Screen for Potential Candidate Alternatives of Sargentodoxa cuneata from Its Six Adulterants Based on Their Phenolic Compositions and Antioxidant Activities


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Differences in Total Phenols and Flavonoids in Stems of S. cuneata and Its Six Adulterants
2.2. Differences in Antioxidant Activities in Stems of S. cuneata and Its Six Adulterants
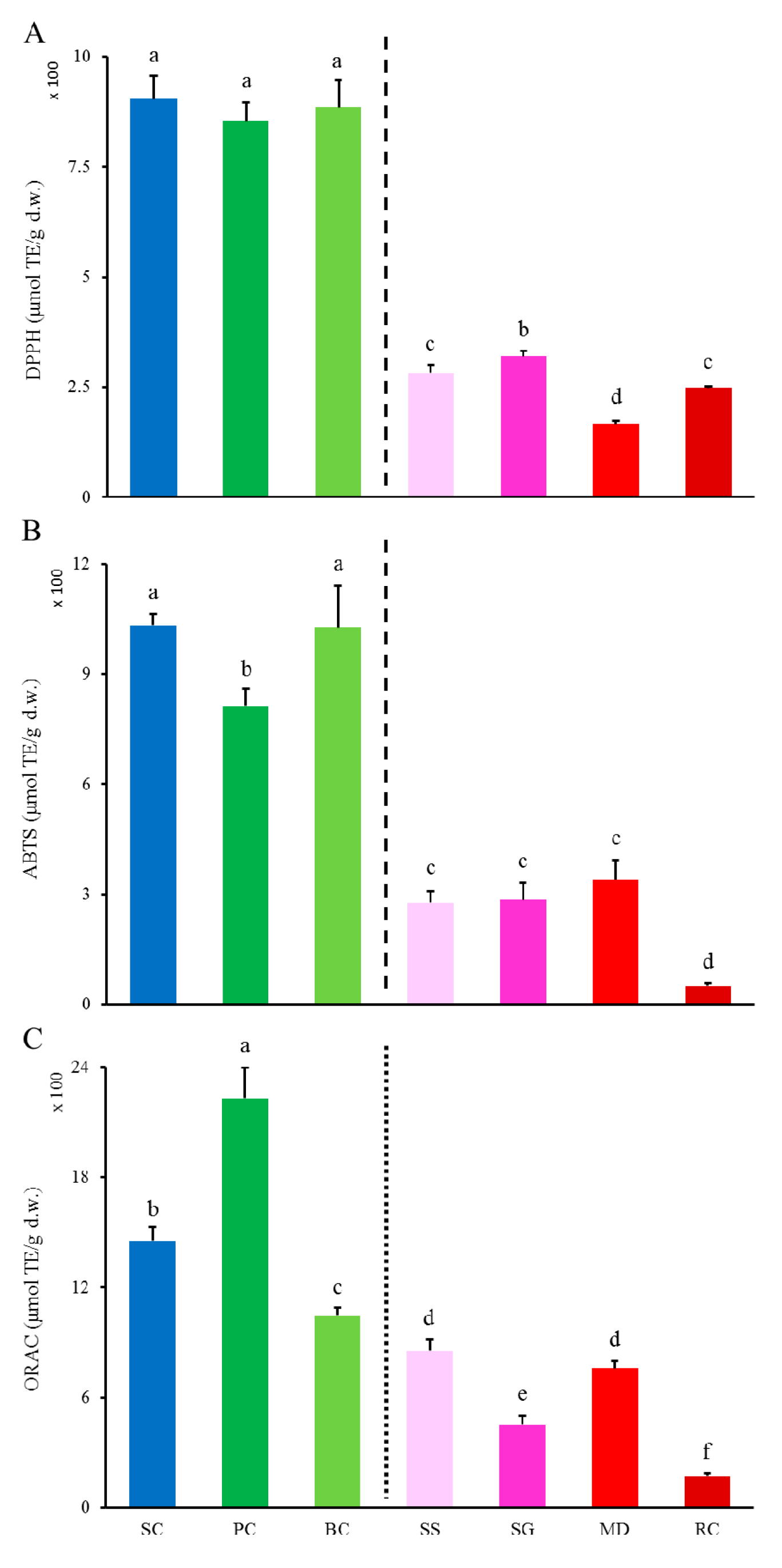
2.2.1. DPPH• and ABTS•+ Scavenging Activities and ORAC
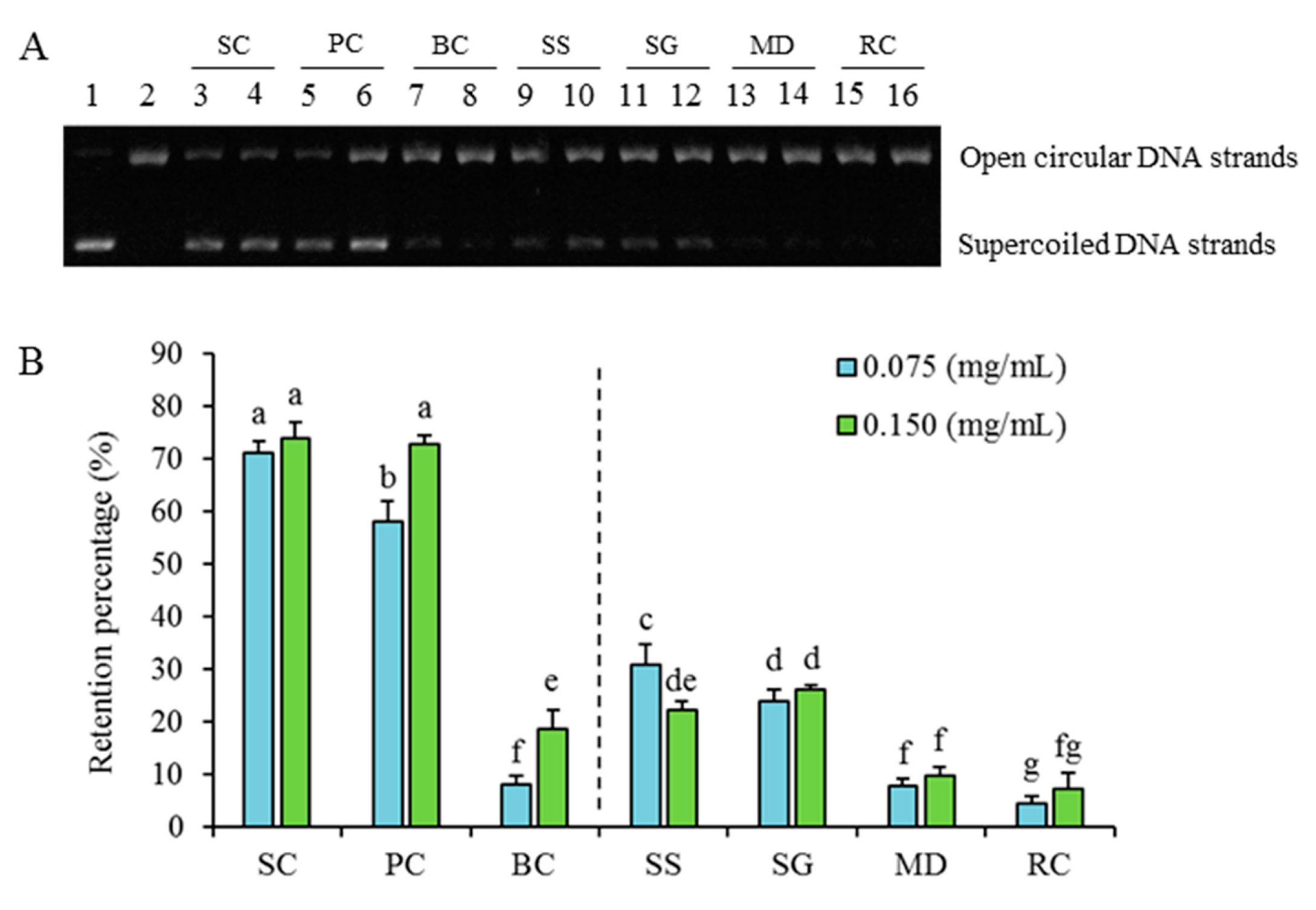
2.2.2. Differences in Inhibition against Radicals-Induced Supercoiled Plasmid DNA Strand Scission
2.3. Differences in HPLC Fingerprints of S. cuneata and Its Six Adulterants
2.4. Characterization of Phenolic Compounds by UPLC-DAD-QTOF-MS/MS in S. cuneata and Its Two Potential Candidate Alternatives
2.5. Antioxidant Contribution of Key Individual Phenolics in S. cuneata and Its Two Potential Candidate Alternatives
3. Materials and Methods
3.1. Chemicals, Reagents and Plant Materials
3.2. Extraction of Phenolics from Stems of S. cuneata and Its Six Adulterants
3.3. Measurements of Phenolics and Assessments of Antioxidant Capacities
3.3.1. Total Phenols and Total Flavonoids
3.3.2. DPPH• and ABTS•+ Scavenging Activities and ORAC
3.3.3. Inhibition of Radicals-Induced Supercoiled Plasmid DNA Strand Scission
3.4. Characterization and Quantification of Phenolic Compositions by UPLC-DAD-QTOF-MS/MS and HPLC-UV
3.5. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DPPH | 2,2′-diphenyl-1-picrylhydrazyl |
| ABTS | 2,2′-azobis-(3-ethylbenzothiazoline-6-sulfonic acid) |
| ORAC | oxygen radical absorbance capacity |
| AAPH | 2,2′-azobis (2-methylpropionamidine) dihydrochloride |
| UPLC-QTOF-MS/MS | ultra performance liquid chromatography-diode array detector-quadrupole time-of-flight-mass spectrometer/mass spectrometer |
| HPLC | high performance liquid chromatography |
| GAE | gallic acid equivalent |
| RTE | rutin equivalent |
| TE | Trolox equivalent |
| ROS | reactive oxygen species |
| SC | Sargentodoxa cuneata |
| PC | Polygonum cuspidatum |
| BC | Bauhinia championii |
| SS | Schisandra sphenanthera |
| SG | Schisandra grandiflora |
| MD | Millettia dielsiana |
| RC | Rubia cordifolia |
References
- Jiangsu New Medical College. Dictionary of Chinese Traditional Medicine; Shanghai Press of Science and Technology: Shanghai, China, 1986; pp. 268–270. [Google Scholar]
- Chen, D.Z.; Shimizu, T. Lardizabalaceae. In Flora of China; Wu, Z.Y., Ed.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2001; Volume 6, pp. 440–454. [Google Scholar]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science and Technology Press: Beijing, China, 2015; Volume 1, p. 20. [Google Scholar]
- Sakakibara, I.; Yoshida, M.; Hayashi, K.; Maruno, M. Anti-inflammatory activities of glycosides from Sargentodoxa cuneata stems. Chem. Abstr. 1997, 122. [Google Scholar]
- Wan, X.M.; Zhou, X.L.; Chen, H. Anti-thrombotic and anti-tumor effect of water extract of caulis of Sargentodoxa cuneata (Oliv) Rehd et Wils (Lardizabalaceae) in animal models. Trop. J. Pharm. Res. 2016, 15, 2391. [Google Scholar]
- Zeng, X.; Wang, H.; Gong, Z.; Huang, J.; Pei, W.; Wang, X.; Zhang, J.; Tang, X. Antimicrobial and cytotoxic phenolics and phenolic glycosides from Sargentodoxa cuneata. Fitoterapia 2015, 101, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.P.; Pang, J.; Wang, X.W.; Shen, Z.Q.; Jin, M.; Li, J.W. In vitro screening of traditionally used medicinal plants in China against enteroviruses. World J. Gastroenterol. 2006, 12, 4078–4081. [Google Scholar] [CrossRef] [PubMed]
- Krishnaiah, D.; Sarbatly, R.; Nithyanandam, R. A review of the antioxidant potential of medicinal plant species. Food Bioprod. Process. 2011, 89, 217–233. [Google Scholar] [CrossRef]
- Li, H.B.; Wong, C.C.; Cheng, K.W.; Chen, F. Antioxidant properties in vitro and total phenolic contents in methanol extracts from medicinal plants. LWT 2008, 41, 385–390. [Google Scholar] [CrossRef]
- Barbour, V.; Clark, J.; Jones, S.; Norton, M.; Simpson, P.; Veitch, E. Why drug safety should not take a back seat to efficacy. PLoS Med. 2011, 8, e1001097. [Google Scholar]
- Chan, T.Y.K. Herbal Medicines Induced Anticholinergic Poisoning in Hong Kong. Toxins 2016, 8, 80. [Google Scholar] [CrossRef]
- Sun, Z.X.; Ye, L.J.; Zhang, F.; Hu, W.; Fan, D.M.; Zhang, Z.Y. Development of microsatellite markers for Sargentodoxa cuneata (Lardizabalaceae) using next-generation sequencing technology1. Appl. Plant Sci. 2016, 4, 1600003. [Google Scholar] [CrossRef]
- Zheng, X.; Zheng, W.; Zhou, J.; Gao, X.; Liu, Z.; Han, N.; Yin, J. Study on the discrimination between Corydalis Rhizoma and its adulterants based on HPLC-DAD-Q-TOF-MS associated with chemometric analysis. J. Chromatogr. B 2018, 1090, 110–121. [Google Scholar] [CrossRef]
- Heo, S.; Choi, J.Y.; Yoo, G.J.; Park, S.K.; Baek, S.Y. Simultaneous analysis of 35 specific antihypertensive adulterants in dietary supplements using LC/MS/MS. Biomed. Chromatogr. 2017, 31, e3856. [Google Scholar] [CrossRef] [PubMed]
- Paíga, P.; Rodrigues, M.J.; Correia, M.; Amaral, J.S.; Oliveira, M.B.P.; Delerue-Matos, C. Analysis of pharmaceutical adulterants in plant food supplements by UHPLC-MS/MS. Eur. J. Pharm. Sci. 2017, 99, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Liu, Y.; Wang, X.; Song, J.; Chen, S.; Ragupathy, S.; Han, J.; Newmaster, S.G. Derivative Technology of DNA Barcoding (Nucleotide Signature and SNP Double Peak Methods) Detects Adulterants and Substitution in Chinese Patent Medicines. Sci. Rep. 2017, 7, 5858. [Google Scholar] [CrossRef]
- Han, J.; Pang, X.; Liao, B.; Yao, H.; Song, J.; Chen, S. An authenticity survey of herbal medicines from markets in China using DNA barcoding. Sci. Rep. 2016, 6, 18723. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.X.; Liu, D.L.; Gao, W.Y.; Zhang, T.J. A new macrolide and glycosides from the stem of Sargentodoxa cuneata. Chin. Chem. Lett. 2009, 20, 1339–1341. [Google Scholar] [CrossRef]
- Tang, J.; Ma, R.L.; Ouyang, Z.; Chen, H.S. Chemical constituents from the water-soluble fraction of wild Sargentodoxa cuneata. Chin. J. Nat. Med. 2012, 10, 115–118. [Google Scholar] [CrossRef]
- Miao, K.; Zhang, J.; Wang, F.; Qin, Y. Studies on the chemical constituents of Sargentgloryvine (Sargentodoxa cuneata). Chin. Tradit. Herb. Drugs 1995, 26, 171–173. [Google Scholar]
- Zhang, P.; Yan, S.; Shao, Y.; Li, Z. Effect of some water soluble substances of Sargentodoxa cuneata on myocardial ischemia. J. Shanghai Med. 1988, 15, 191–194. [Google Scholar]
- Chang, J.; Case, R. Phenolic glycosides and ionone glycoside from the stem of Sargentodoxa cuneata. Phytochemistry 2005, 66, 2752–2758. [Google Scholar] [CrossRef]
- Zhang, B.; Deng, Z.; Tang, Y.; Chen, P.X.; Liu, R.; Ramdath, D.D.; Liu, Q.; Hernandez, M.; Tsao, R. Reprint of “Bioaccessibility, in vitro antioxidant and anti-inflammatory activities of phenolics in cooked green lentil (Lens culinaris)”. J. Funct. Foods 2017, 38, 698–705. [Google Scholar] [CrossRef]
- Sumczynski, D.; Kotásková, E.; Orsavová, J.; Valášek, P. Contribution of individual phenolics to antioxidant activity and in vitro digestibility of wild rices (Zizania aquatica L.). Food Chem. 2017, 218, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazúr, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Boil. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.Y.; Zhou, B.; Cai, Y.J.; Yang, L.; Liu, Z.L. Synergistic effect of green tea polyphenols with trolox on free radical-induced oxidative DNA damage. Food Chem. 2006, 96, 90–95. [Google Scholar] [CrossRef]
- Heras, R.M.L.; Rada, P.Q.; Andrés, A.; Lamuela-Raventós, R. Polyphenolic profile of persimmon leaves by high resolution mass spectrometry (LC-ESI-LTQ-Orbitrap-MS). J. Funct. Foods 2016, 23, 370–377. [Google Scholar] [CrossRef]
- Hu, X.; Chen, L.; Shi, S.; Cai, P.; Liang, X.; Zhang, S. Antioxidant capacity and phenolic compounds of Lonicerae macranthoides by HPLC–DAD–QTOF-MS/MS. J. Pharm. Biomed. Anal. 2016, 124, 254–260. [Google Scholar] [CrossRef]
- Liu, A.H.; Lin, Y.H.; Yang, M.; Guo, H.; Guan, S.H.; Sun, J.H.; Guo, D.A. Development of the fingerprints for the quality of the roots of Salvia miltiorrhiza and its related preparations by HPLC-DAD and LC-MS(n). J. Chromatogr. B 2007, 846, 32–41. [Google Scholar] [CrossRef]
- Cádiz-Gurrea, M.D.L.L.; Fernández-Arroyo, S.; Joven, J.; Carretero, A.S. Comprehensive characterization by UHPLC-ESI-Q-TOF-MS from an Eryngium bourgatii extract and their antioxidant and anti-inflammatory activities. Food Res. Int. 2013, 50, 197–204. [Google Scholar] [CrossRef]
- Yang, L.; Yin, P.; Fan, H.; Xue, Q.; Li, K.; Li, X.; Sun, L.; Liu, Y. Response Surface Methodology Optimization of Ultrasonic-Assisted Extraction of Acer Truncatum Leaves for Maximal Phenolic Yield and Antioxidant Activity. Molecules 2017, 22, 232. [Google Scholar] [CrossRef]
- Ramos, P.A.B.; Santos, S.A.; Guerra, Â.R.; Guerreiro, O.; Felício, L.; Jerónimo, E.; Silvestre, A.J.; Neto, C.P.; Duarte, M. Valorization of olive mill residues: Antioxidant and breast cancer antiproliferative activities of hydroxytyrosol-rich extracts derived from olive oil by-products. Ind. Crops Prod. 2013, 46, 359–368. [Google Scholar] [CrossRef]
- Han, F.; Li, Y.; Ma, L.; Liu, T.; Wu, Y.; Xu, R.; Song, A.; Yin, R. A rapid and sensitive UHPLC-FT-ICR MS/MS method for identification of chemical constituents in Rhodiola crenulata extract, rat plasma and rat brain after oral administration. Talanta 2016, 160, 183–193. [Google Scholar] [CrossRef]
- Lin, L.; Harnly, J.M. LC-MS profiling and quantification of food phenolic components using a standard analytical approach for all plants. In Food Science and Technology: New Research; Greco, L.V., Bruno, M.N., Eds.; Nova Publisher: New York, NY, USA, 2008. [Google Scholar]
- Melguizo-Melguizo, D.; Díaz-De-Cerio, E.; Quirantes-Piné, R.; Švarc-Gajić, J.; Carretero, A.S. The potential of Artemisia vulgaris leaves as a source of antioxidant phenolic compounds. J. Funct. Foods 2014, 10, 192–200. [Google Scholar] [CrossRef]
- Pollio, A.; Zarrelli, A.; Romanucci, V.; Di Mauro, A.; Barra, F.; Pinto, G.; Crescenzi, E.; Roscetto, E.; Palumbo, G. Polyphenolic Profile and Targeted Bioactivity of Methanolic Extracts from Mediterranean Ethnomedicinal Plants on Human Cancer Cell Lines. Molecules 2016, 21, 395. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ding, L.; Jin, H.; Shi, R.; Li, Y.; Wu, J.; Li, Y.; Zhu, L.; Ma, Y. Simultaneous quantification of catechin, epicatechin, liquiritin, isoliquiritin, liquiritigenin, isoliquiritigenin, piperine, and glycyrrhetinic acid in rat plasma by HPLC-MS/MS: Application to a pharmacokinetic study of Longhu Rendan pills. Biomed. Chromatogr. 2016, 30, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Shan, B.; Cai, Y.Z.; Brooks, J.D.; Corke, H. Antibacterial properties of Polygonum cuspidatum roots and their major bioactive constituents. Food Chem. 2008, 109, 530–537. [Google Scholar] [CrossRef]
- Furuuchi, R.; Yokoyama, T.; Watanabe, Y.; Hirayama, M. Identification and Quantification of Short Oligomeric Proanthocyanidins and Other Polyphenols in Boysenberry Seeds and Juice. J. Agric. Food Chem. 2011, 59, 3738–3746. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, A.; DellaGreca, M.; D’Abrosca, B.; Oriano, P.; Golino, A.; Izzo, A.; Zarrelli, A.; Monaco, P. Lignans, neolignans and sesquilignans from Cestrum parqui l’Her. Biochem. Syst. Ecol. 2007, 35, 392–396. [Google Scholar] [CrossRef]
- Huang, X.; Mazza, G. Simultaneous analysis of serotonin, melatonin, piceid and resveratrol in fruits using liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2011, 1218, 3890–3899. [Google Scholar] [CrossRef]
- Yao, S.; Zhang, J.; Wang, D.; Hou, J.; Yang, W.; Da, J.; Cai, L.; Yang, M.; Jiang, B.; Liu, X.; et al. Discriminatory components retracing strategy for monitoring the preparation procedure of Chinese patent medicines by fingerprint and chemometric analysis. PLoS ONE 2015, 10, e0121366. [Google Scholar] [CrossRef]
- Metlin. Available online: https://metlin.scripps.edu/ (accessed on 15 July 2019).
- Tabart, J.; Kevers, C.; Pincemail, J.; Defraigne, J.O.; Dommes, J. Comparative antioxidant capacities of phenolic compounds measured by various tests. Food Chem. 2009, 113, 1226–1233. [Google Scholar] [CrossRef]
- Ciesla, L.M.; Wojtunik-Kulesza, K.A.; Oniszczuk, A.; Waksmundzka-Hajnos, M.; Wojtunik-Kulesza, K.A.; Waksmundzka-Hajnos, M. Antioxidant synergism and antagonism between selected monoterpenes using the 2,2-diphenyl-1-picrylhydrazyl method. Flavour Fragr. J. 2016, 31, 412–419. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. In Methods in Enzymology; Academic Press: New York, NY, USA, 1955; Volume 1, p. 138. [Google Scholar]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal Processing Enhances the Nutritional Value of Tomatoes by Increasing Total Antioxidant Activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yin, P.; Li, K.; Fan, H.; Xue, Q.; Li, X.; Sun, L.; Liu, Y. Seasonal dynamics of constitutive levels of phenolic components lead to alterations of antioxidant capacities in Acer truncatum leaves. Arab. J. Chem. 2018, 11, 14–25. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Boil. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Sun, L.; Isaak, C.K.; Zhou, Y.; Petkau, J.C.; Karmin, O.; Liu, Y.; Siow, Y.L. Salidroside and tyrosol from Rhodiola protect H9c2 cells from ischemia/reperfusion-induced apoptosis. Life Sci. 2012, 91, 151–158. [Google Scholar] [CrossRef]
- Chandrasekara, A.; Shahidi, F. Antiproliferative potential and DNA scission inhibitory activity of phenolics from whole millet grains. J. Funct. Foods 2011, 3, 159–170. [Google Scholar] [CrossRef]
- Liyana-Pathirana, C.M.; Shahidi, F. Importance of Insoluble-Bound Phenolics to Antioxidant Properties of Wheat. J. Agric. Food Chem. 2006, 54, 1256–1264. [Google Scholar] [CrossRef]
- Hiramoto, K.; Ojima, N.; Sako, K.I.; Kikugawa, K. Effect of Plant Phenolics on the Formation of the Spin-Adduct of Hydroxyl Radical and the DNA Strand Breaking by Hydroxyl Radical. Boil. Pharm. Bull. 1996, 19, 558–563. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | λmax | [M−H]− (m/z) | Error (ppm) | Formula | MS/MS Fragments (m/z) | Identified Compounds | Compound Group | Samples | References |
|---|---|---|---|---|---|---|---|---|---|
| (nm) | |||||||||
| 1 | 230, 250, 294 | 153 | −3.9 | C7H5O4 | 109 | Protocatechuic acid | Hydroxybenzoic acids | SC | [29] |
| 2 | 255, 286 | 167 | −2.4 | C8H7O4 | 108, 123, 152 | Hydroxybenzoic acids | SC | [27] | |
| 3 | 262 | 197 | −9.6 | C9H9O5 | 182, 166, 153 | Syringic acid | Hydroxybenzoic acids | SC | [30] |
| 4 | 228, 272 | 169 | 3.5 | C7H5O5 | 125 | Gallic acid | Hydroxybenzoic acids | PC | [31] * |
| 5 | 230, 280 | 315 | −1.6 | C14H19O8 | 153, 135 | Hydroxytyrosol-1-O-glucoside | Tyrosols | SC | [32] |
| 6 | 228, 276 | 299 | 2 | C14H19O7 | 137, 119, 101, 96 | Salidroside | Tyrosols | SC | [43] * |
| 7 | 242, 326 | 353 | 1.1 | C16H17O9 | 191 | 3-O-Caffeoylquinic acid | Caffeoylquinic acids | SC | [35] |
| 8 | 247, 316 | 353 | −0.8 | C16H17O9 | 191, 78, 96 | 5-O-Caffeoylquinic acid | Caffeoylquinic acids | SC | [35] * |
| 9 | 235, 280 | 289 | −0.3 | C15H13O6 | 137, 109 | Catechin | Flavanols | PC BC | [37] |
| 10 | 234, 278 | 289 | −0.7 | C15H13O6 | 137, 109 | Epicatechin | Flavanols | SC PC BC | [37] * |
| 11 | 239, 280 | 441 | −1.8 | C22H17O10 | 289, 169 | Catechin-gallate | Flavanols | PC | [38] |
| 12 | 236, 278 | 577 | 1.2 | C30H25O12 | 407, 289, 425 | B type proanthocyanidin dimer 1 | Flavanol oligomer | SC BC | [37] * |
| 13 | 237, 279 | 577 | 5.5 | C30H25O12 | 289, 407 | B type proanthocyanidin dimer 2 | Flavanol oligomers | BC | [43] |
| 14 | 243, 279 | 577 | −5 | C30H25O12 | 287, 289 | B type proanthocyanidin dimer 3 | Flavanol oligomers | BC | [43] * |
| 15 | 239, 278 | 865 | 2.1 | C45H37O18 | 577, 695, 407, 287 | B type proanthocyanidin trimer | Flavanol oligomers | SC BC | [38] |
| 16 | 238, 276 | 579 | −2.8 | C28H35O13 | 417, 181, 96 | Syringaresinol-4-O-glucoside | Lignans | SC | [22] |
| 17 | 242, 280 | 581 | 10.3 | C28H37O13 | 419 | Lyoniresinol glucoside | Lignans | BC | [43] |
| 18 | 245, 283, 330 | 359 | −4.2 | C18H15O8 | 359, 197, 161, 135 | Rosmarinate | Hydroxycinnamic acids | SC | [29] * |
| 19 | 245, 283 | 493 | −4.1 | C26H21O10 | 295 | Salvianolic acid A | Hydroxycinnamic acids | SC | [29] * |
| 20 | 244, 285 | 717 | −1.8 | C36H29O16 | 519, 321 | Salvianolic acid B | Hydroxycinnamic acids | SC | [29] * |
| 21 | 249, 300 | 469 | 8.3 | C20H21O11S | 227, 215 | Resveratrol-sulfoglucoside | Stilbenes | PC | [43] |
| 22 | 249, 304 | 389 | 3.1 | C20H21O8 | 227 | Resveratrol-4′-glucoside (Resveratroloside) | Stilbenes | PC | [38] |
| 23 | 245, 280 | 469 | 10 | C20H21O11S | 227, 407, 289 | Resveratrol-sulfoglucoside (isomer) | Stilbenes | PC | [43] |
| 24 | 236, 319 | 389 | 6.4 | C20H21O8 | 227 | Resveratrol-3-O-glucoside (Piceid) | Stilbenes | PC | [38] |
| 25 | 246, 300 | 541 | 3 | C27H25O12 | 313, 169, 227 | Resveratrol-galloylglucoside | Stilbenes | PC | [38] |
| 26 | 237, 306 | 227 | −3.5 | C7H15O8 | 185, 143 | Resveratrol | Stilbenes | PC | [41] * |
| 27 | 254, 282, 426 | 431 | −2.6 | C21H19O10 | 269 | Emodin-8-O-glucoside | Anthraquinones | PC | [38] * |
| 28 | 229, 281, 427 | 431 | 2.3 | C21H19O10 | 269 | Emodin-1-O-glucoside | Anthraquinones | PC | [38] * |
| 29 | 248, 280, 427 | 517 | −2.7 | C24H21O13 | 269, 473 | Emodin-8-O-(6′-O-malonyl)-glucoside | Anthraquinones | PC | [38] |
| 30 | 247, 272, 421 | 283 | 4.2 | C16H11O5 | 268, 240 | Physcion | Anthraquinones | PC | [38] |
| 31 | 250, 268, 441 | 285 | 4.6 | C15H9O6 | 267, 257, 255 | Citreorosein | Anthraquinones | PC | [43] |
| 32 | 249, 273, 441 | 299 | 0.7 | C15H7O7 | 255, 227 | Emodic acid | Anthraquinones | PC | [43] |
| 33 | 267, 288, 441 | 269 | 3 | C19H9O2 | 225, 241 | Emodin | Anthraquinones | PC | [38] * |
| 34 | 242, 279 | 417 | 10.1 | C21H21O9 | 255 | Liquiritin | Flavanones | BC | [37] |
| 35 | 242, 279 | 417 | 4.1 | C21H21O9 | 255 | Neoliquiritin | Flavanones | BC | [37] * |
| 36 | 244, 279 | 417 | −1.9 | C21H21O9 | 255 | Neoisoliquiritin | Flavanones | BC | [35] |
| 37 | 244, 279 | 255 | 5.9 | C15H11O4 | 119 | Liquiritigenin | Flavanones | BC | [37] * |
| 38 | 247, 279 | 255 | 4.7 | C15H11O4 | 119 | Isoliquiritigenin | Flavanones | BC | [37] * |
| Categories | Antioxidant Capacities * | Semi-Contents # | Contributions (%) to | |||||
|---|---|---|---|---|---|---|---|---|
| DPPH | ABTS | ORAC | DPPH | ABTS | ORAC | |||
| in SC | ||||||||
| gallic acid | Hydroxybenzoic acids | 3.98 ± 0.38 a | 6.08 ± 0.36 a | 2.23 ± 0.03 f | 9.19 ± 0.12 | 23.81 ± 3.49 | 31.88 ± 0.14 | 8.29 ± 0.01 |
| tyrosol | Tyrosols | 0.06 ± 0.02 f | 0.21 ± 0.04 e | 1.96 ± 0.03 g | 116.44 ± 0.44 | 5.60 ± 2.33 | 17.18 ± 0.00 | 113.63 ± 0.01 |
| chlorogenic acid | Caffeoylquinic acids | 0.69 ± 0.02 c | 0.41 ± 0.05 c | 5.01 ± 0.17 b | 112.04 ± 0.87 | 24.16 ± 2.24 | 12.58 ± 0.00 | 108.95 ± 0.01 |
| epicatechin | Flavanols | 1.37 ± 0.11 b | 0.41 ± 0.03 c | 4.45 ± 0.02 c | 21.47 ± 0.21 | 11.22 ± 2.36 | 2.94 ± 0.00 | 22.64 ± 0.00 |
| proanthocyanidin B2 | Flavanol oligomers | 0.10 ± 0.00 e | 0.16 ± 0.01 f | 6.93 ± 0.05 a | 22.24 ± 0.41 | 0.43 ± 0.00 | 0.60 ± 0.00 | 18.32 ± 0.00 |
| rosmarinic acid | Hydroxycinnamic acids | 0.65 ± 0.10 c | 0.53 ± 0.05 b | 4.80 ± 0.06 b | 26.32 ± 0.26 | 5.26 ± 2.63 | 3.76 ± 0.00 | 24.12 ± 0.00 |
| Sum of contributions | 70.48 ± 2.08 | 68.95 ± 0.02 | 295.94 ± 0.01 | |||||
| in PC | ||||||||
| gallic acid | Hydroxybenzoic acids | 3.98 ± 0.38 a | 6.08 ± 0.36 a | 2.23 ± 0.03 f | 6.8 ± 0.14 | 18.64 ± 2.58 | 29.91 ± 0.14 | 4.00 ± 0.01 |
| epicatechin | Flavanols | 1.37 ± 0.11 b | 0.41 ± 0.03 c | 4.45 ± 0.02 c | 62.62 ± 0.35 | 34.65 ± 6.89 | 10.89 ± 0.00 | 43.16 ± 0.00 |
| resveratrol | Stilbenes | 0.44 ± 0.01 d | 0.28 ± 0.11 d | 2.43 ± 0.03 e | 152.66 ± 1.18 | 34.51 ± 1.53 | 23.07 ± 0.00 | 73.07 ± 0.00 |
| emodin | Anthraquinones | ND h | 0.22 ± 0.04 e | 2.50 ± 0.10 e | 185.11 ± 0.62 | 0.00 ± 0.00 | 1.69 ± 0.00 | 76.98 ± 0.00 |
| Sum of contributions | 87.80 ± 2.75 | 65.56 ± 0.04 | 197.22 ± 0.01 | |||||
| in BC | ||||||||
| epicatechin | Flavanols | 1.37 ± 0.11 b | 0.41 ± 0.03 c | 4.45 ± 0.02 c | 112.39 ± 0.76 | 59.97 ± 12.36 | 15.49 ± 0.00 | 164.40 ± 0.00 |
| proanthocyanidin B2 | Flavanol oligomers | 0.10 ± 0.00 e | 0.16 ± 0.01 f | 6.93 ± 0.05 a | 212.98 ± 1.52 | 4.16 ± 0.00 | 5.75 ± 0.00 | 243.42 ± 0.00 |
| liquiritigenin | Flavanones | 0.02 ± 0.00 g | 0.01 ± 0.00 g | 2.89 ± 0.05 d | 21.01 ± 0.08 | 0.19 ± 0.00 | 0.08 ± 0.00 | 22.61 ± 0.00 |
| Sum of contributions | 64.32 ± 4.12 | 21.32 ± 0.00 | 430.42 ± 0.00 | |||||
| Abbr. | Species | Collection Site | Collection Date | Material Conditions |
|---|---|---|---|---|
| SC | Sargentodoxa cuneata (Oliv.) Rehd. et Wils. | Beijing | 2015.7.26 | dry slices |
| PC | Polygonum cuspidatum Sieb.et Zucc. | Anguo, Heibei | 2015.8.30 | dry slices |
| BC | Bauhinia championii (Benth.) Benth. | Anguo, Heibei | 2015.8.30 | dry slices |
| SS | Schisandra sphenanthera Rehd. et Wils. | Anguo, Heibei | 2015.8.30 | dry slices |
| SG | Schisandra grandiflora (Wall.) Hook. f. et Thoms | Anguo, Heibei | 2015.8.30 | dry slices |
| MD | Millettia dielsiana Harms. | Anguo, Heibei | 2015.8.30 | dry slices |
| RC | Rubia cordifolia L. | Anguo, Heibei | 2015.8.30 | dry slices |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Yin, P.; Cao, X.; Liu, Y. Screen for Potential Candidate Alternatives of Sargentodoxa cuneata from Its Six Adulterants Based on Their Phenolic Compositions and Antioxidant Activities. Int. J. Mol. Sci. 2019, 20, 5427. https://doi.org/10.3390/ijms20215427
Yang L, Yin P, Cao X, Liu Y. Screen for Potential Candidate Alternatives of Sargentodoxa cuneata from Its Six Adulterants Based on Their Phenolic Compositions and Antioxidant Activities. International Journal of Molecular Sciences. 2019; 20(21):5427. https://doi.org/10.3390/ijms20215427
Chicago/Turabian StyleYang, Lingguang, Peipei Yin, Xinxin Cao, and Yujun Liu. 2019. "Screen for Potential Candidate Alternatives of Sargentodoxa cuneata from Its Six Adulterants Based on Their Phenolic Compositions and Antioxidant Activities" International Journal of Molecular Sciences 20, no. 21: 5427. https://doi.org/10.3390/ijms20215427