level of Active Caspase 3 in GECs
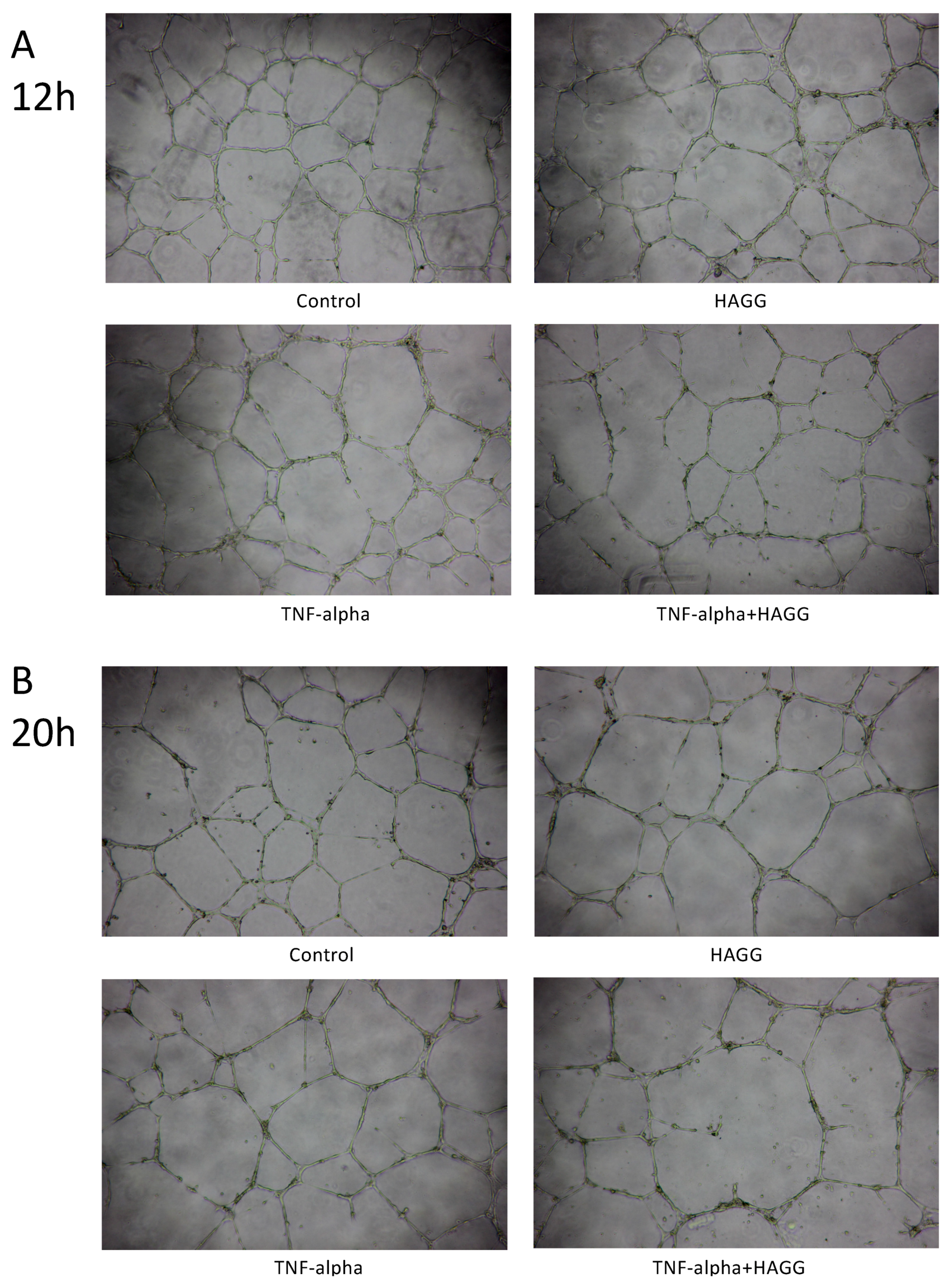
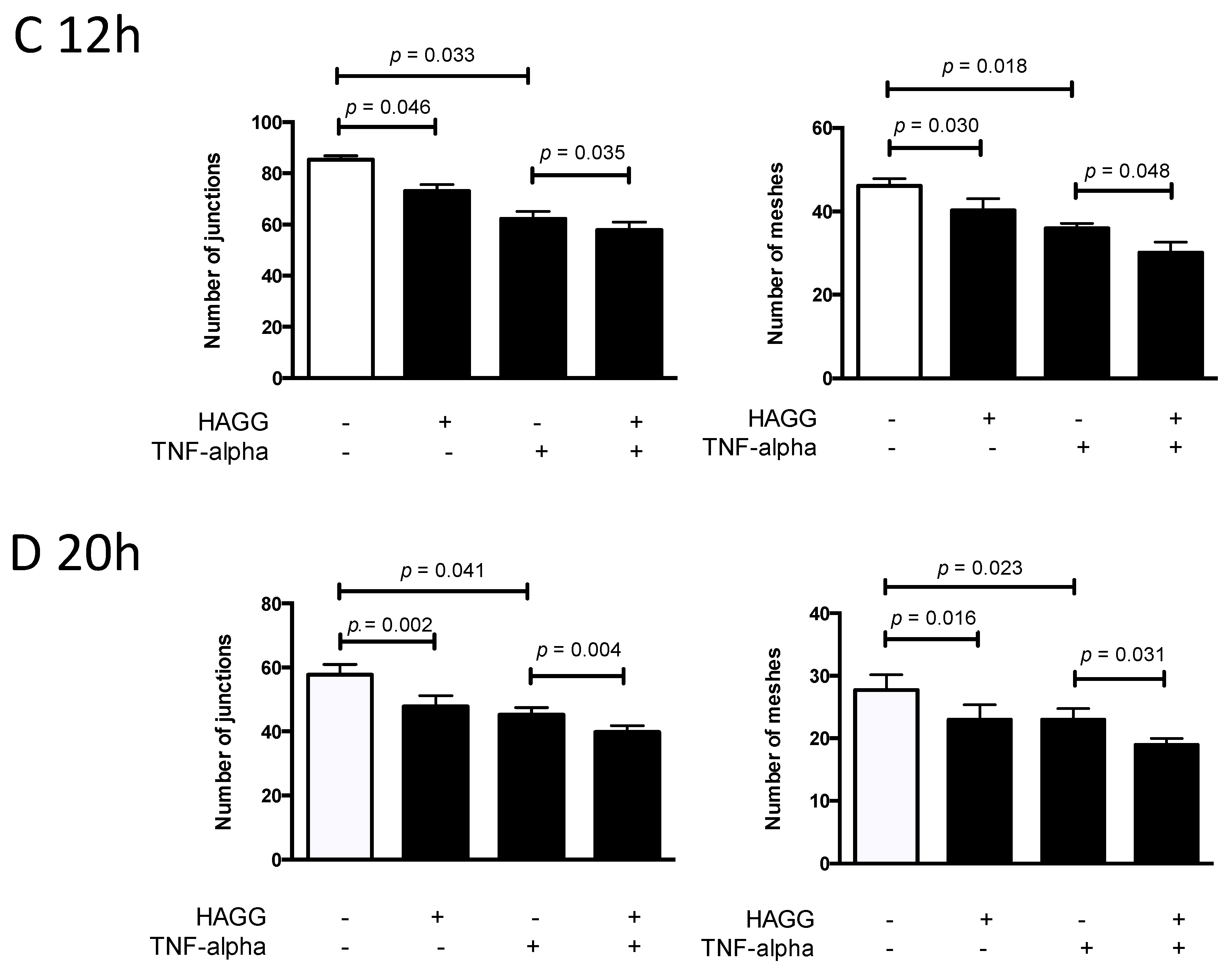
2.4. HAGG Suppressed GEC Tube Formation
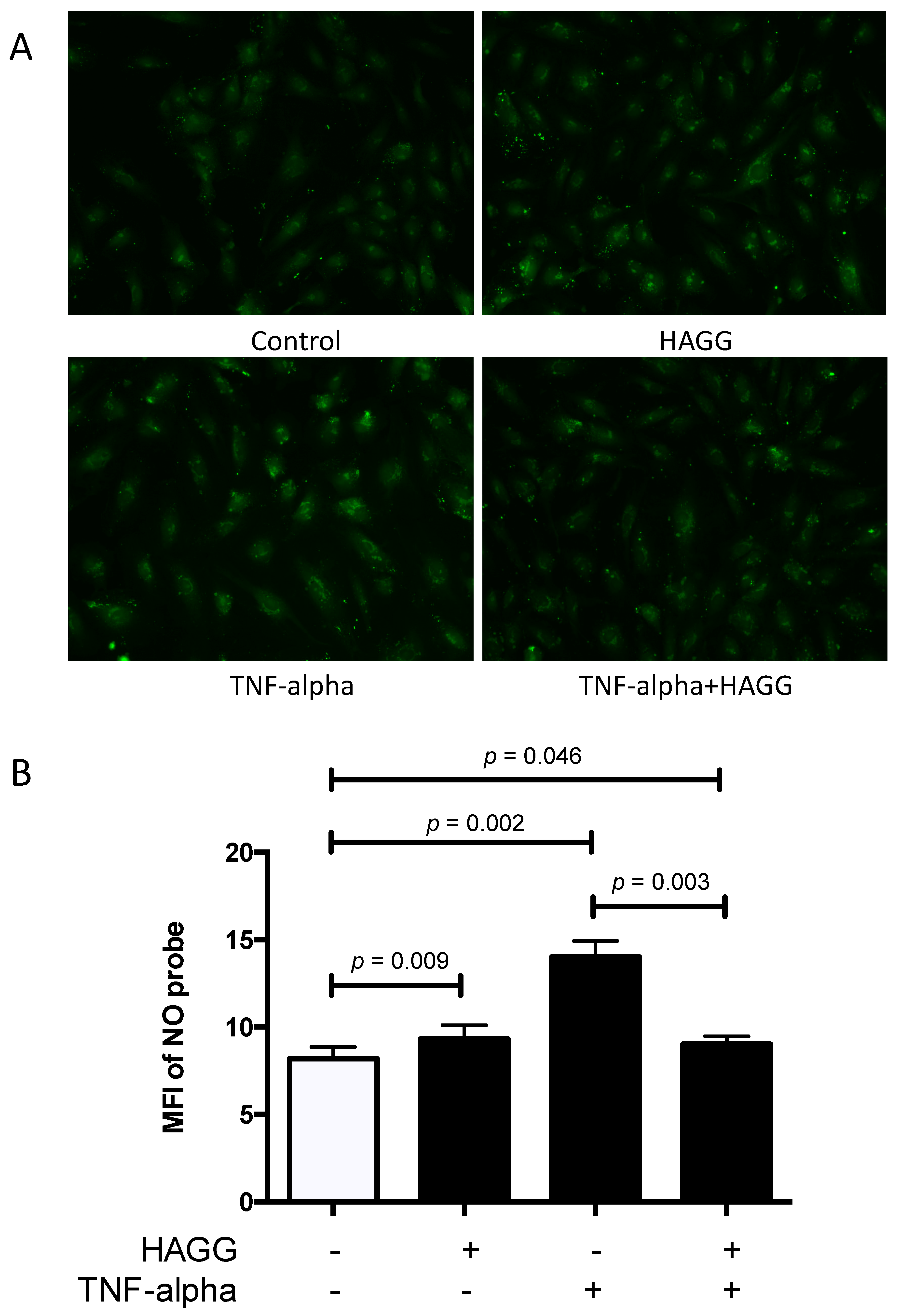
2.5. HAGG Alleviated the Intracellular Nitric Oxide Production in GECs Stimulated by TNF-α
3. Discussion
3.1. Cell Morphology and Cell Function
3.2. Angiogenesis
3.3. Intracellular NO Production
3.4. Cell Viability and Cell Death
4. Materials and Methods
4.1. Cell Culture
4.2. Heat-Aggregated Gamma Globulin Preparation
4.3. Cell Morphological Analysis
4.4. Apoptosis Assay by Flow Cytometry
4.5. Necrosis Measurement
4.6. Tube Formation Assay
4.7. Intracellular Nitric Oxide Measurement
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| eNOS | Endothelial nitric oxide synthase |
| GFM | Glomerular filtration membrane |
| GECs | Glomerular endothelial cells |
| HAGG | Heat-aggregated gamma globulin |
| HUVECs | Human umbilical vein endothelial cells |
| IL | Interleukin |
| LDH | Lactate dehydrogenase |
| LN | Lupus nephritis |
| MCP | Monocyte chemoattractant protein |
| MFI | Mean fluorescence intensity |
| NO | Nitric oxide |
| TNF | Tumor necrosis factor |
References
- Maroz, N.; Segal, M.S. Lupus nephritis and end-stage kidney disease. Am. J. Med. Sci. 2013, 346, 319–323. [Google Scholar] [CrossRef]
- Davidson, A. What is damaging the kidney in lupus nephritis? Nat. Rev. Rheumatol. 2016, 12, 143–153. [Google Scholar] [CrossRef]
- Lech, M.; Anders, H.J. The pathogenesis of lupus nephritis. JASN 2013, 24, 1357–1366. [Google Scholar] [CrossRef]
- Arkill, K.P.; Qvortrup, K.; Starborg, T.; Mantell, J.M.; Knupp, C.; Michel, C.C.; Harper, S.J.; Salmon, A.H.; Squire, J.M.; Bates, D.O.; et al. Resolution of the three dimensional structure of components of the glomerular filtration barrier. BMC Nephrol. 2014, 15, 24. [Google Scholar] [CrossRef] [PubMed]
- Mak, A.; Kow, N.Y. Imbalance between endothelial damage and repair: A gateway to cardiovascular disease in systemic lupus erythematosus. Biomed. Res. Int. 2014, 2014, 178721. [Google Scholar] [CrossRef] [PubMed]
- Jourde-Chiche, N.; Fakhouri, F.; Dou, L.; Bellien, J.; Burtey, S.; Frimat, M.; Jarrot, P.A.; Kaplanski, G.; Le Quintrec, M.; Pernin, V.; et al. Endothelium structure and function in kidney health and disease. Nat. Rev. Nephrol. 2019, 15, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.H.; Yu, F.; Tan, Y.; Qu, Z.; Chen, M.H.; Wang, S.X.; Liu, G.; Zhao, M.H. Inclusion of renal vascular lesions in the 2003 ISN/RPS system for classifying lupus nephritis improves renal outcome predictions. Kidney Int. 2013, 83, 715–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, E.; Nagahama, K.; Shimizu, A.; Aoki, M.; Higo, S.; Yasuda, F.; Mii, A.; Fukui, M.; Kaneko, T.; Tsuruoka, S. Glomerular capillary and endothelial cell injury is associated with the formation of necrotizing and crescentic lesions in crescentic glomerulonephritis. J. Nippon. Med. Sch. 2015, 82, 27–35. [Google Scholar] [CrossRef]
- Elshal, M.; Abdelaziz, A.; Abbas, A.; Mahmoud, K.; Fathy, H.; El Mongy, S.; El-Basyuoni, S.; Ahmed, H.; McCoy, P. Quantification of circulating endothelial cells in peripheral blood of systemic lupus erythematosus patients: A simple and reproducible method of assessing endothelial injury and repair. Nephrol. Dialysis Transpl. Off. Publ. Eur. DTA ERA 2009, 24, 1495–1499. [Google Scholar] [CrossRef]
- Nakatani, K.; Fujii, H.; Hasegawa, H.; Terada, M.; Arita, N.; Ito, M.R.; Ono, M.; Takahashi, S.; Saiga, K.; Yoshimoto, S.; et al. Endothelial adhesion molecules in glomerular lesions: Association with their severity and diversity in lupus models. Kidney Int. 2004, 65, 1290–1300. [Google Scholar] [CrossRef] [Green Version]
- Gilkeson, G.S.; Mashmoushi, A.K.; Ruiz, P.; Caza, T.N.; Perl, A.; Oates, J.C. Endothelial nitric oxide synthase reduces crescentic and necrotic glomerular lesions, reactive oxygen production, and MCP1 production in murine lupus nephritis. PLoS ONE 2013, 8, e64650. [Google Scholar] [CrossRef] [PubMed]
- Berthier, C.C.; Bethunaickan, R.; Gonzalez-Rivera, T.; Nair, V.; Ramanujam, M.; Zhang, W.; Bottinger, E.P.; Segerer, S.; Lindenmeyer, M.; Cohen, C.D.; et al. Cross-species transcriptional network analysis defines shared inflammatory responses in murine and human lupus nephritis. J. Immunol. 2012, 189, 988–1001. [Google Scholar] [CrossRef] [PubMed]
- Fogo, A.B.; Kon, V. The glomerulus—A view from the inside—The endothelial cell. Int. J. Biochem. Cell Biol. 2010, 42, 1388–1397. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.; Rengarajan, T.; Thangavel, J.; Nishigaki, Y.; Sakthisekaran, D.; Sethi, G.; Nishigaki, I. The vascular endothelium and human diseases. Int. J. Biol. Sci. 2013, 9, 1057–1069. [Google Scholar] [CrossRef] [PubMed]
- Tsokos, G.C. Systemic lupus erythematosus. N. Engl. J. Med. 2011, 365, 2110–2121. [Google Scholar] [CrossRef]
- Nowling, T.K.; Gilkeson, G.S. Mechanisms of tissue injury in lupus nephritis. Arthr. Res. Ther. 2011, 13, 250. [Google Scholar] [CrossRef]
- Weening, J.J.; D’Agati, V.D.; Schwartz, M.M.; Seshan, S.V.; Alpers, C.E.; Appel, G.B.; Balow, J.E.; Bruijn, J.A.; Cook, T.; Ferrario, F.; et al. The classification of glomerulonephritis in systemic lupus erythematosus revisited. Kidney Int. 2004, 65, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Jiao, Y.; Cui, B.; Gao, X.; Xia, Y.; Zhao, Y. Immune complexes activate human endothelium involving the cell-signaling HMGB1-RAGE axis in the pathogenesis of lupus vasculitis. Lab. Investig. 2013, 93, 626–638. [Google Scholar] [CrossRef]
- Wang, L.; Law, H.K.W. Immune complexes suppressed autophagy in glomerular endothelial cells. Cell Immunol. 2018, 328, 1–8. [Google Scholar] [CrossRef]
- Pincus, Z.; Theriot, J.A. Comparison of quantitative methods for cell-shape analysis. J. Microsc. 2007, 227, 140–156. [Google Scholar] [CrossRef]
- Telikepalli, S.N.; Kumru, O.S.; Kalonia, C.; Esfandiary, R.; Joshi, S.B.; Middaugh, C.R.; Volkin, D.B. Structural characterization of IgG1 mAb aggregates and particles generated under various stress conditions. J. Pharm. Sci. 2014, 103, 796–809. [Google Scholar] [CrossRef] [PubMed]
- Filipe, V.; Poole, R.; Oladunjoye, O.; Braeckmans, K.; Jiskoot, W. Detection and characterization of subvisible aggregates of monoclonal IgG in serum. Pharm. Res. 2012, 29, 2202–2212. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Takahashi, T.; Asahara, T.; Ohura, N.; Sokabe, T.; Kamiya, A.; Ando, J. Proliferation, differentiation, and tube formation by endothelial progenitor cells in response to shear stress. J. Appl. Physiol. 2003, 95, 2081–2088. [Google Scholar] [CrossRef] [PubMed]
- Arnaoutova, I.; George, J.; Kleinman, H.K.; Benton, G. The endothelial cell tube formation assay on basement membrane turns 20: State of the science and the art. Angiogenesis 2009, 12, 267–274. [Google Scholar] [CrossRef]
- Kleinman, H.K.; Martin, G.R. Matrigel: Basement membrane matrix with biological activity. Semin. Cancer Biol. 2005, 15, 378–386. [Google Scholar] [CrossRef]
- Lin, J.R.; Shen, W.L.; Yan, C.; Gao, P.J. Downregulation of dynamin-related protein 1 contributes to impaired autophagic flux and angiogenic function in senescent endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1413–1422. [Google Scholar] [CrossRef]
- Marquez-Curtis, L.A.; Sultani, A.B.; McGann, L.E.; Elliott, J.A. Beyond membrane integrity: Assessing the functionality of human umbilical vein endothelial cells after cryopreservation. Cryobiology 2016, 72, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Denninger, J.W.; Marletta, M.A. Guanylate cyclase and the .NO/cGMP signaling pathway. Biochim. Biophys. Acta 1999, 1411, 334–350. [Google Scholar] [CrossRef]
- Armstead, V.E.; Minchenko, A.G.; Schuhl, R.A.; Hayward, R.; Nossuli, T.O.; Lefer, A.M. Regulation of P-selectin expression in human endothelial cells by nitric oxide. Am. J. Physiol. 1997, 273, H740–H746. [Google Scholar] [CrossRef]
- Schafer, A.; Wiesmann, F.; Neubauer, S.; Eigenthaler, M.; Bauersachs, J.; Channon, K.M. Rapid regulation of platelet activation in vivo by nitric oxide. Circulation 2004, 109, 1819–1822. [Google Scholar] [CrossRef]
- Oates, J.C. The biology of reactive intermediates in systemic lupus erythematosus. Autoimmunity 2010, 43, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Forstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart. J. 2012, 33, 829–837, 837a–837d. [Google Scholar] [CrossRef] [PubMed]
- Stroka, K.M.; Vaitkus, J.A.; Aranda-Espinoza, H. Endothelial cells undergo morphological, biomechanical, and dynamic changes in response to tumor necrosis factor-alpha. Eur. Biophys. J. 2012, 41, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Szczygiel, A.M.; Brzezinka, G.; Targosz-Korecka, M.; Chlopicki, S.; Szymonski, M. Elasticity changes anti-correlate with NO production for human endothelial cells stimulated with TNF-alpha. Pflugers Arch. 2012, 463, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Roca-Cusachs, P.; Alcaraz, J.; Sunyer, R.; Samitier, J.; Farre, R.; Navajas, D. Micropatterning of single endothelial cell shape reveals a tight coupling between nuclear volume in G1 and proliferation. Biophys. J. 2008, 94, 4984–4995. [Google Scholar] [CrossRef]
- Potente, M.; Gerhardt, H.; Carmeliet, P. Basic and therapeutic aspects of angiogenesis. Cell 2011, 146, 873–887. [Google Scholar] [CrossRef]
- Hsu, K.S.; Guan, B.J.; Cheng, X.; Guan, D.; Lam, M.; Hatzoglou, M.; Kao, H.Y. Translational control of PML contributes to TNFalpha-induced apoptosis of MCF7 breast cancer cells and decreased angiogenesis in HUVECs. Cell Death Differ. 2016, 23, 469–483. [Google Scholar] [CrossRef]
- Du, G.; Song, Y.; Zhang, T.; Ma, L.; Bian, N.; Chen, X.; Feng, J.; Chang, Q.; Li, Z. Simvastatin attenuates TNFalphainduced apoptosis in endothelial progenitor cells via the upregulation of SIRT1. Int. J. Mol. Med. 2014, 34, 177–182. [Google Scholar] [CrossRef]
- Vanderslice, P.; Munsch, C.L.; Rachal, E.; Erichsen, D.; Sughrue, K.M.; Truong, A.N.; Wygant, J.N.; McIntyre, B.W.; Eskin, S.G.; Tilton, R.G.; et al. Angiogenesis induced by tumor necrosis factor-agr; is mediated by alpha4 integrins. Angiogenesis 1998, 2, 265–275. [Google Scholar] [CrossRef]
- Yang, H.L.; Chang, H.C.; Lin, S.W.; Senthil Kumar, K.J.; Liao, C.H.; Wang, H.M.; Lin, K.Y.; Hseu, Y.C. Antrodia salmonea inhibits TNF-alpha-induced angiogenesis and atherogenesis in human endothelial cells through the down-regulation of NF-kappaB and up-regulation of Nrf2 signaling pathways. J. Ethnopharmacol. 2014, 151, 394–406. [Google Scholar] [CrossRef]
- Sainson, R.C.; Johnston, D.A.; Chu, H.C.; Holderfield, M.T.; Nakatsu, M.N.; Crampton, S.P.; Davis, J.; Conn, E.; Hughes, C.C. TNF primes endothelial cells for angiogenic sprouting by inducing a tip cell phenotype. Blood 2008, 111, 4997–5007. [Google Scholar] [CrossRef] [PubMed]
- Bikfalvi, A.; Bicknell, R. Recent advances in angiogenesis, anti-angiogenesis and vascular targeting. Trends Pharmacol. Sci. 2002, 23, 576–582. [Google Scholar] [CrossRef]
- Carmeliet, P.; Collen, D. Molecular basis of angiogenesis. Role of VEGF and VE-cadherin. Ann. N. Y. Acad. Sci. 2000, 902, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, U.; Augustin, H.G. Angiopoietins: A link between angiogenesis and inflammation. Trends Immunol. 2006, 27, 552–558. [Google Scholar] [CrossRef]
- Avihingsanon, Y.; Benjachat, T.; Tassanarong, A.; Sodsai, P.; Kittikovit, V.; Hirankarn, N. Decreased renal expression of vascular endothelial growth factor in lupus nephritis is associated with worse prognosis. Kidney Int. 2009, 75, 1340–1348. [Google Scholar] [CrossRef] [Green Version]
- Peterson, K.S.; Huang, J.F.; Zhu, J.; D’Agati, V.; Liu, X.; Miller, N.; Erlander, M.G.; Jackson, M.R.; Winchester, R.J. Characterization of heterogeneity in the molecular pathogenesis of lupus nephritis from transcriptional profiles of laser-captured glomeruli. J. Clinc. Investig. 2004, 113, 1722–1733. [Google Scholar] [CrossRef] [Green Version]
- Wongpiyabovorn, J.; Hirankarn, N.; Ruchusatsawat, K.; Yooyongsatit, S.; Benjachat, T.; Avihingsanon, Y. The association of single nucleotide polymorphism within vascular endothelial growth factor gene with systemic lupus erythematosus and lupus nephritis. Int. J. Immunogenet. 2011, 38, 63–67. [Google Scholar] [CrossRef]
- Bakr, S.I.; Shehab, A.A.-S.; Shennawy, D.E.; Ahmed, R.; Abo-shady; Zeitoun, Y.A.; Shedid, N.H. Serum angiopoietin-2 and soluble thrombomodulin in patients with systemic lupus erythematosus and their relation to disease activity and renal affection. J. Med. Sci. Clin. Res. 2014, 2, 14. [Google Scholar]
- El-Banawy, H.S.; Gaber, E.W.; Maharem, D.A.; Matrawy, K.A. Angiopoietin-2, endothelial dysfunction and renal involvement in patients with systemic lupus erythematosus. J. Nephrol. 2012, 25, 541–550. [Google Scholar] [CrossRef]
- Weinberg, J.B.; Granger, D.L.; Pisetsky, D.S.; Seldin, M.F.; Misukonis, M.A.; Mason, S.N.; Pippen, A.M.; Ruiz, P.; Wood, E.R.; Gilkeson, G.S. The role of nitric oxide in the pathogenesis of spontaneous murine autoimmune disease: Increased nitric oxide production and nitric oxide synthase expression in MRL-lpr/lpr mice, and reduction of spontaneous glomerulonephritis and arthritis by orally administered NG-monomethyl-L-arginine. J. Exp. Med. 1994, 179, 651–660. [Google Scholar]
- Wanchu, A.; Khullar, M.; Deodhar, S.D.; Bambery, P.; Sud, A. Nitric oxide synthesis is increased in patients with systemic lupus erythematosus. Rheumatol. Int. 1998, 18, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Belmont, H.M.; Levartovsky, D.; Goel, A.; Amin, A.; Giorno, R.; Rediske, J.; Skovron, M.L.; Abramson, S.B. Increased nitric oxide production accompanied by the up-regulation of inducible nitric oxide synthase in vascular endothelium from patients with systemic lupus erythematosus. Arthr. Rheum. 1997, 40, 1810–1816. [Google Scholar] [CrossRef] [PubMed]
- Cines, D.B.; Lyss, A.P.; Reeber, M.; Bina, M.; DeHoratius, R.J. Presence of complement-fixing anti-endothelial cell antibodies in systemic lupus erythematosus. J. Clinc. Investig. 1984, 73, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Wunder, M.B.; Norris, D.A.; Shellman, Y.G. A simple protocol for using a LDH-based cytotoxicity assay to assess the effects of death and growth inhibition at the same time. PLoS ONE 2011, 6, e26908. [Google Scholar] [CrossRef] [PubMed]
- Arnaoutova, I.; Kleinman, H.K. In vitro angiogenesis: Endothelial cell tube formation on gelled basement membrane extract. Nat. Protoc. 2010, 5, 628–635. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Law, H.K.W. Immune Complexes Impaired Glomerular Endothelial Cell Functions in Lupus Nephritis. Int. J. Mol. Sci. 2019, 20, 5281. https://doi.org/10.3390/ijms20215281
Wang L, Law HKW. Immune Complexes Impaired Glomerular Endothelial Cell Functions in Lupus Nephritis. International Journal of Molecular Sciences. 2019; 20(21):5281. https://doi.org/10.3390/ijms20215281
Chicago/Turabian StyleWang, Linlin, and Helen Ka Wai Law. 2019. "Immune Complexes Impaired Glomerular Endothelial Cell Functions in Lupus Nephritis" International Journal of Molecular Sciences 20, no. 21: 5281. https://doi.org/10.3390/ijms20215281
APA StyleWang, L., & Law, H. K. W. (2019). Immune Complexes Impaired Glomerular Endothelial Cell Functions in Lupus Nephritis. International Journal of Molecular Sciences, 20(21), 5281. https://doi.org/10.3390/ijms20215281