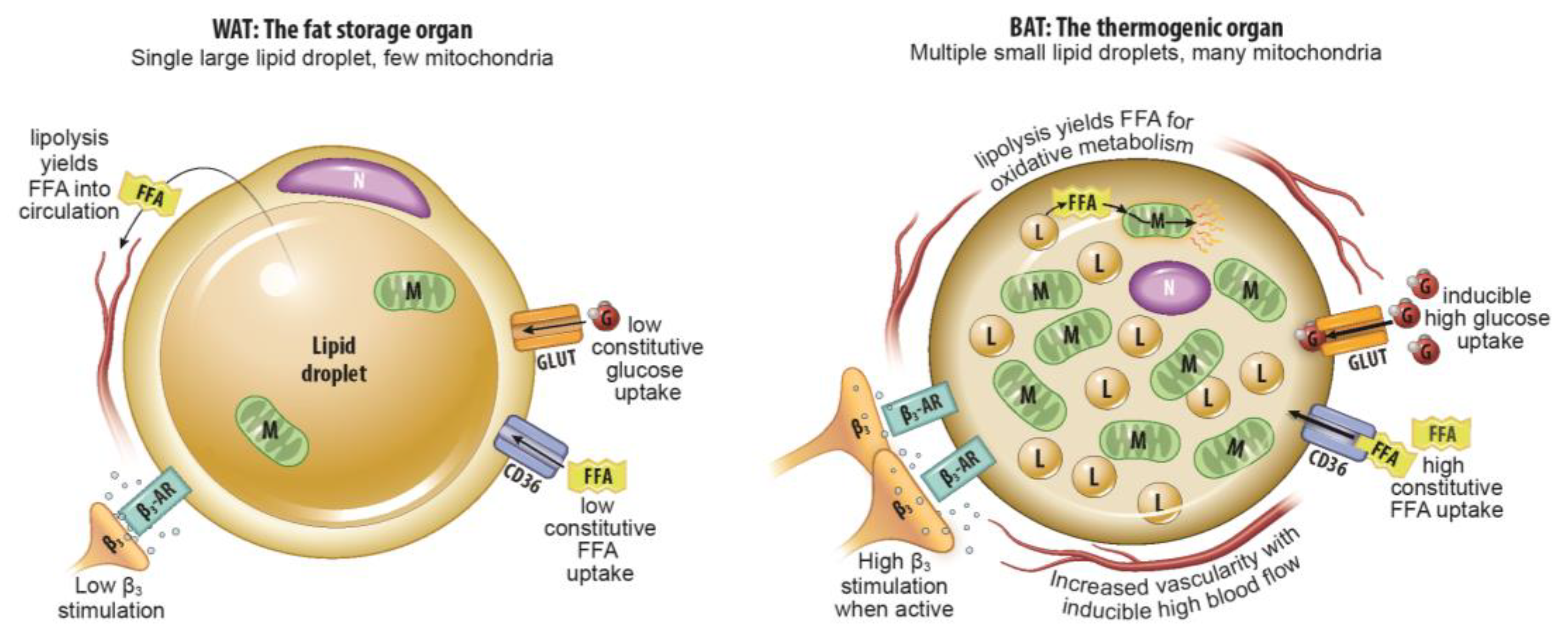
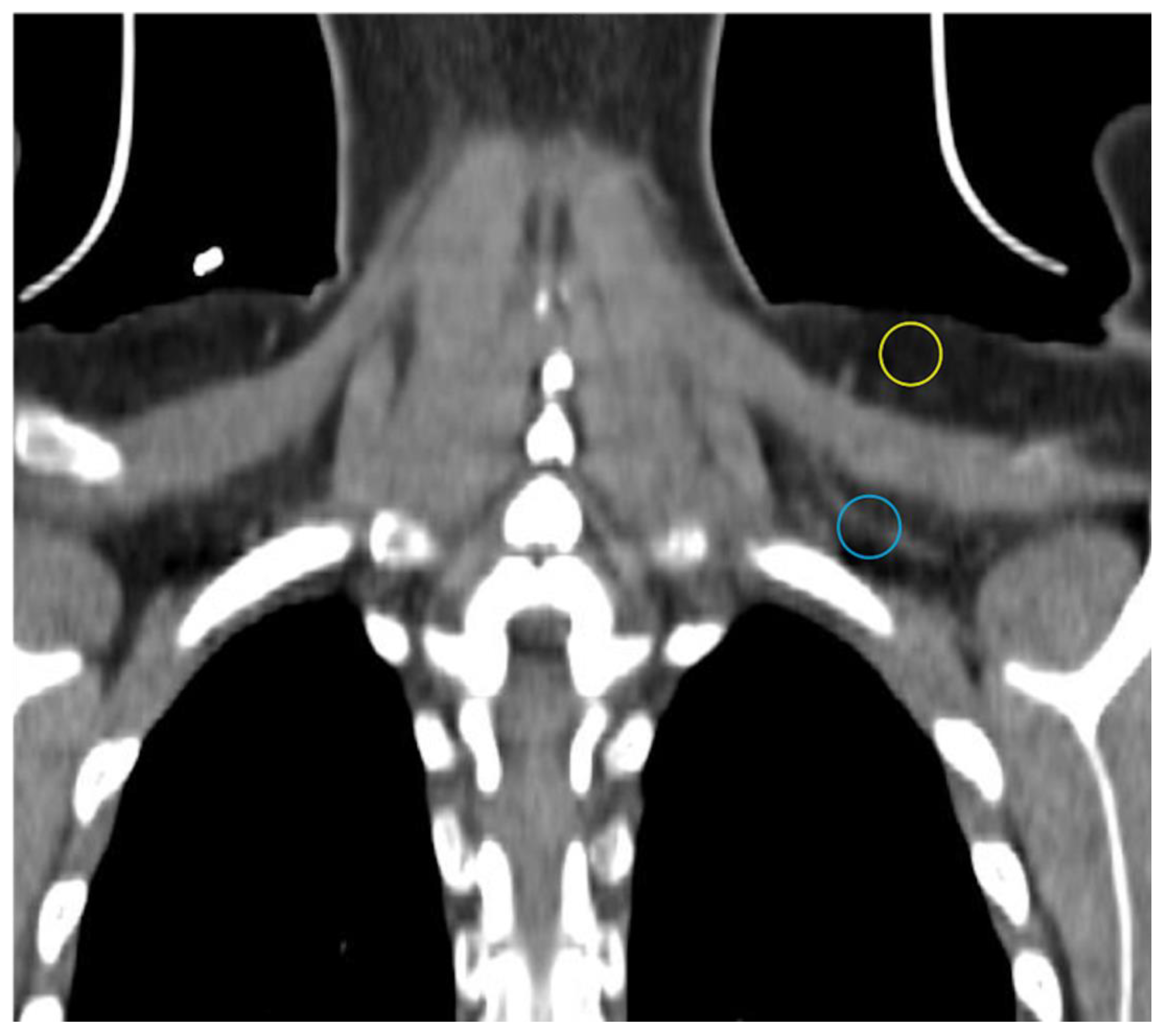
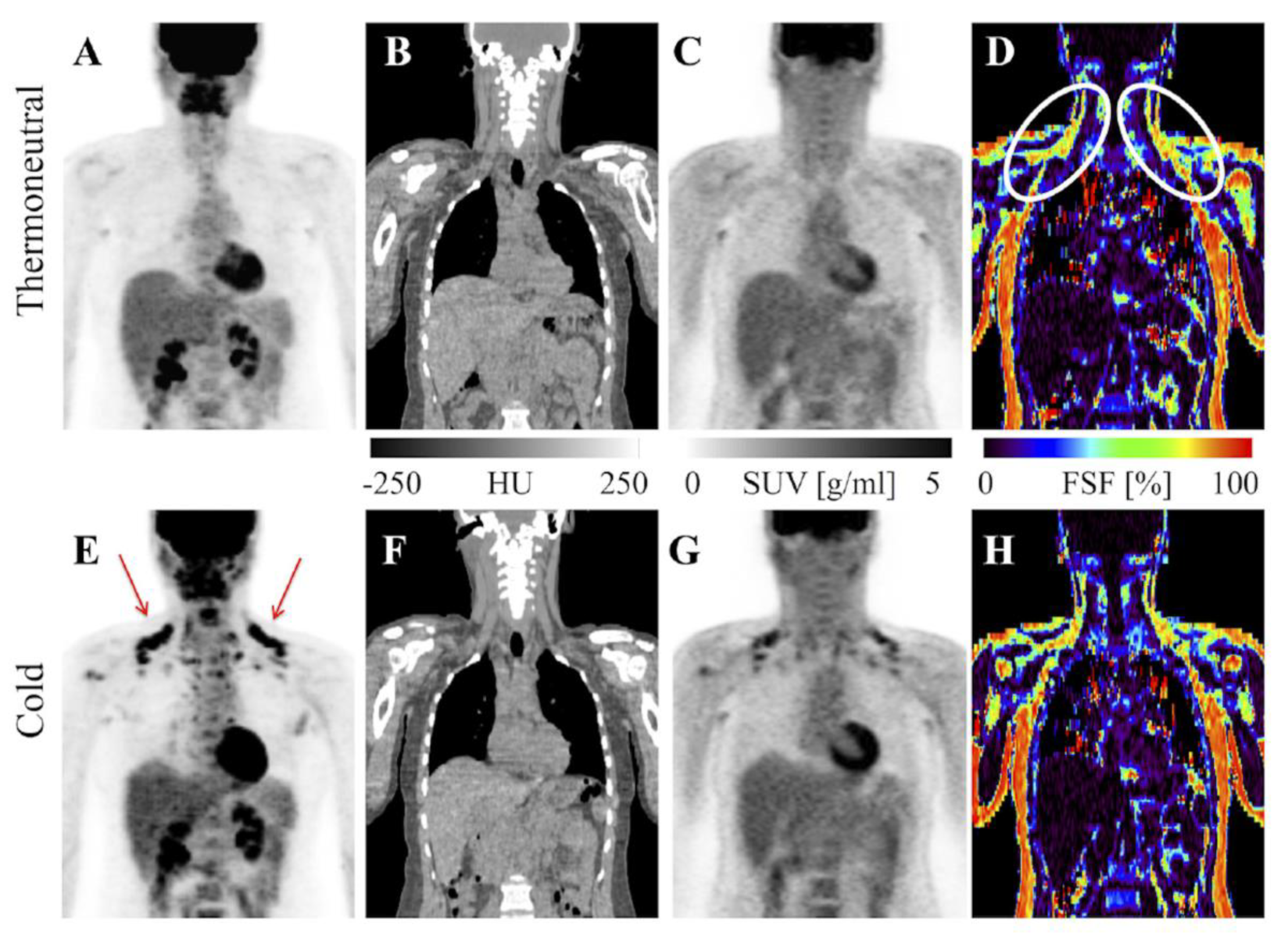
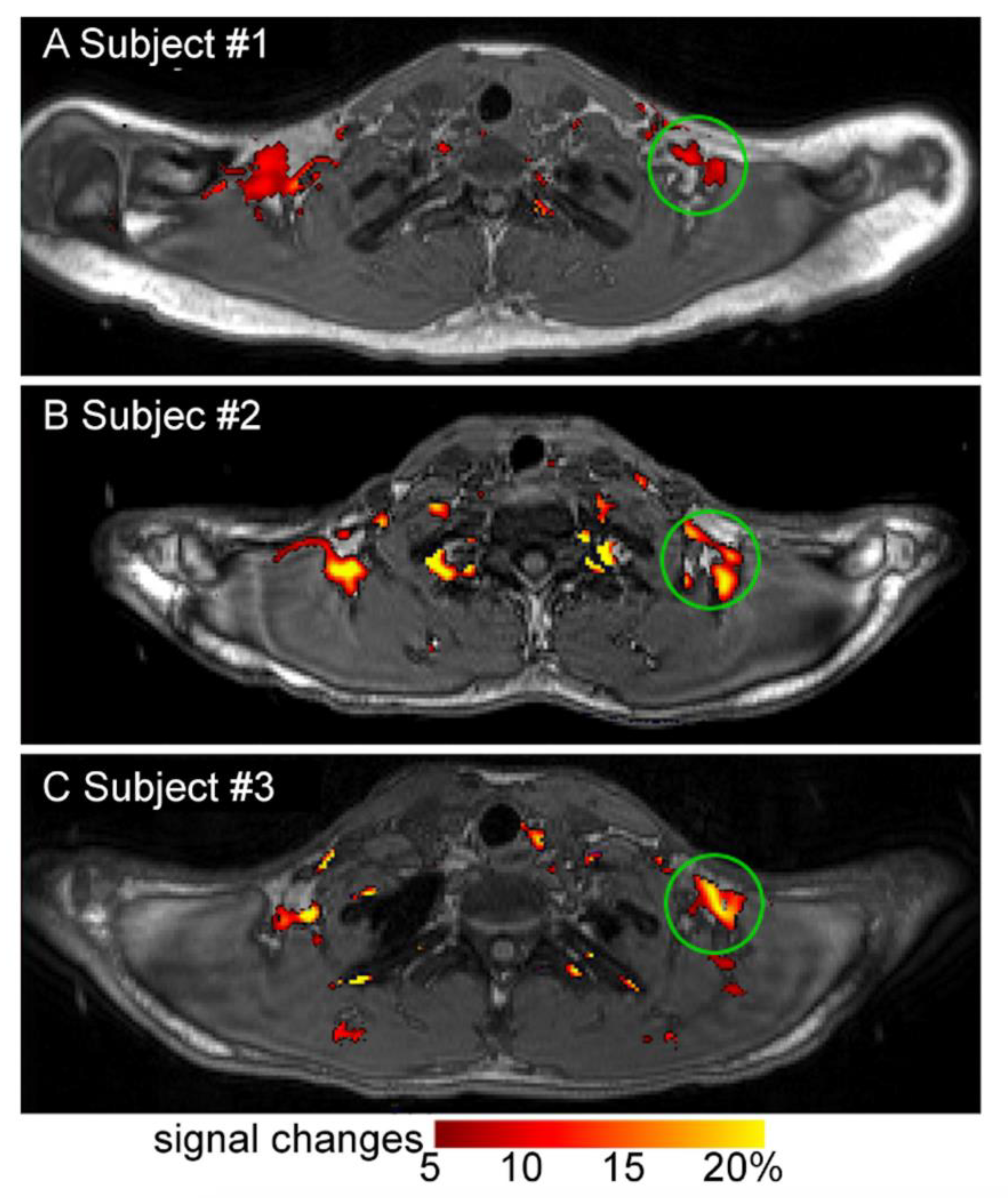
Imaging Metabolically Active Fat: A Literature Review and Mechanistic Insights


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
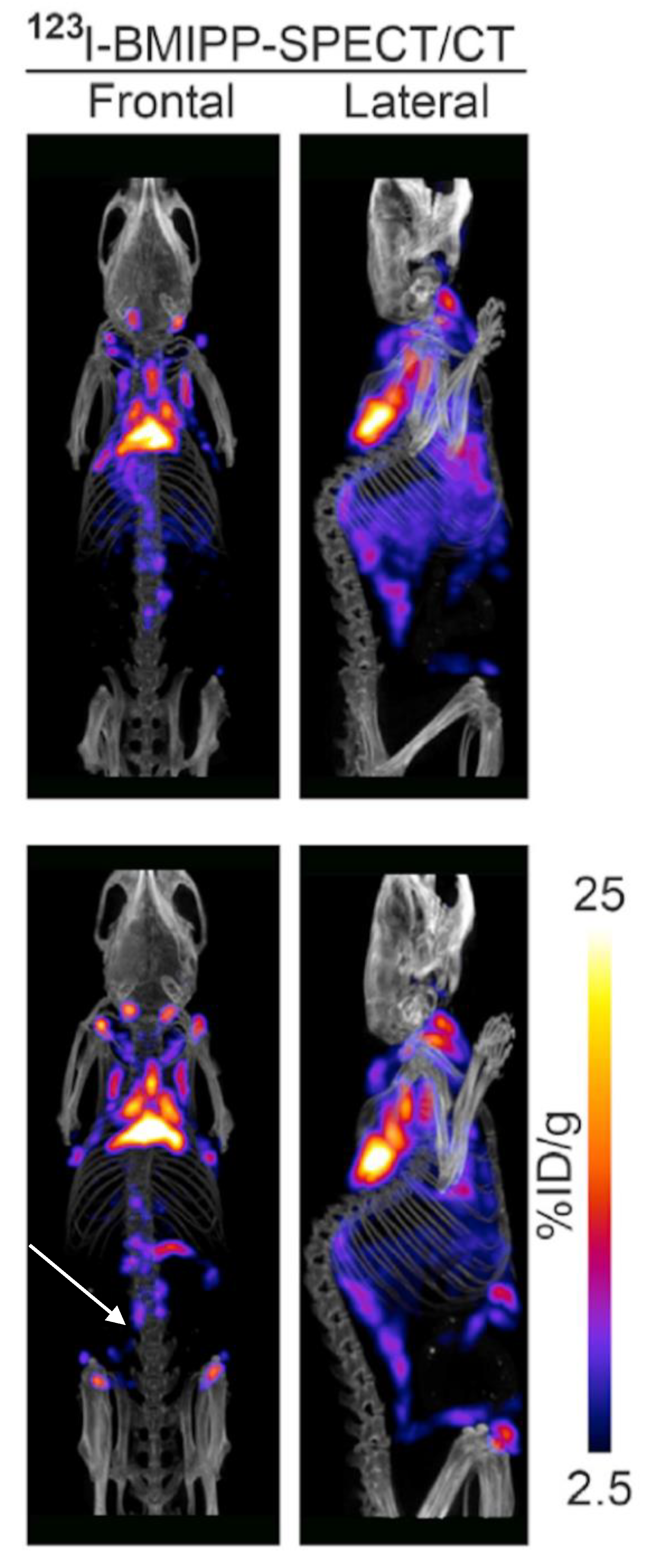
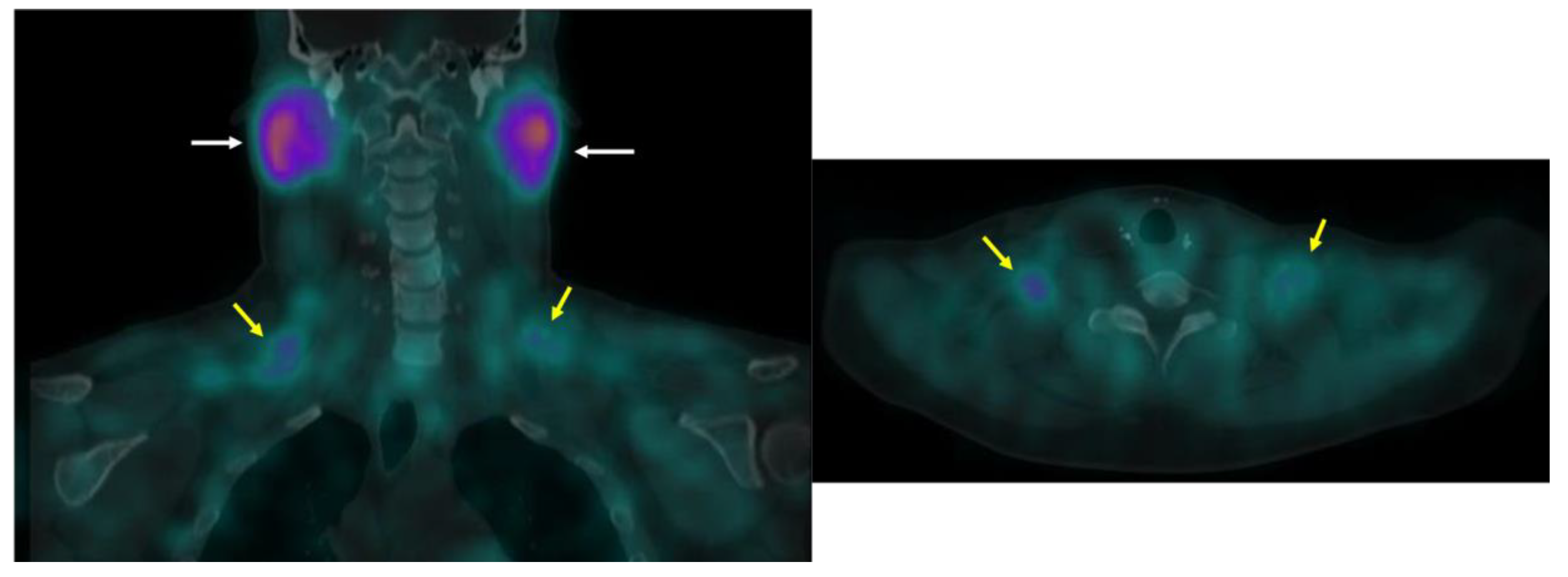
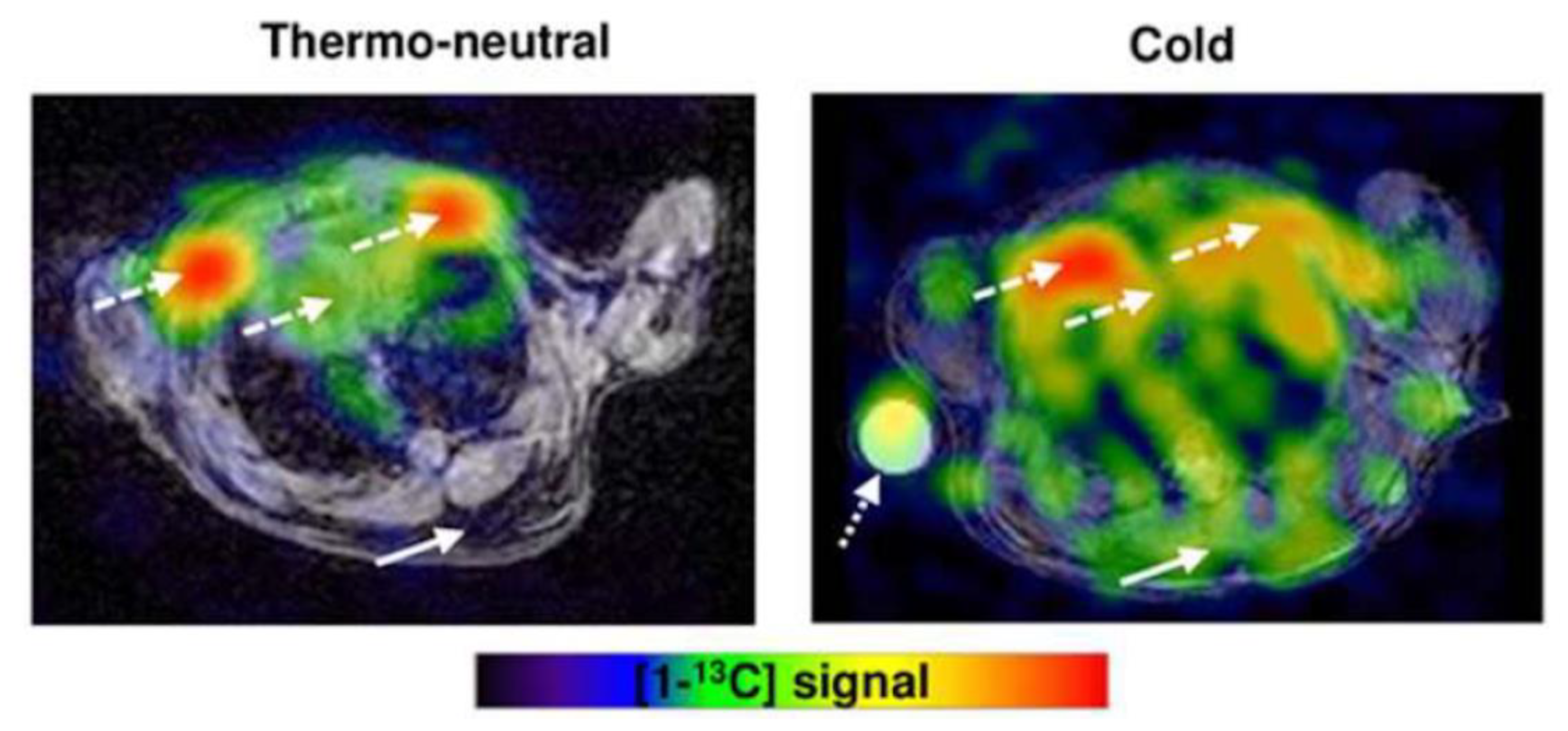
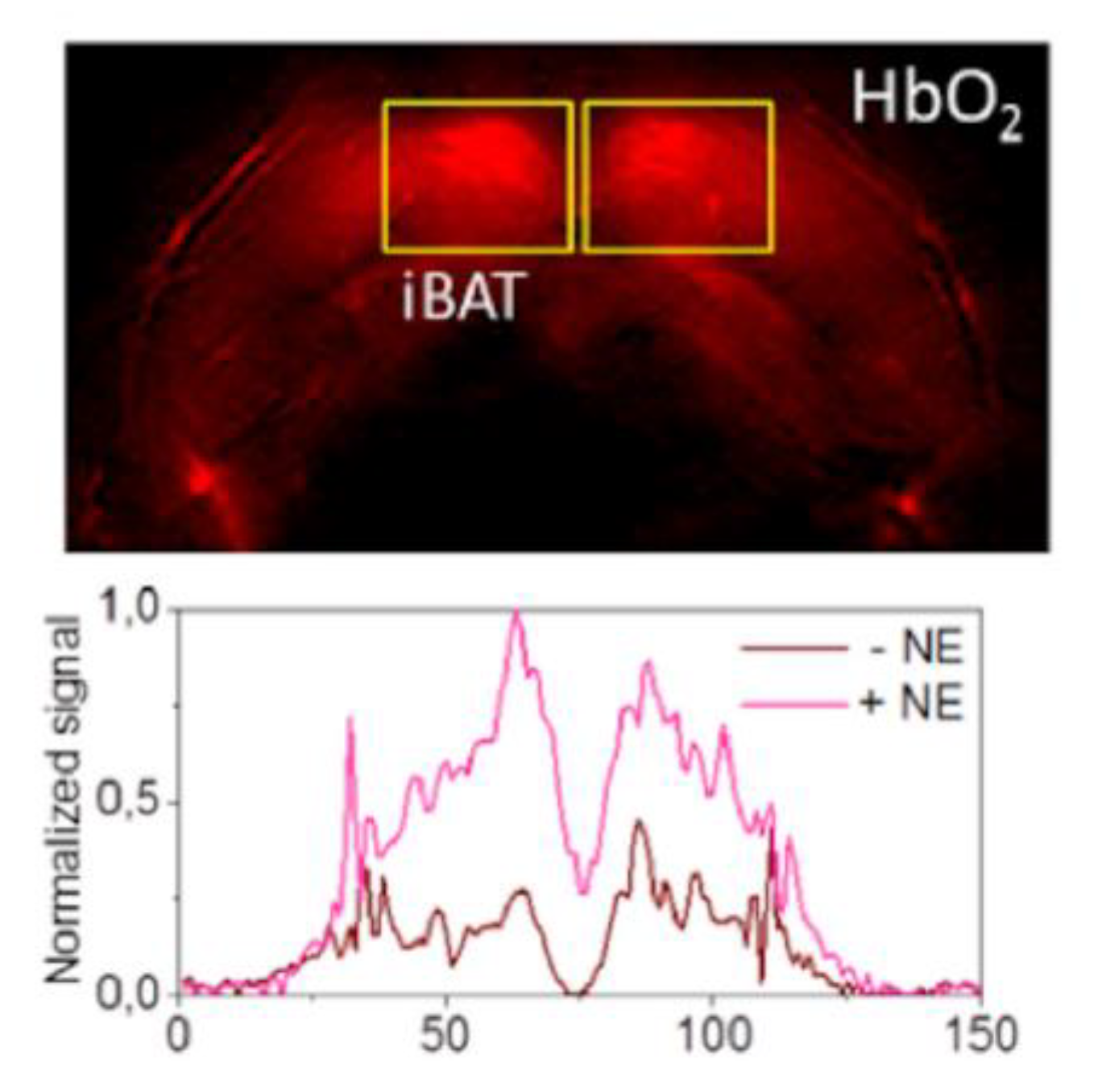
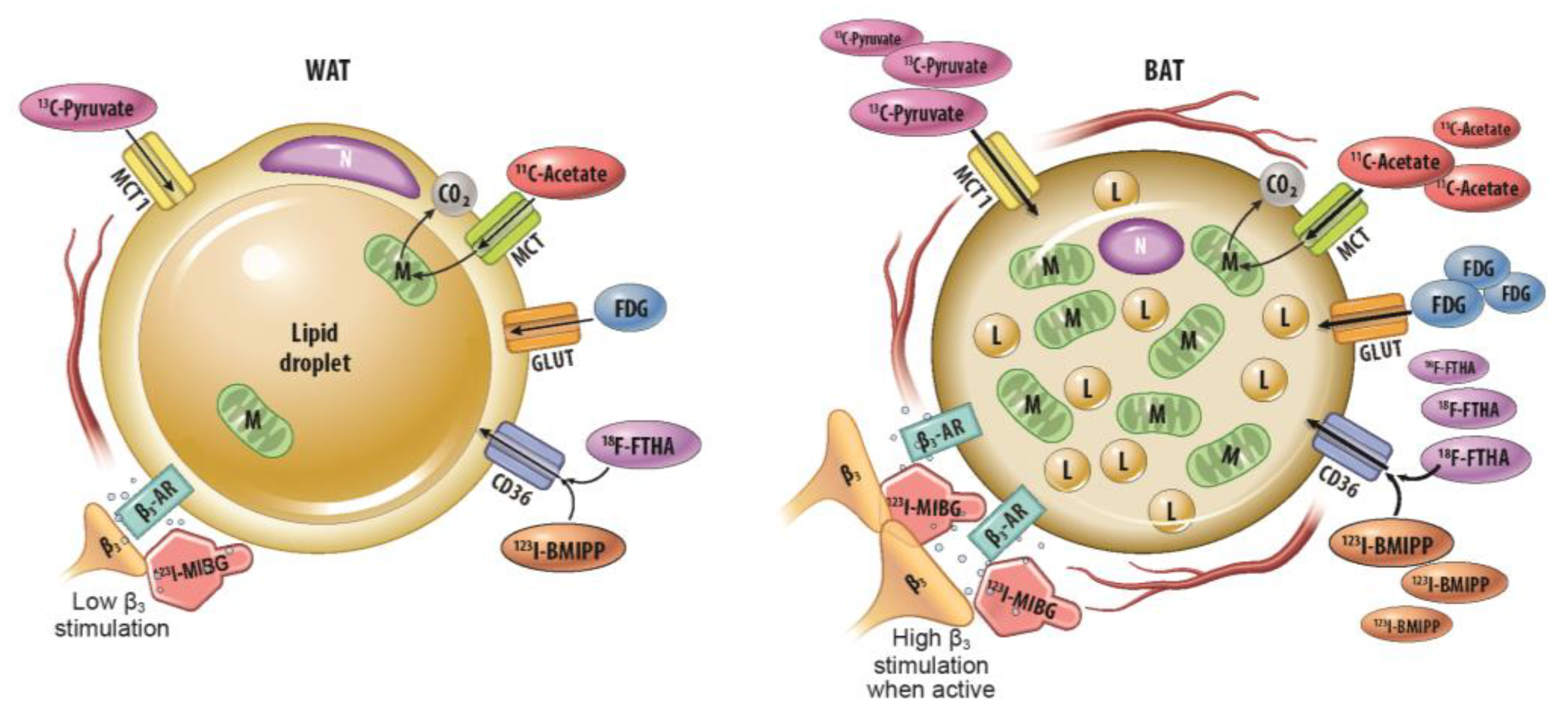
Abstract
Share and Cite
Frankl, J.; Sherwood, A.; Clegg, D.J.; Scherer, P.E.; Öz, O.K. Imaging Metabolically Active Fat: A Literature Review and Mechanistic Insights. Int. J. Mol. Sci. 2019, 20, 5509. https://doi.org/10.3390/ijms20215509
Frankl J, Sherwood A, Clegg DJ, Scherer PE, Öz OK. Imaging Metabolically Active Fat: A Literature Review and Mechanistic Insights. International Journal of Molecular Sciences. 2019; 20(21):5509. https://doi.org/10.3390/ijms20215509
Chicago/Turabian StyleFrankl, Joseph, Amber Sherwood, Deborah J. Clegg, Philipp E. Scherer, and Orhan K. Öz. 2019. "Imaging Metabolically Active Fat: A Literature Review and Mechanistic Insights" International Journal of Molecular Sciences 20, no. 21: 5509. https://doi.org/10.3390/ijms20215509
APA StyleFrankl, J., Sherwood, A., Clegg, D. J., Scherer, P. E., & Öz, O. K. (2019). Imaging Metabolically Active Fat: A Literature Review and Mechanistic Insights. International Journal of Molecular Sciences, 20(21), 5509. https://doi.org/10.3390/ijms20215509