1. Introduction
Endometriosis is a chronic gynecological estrogen-dependent disease characterized by the presence of endometrial tissue, both glands and stroma, outside the uterus. There are multiple possible locations for this ectopic tissue which may be grouped into three endometriosis subtypes: peritoneal, ovarian, with ovarian cysts called endometriomas, and deeply infiltrative. It is a debilitating disorder affecting around 10% of women of reproductive age [
1], with pelvic pain and infertility as the two main symptoms. The etiology and physiopathology of this disease remain unknown and there are no clinical biomarkers; in consequence, there is no cure and a long delay in the diagnosis. Studies focused on the discovery of diagnostic tools for endometriosis to help to design effective treatments that preserve fertility are of great interest.
Inflammation is necessary for the establishment and maintenance of endometrial cells in ectopic locations [
2,
3,
4,
5]. Purinergic signaling, the group of biological effects mediated by extracellular nucleotides, such as adenosine triphosphate (ATP), and nucleosides, such as adenosine, is involved in a wide range of physiological and pathological inflammatory conditions [
6]. Extracellular ATP is mostly a proinflammatory molecule released during tissue stress situations, such as necrosis or apoptosis, hypoxia, and inflammation. Purinergic signaling is also studied in the context of human reproduction [
7,
8,
9,
10,
11]; for instance, ATP is involved in the initiation and maintenance of myometrium and oviduct contractions. It increases the oviductal ciliary beat frequency [
12] and contributes to the regulation of the uterine fluid microenvironment [
9]. Moreover, adenosine, an ATP hydrolysis product, is necessary for sperm capacitation [
10]. ATP is also a pain-related molecule, and some of the pharmacological treatments used to relieve pain in endometriosis do indeed affect ATP levels or their effects. Moreover, extracellular ATP and its derivative adenosine influence cell migration, proliferation and survival—three necessary events for the establishment of ectopic endometrial foci.
Extracellular ATP and adenosine levels are controlled by the ecto-nucleotidases, which are broadly expressed enzymes that, acting alone or sequentially, hydrolyze ATP into adenosine. There are four families of ectonucleotidases: (i) the ecto-nucleoside triphosphate diphosphohydrolase (E-NTPDase) family, also known as CD39 family, which hydrolyzes the ATP, and adenosine diphosphate (ADP), to adenosine monophosphate (AMP); (ii) the ecto-nucleotide pyrophophatase/phosphodiesterase (E-NPP) family, which mainly hydrolyzes ATP to AMP; (iii) the 5′-nucleotidase (5′-NT) (known as CD73) that dephosphorylates AMP to adenosine; and, (iv) the alkaline phosphatases family that hydrolyzes nucleoside triphosphates and diphosphates to monophosphates [
6]. Adenosine deaminase (ADA) inactivates adenosine. This is a soluble enzyme often associated with CD26/dipeptidyl peptidase IV, expressed at the cell membrane [
13]. Ectonucleotidases and CD26 are well characterized in human cyclic and postmenopausal endometria, showing differences in its expression and distribution throughout the cycle [
8,
14]. The presence of ectonucleotidases in the contents of endometrioma had previously been described [
15,
16] but no studies had yet to be conducted on eutopic and ectopic endometrial tissue from women with endometriosis.
With the present study, we aimed to characterize the expression of ectonucleotidases in the eutopic and ectopic endometrial tissue of women with endometriosis and compare it with the eutopic endometrium of women without this pathology. We believe that assessing the participation of these proteins directly involved in ATP breakdown in endometriosis could contribute to facilitating the diagnosis and ameliorating the treatment status of this pathology.
3. Discussion
Purinergic signaling plays a role in reproduction, and changes in its elements have been described in the pathology of endometriosis. Extracellular ATP may be involved in two of the major symptoms of endometriosis, which are infertility and pain [
9,
15,
16]. In the present study we characterized the expression in eutopic and ectopic endometrial tissue of different ectonucleotidases involved in the regulation of ATP levels in tissue microenvironment. We have compared the results with those previously published in non-pathological endometria [
8,
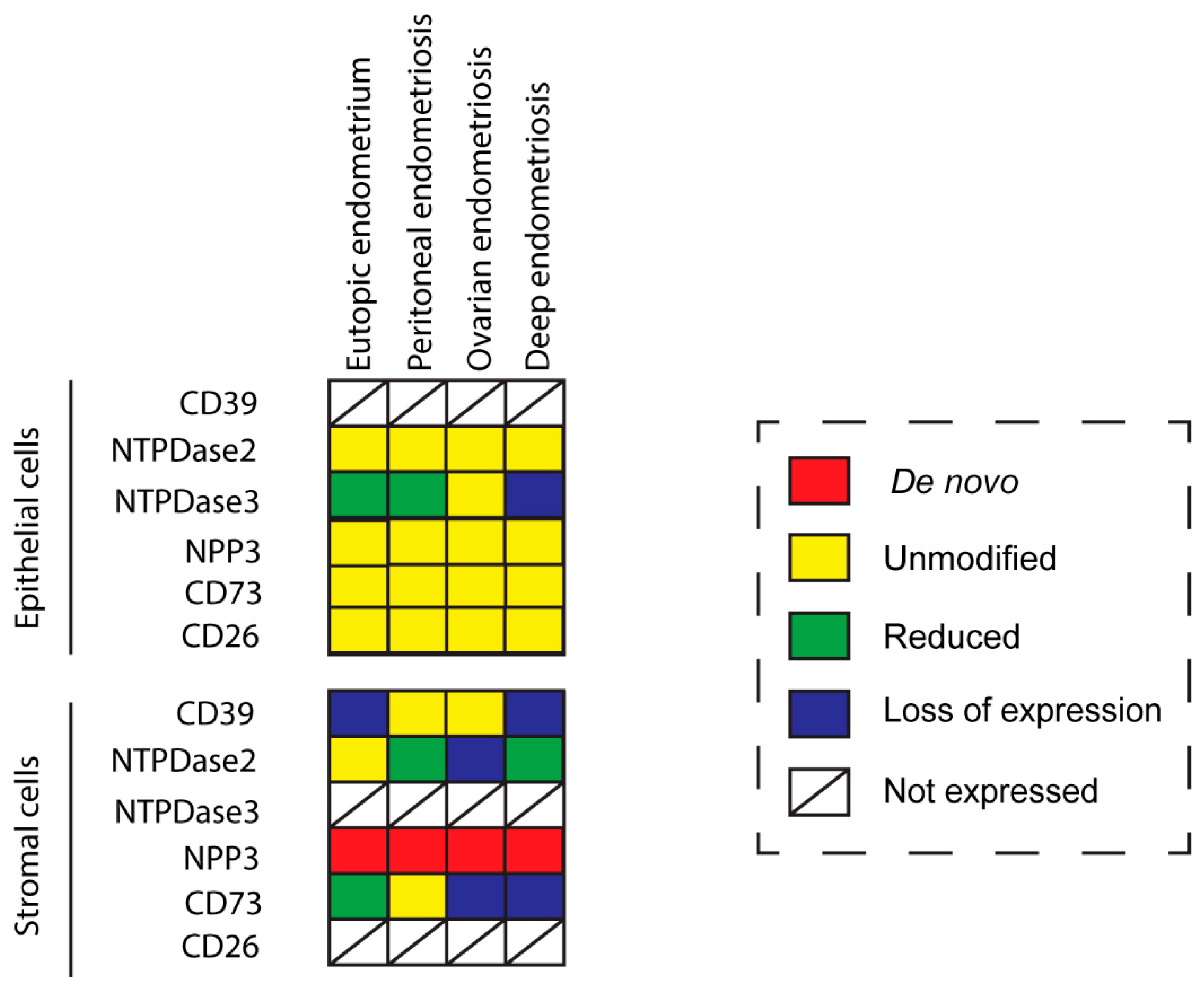
14]. Tissue distribution in endometriosis coincides with the control condition except in the case of NPP3 which is present in stroma only in endometriosis. Ciliated cells of endometria display the same expression pattern as in control fallopian tubes. Changes in expression and activity are consistently recorded, the greatest being in the stroma (
Figure 8). These findings provide information to elucidate the cellular and molecular mechanism as well as the etiology and the progression of the disease, which might help to identify new diagnostic and therapeutic targets.
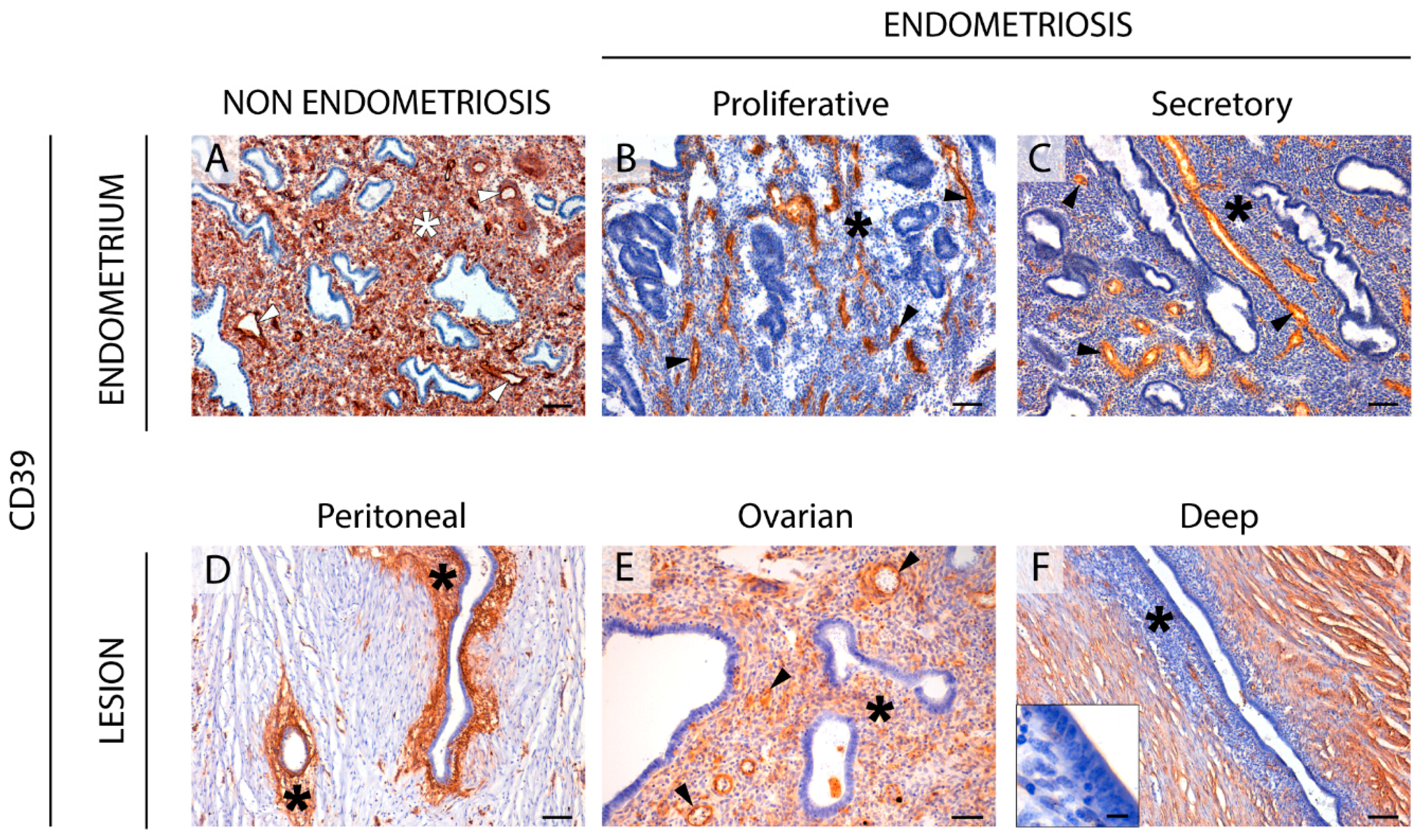
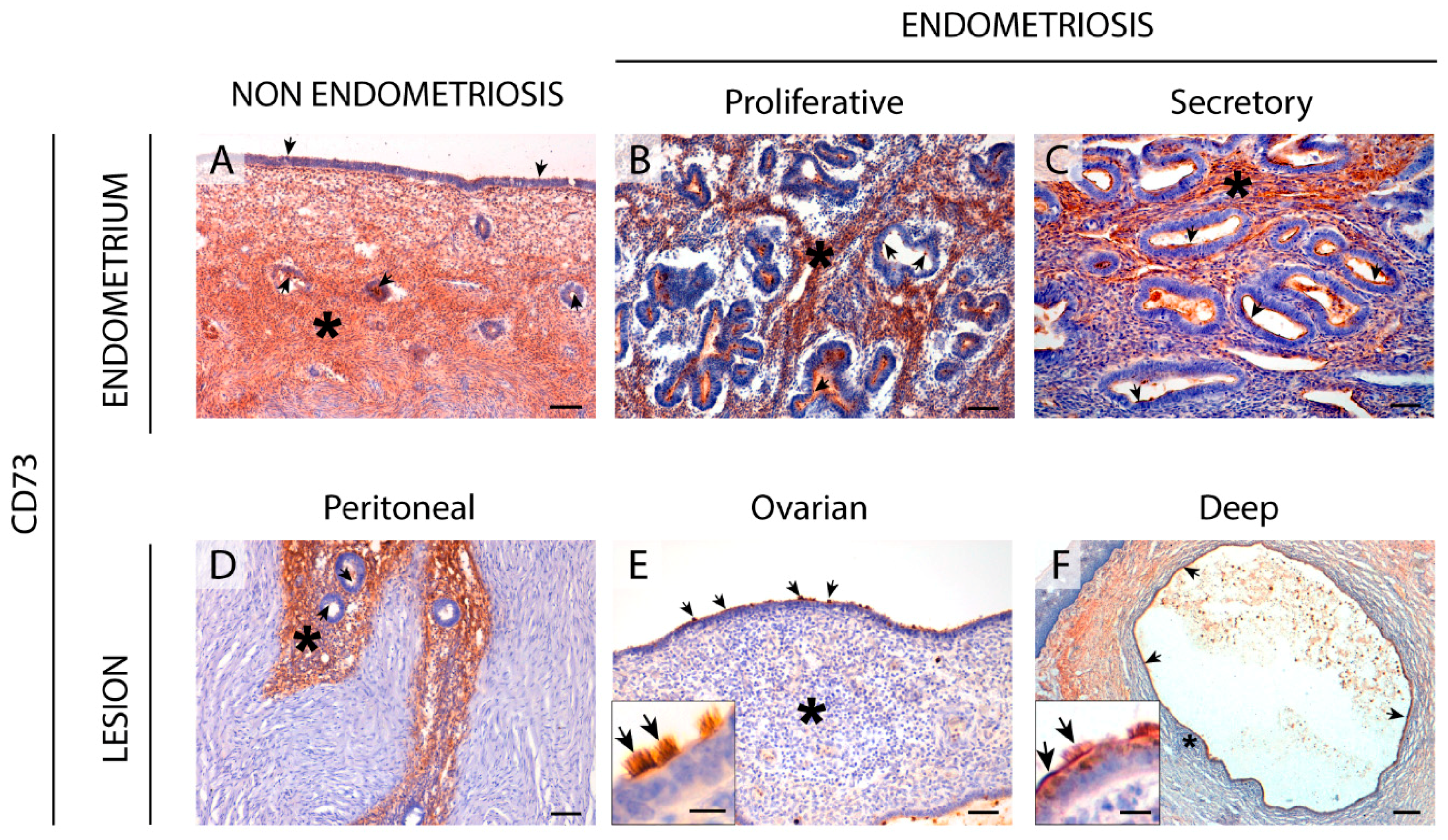
The CD39-adenosinergic axis, with CD39 and CD73 acting sequentially to hydrolyze ATP to adenosine, is considered the main duo responsible for metabolizing extracellular ATP, generating an immunosuppressive adenosine-rich microenvironment in physiological and pathophysiological conditions [
17]. In endometrium, the expression pattern of these ectonucleotidases and their changes throughout the cycle are well studied. Under physiological conditions, endometrial stromal cells express both CD39 and CD73. While CD39 expression is constant throughout the cycle, CD73 fluctuates [
8,
18], thus determining variations in adenosine level in the microenvironment. In the present study we note that eutopic endometrium of women with endometriosis displays the same already known expression pattern of CD73 but mostly loses CD39 stromal expression. A plausible consequence of the concomitant unbalanced ATP hydrolysis is the accumulation of extracellular ATP in the endometrial stromal microenvironment. This might well play a role in the generation and maintenance of the chronic inflammatory state of endometria of women with endometriosis. Moreover, extracellular ATP is closely related to various immune and inflammatory factors that are known to be involved in the infertility of women with endometriosis, by reducing the quality of gametes and their rates of transport and implantation, and by increasing the pregnancy loss rate [
19].
This situation could explain the de novo stromal expression of NPP3 in endometriosis throughout the cycle and in atrophic endometrium, as a cellular tool to offset the loss of ATPase activity due to the lack of CD39. However, NPP3 action would not be sufficient to replace ATPase activity because NPP3 has a lower affinity for ATP than CD39 [
6]; moreover stromal NPP3 expression is not coordinated with CD73 expression throughout the cycle.
Changes in the CD39-CD73 pathway were also found in endometriotic lesions. Our findings indicate that the changes in ATP hydrolysis resulting from CD39 and CD73 activity are related to the severity of endometriosis since their expression is lost in deep infiltrating lesions. These changes of expression would lead to an extracellular ATP accumulation that would in turn promote the secretion of cytokines and growth factors into the ectopic milieu, with a concomitant increase in survival and growth rates of endometrial cells [
20,
21]. Unfortunately, the exact function of extracellular ATP in endometriosis is not clear. While ATP signaling seems to be closely related to the origin and progression of endometriosis, intramuscular injection of ATP in a rat model of endometriosis was found to reduce the size of the ectopic induced lesions [
22]. Our results are in line with previous studies that showed differing expression of the protein ATPase Na+/K+ Transporting Family Member Beta 4 (ATP1B4) between patients with and without endometriosis, in favor of a decrease of the hydrolysis of ATP in the endometriosis patients [
23,
24]. The authors stated that ATP was clearly related with the formation and development of endometriosis disease. Moreover, pain is a characteristic symptom of endometriosis, and ATP is a pain factor mainly acting through the purinoreceptor P2X3 that has also been studied in endometriosis. P2X3 has been found in the epithelial and some stromal cells of eutopic and ectopic endometrial tissue as well as on sensory nerve fibers in endometriotic lesions. Its expression levels correlate with the severity of pain in women with endometriosis [
25]. Moreover, the use of A-317491, a selective P2X3 receptor antagonist, relieved pain with a prolonged antinociceptive effect in rats [
26], and the receptor is thus a target for the pharmacological approach of endometriosis pain relief. Therefore, the increased levels of extracellular ATP might well be related to the endometriosis-associated pain. These results parallel the difference in CD73 and CD39 expression between the three entities of endometriosis, where the most extreme change has been detected in the deep infiltrating lesions, an important indicator of the severity of pain in endometriosis [
27]. Additionally, the downregulation of CD73 has been described in poorly differentiated and advanced-stage endometrial carcinoma. Adenosine generated by the activity of CD73 located in the areas of cell-cell contacts regulates cell-cell adhesions by the regulation of the primary component of filopodia (F-actin). In fact, cell migration and invasion in high-grade and advanced-stage endometrial carcinomas is dependent on the loss of the adenosine generated by CD73 [
28]. According to Sampson’s theory of retrograde menstruation, endometrial tissue detached during menses has to travel through the fallopian tubes to the ectopic site of implantation, such as the ovarian surface or the peritoneal wall, and must then invade and adhere to the self-tissue of the new localization, proliferating and evading the immune response to form the endometriotic lesion [
29]. For this reason, the loss of CD73 in the two most severe entities of endometriosis, ovarian and deep endometriosis, as well as its implication in the inflammatory state of endometriosis, might also play a role in the migration and invasive properties of ectopic cells needed to generate the lesion.
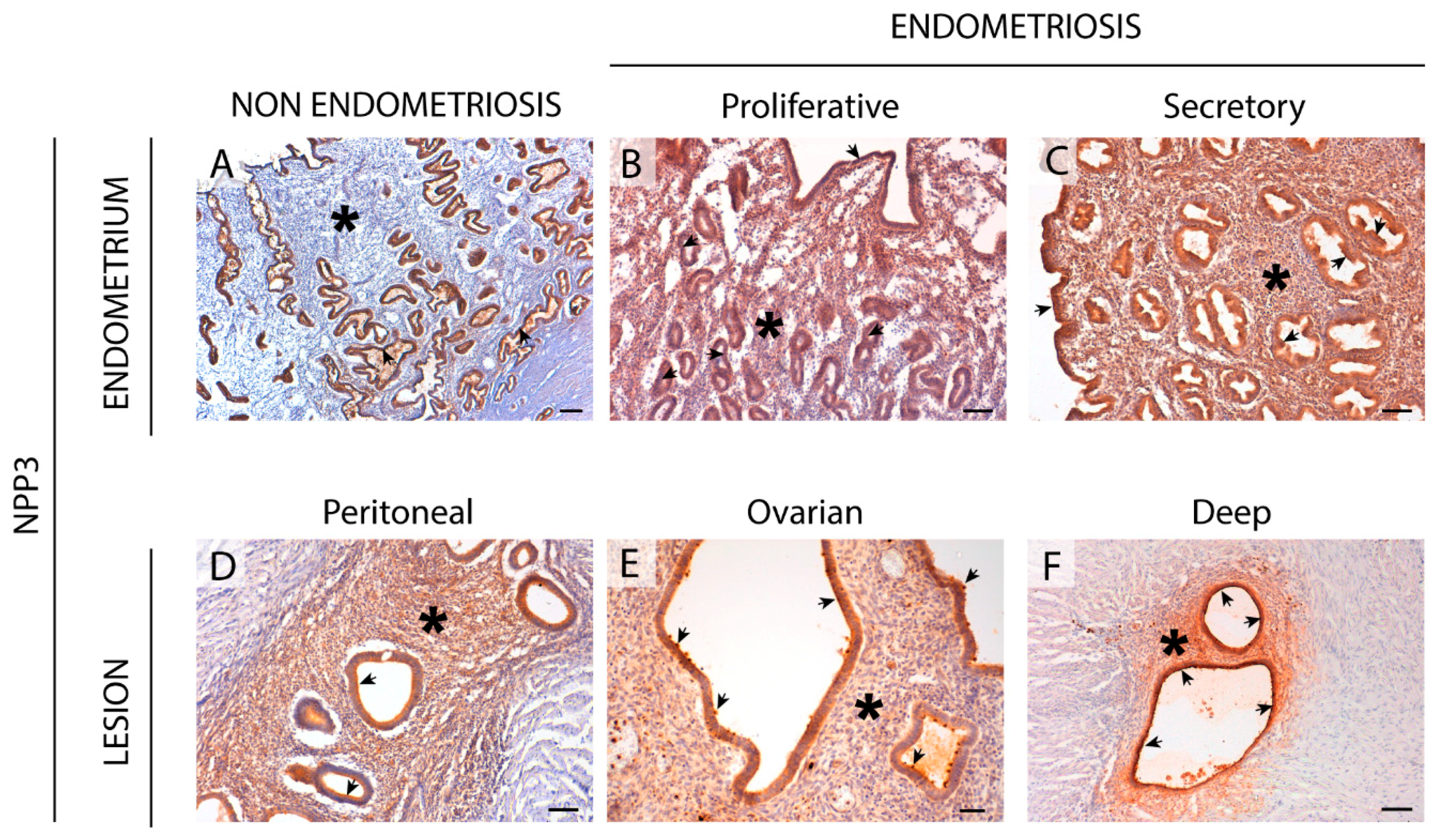
An important finding is the identification of NPP3 label as a new histopathological marker of the disease since we have demonstrated its expression and activity in the stroma only in endometriosis, in both eutopic and ectopic tissues. NPP3 has already been identified in endometrial epithelial cells, a fact that is also confirmed in the case of endometriosis without any variation. A previous study by our group also demonstrated the presence of NPP3 in the contents of endometriomas although the levels did not differ from those of the simple ovarian cysts used as controls and therefore its presence was not exclusive of endometriosis [
16]. The relevance of the study reported here is its presence in eutopic endometria which discriminates between endometriosis and non-endometriosis conditions, which might allow its use as a histopathological diagnostic tool. To our knowledge, NPP3 has been identified in epithelial cell types, in cells of the immune system, mainly mast cells, and in tumor cells with an epithelial or myeloid origin [
8,
30,
31,
32,
33,
34]. The specific detection of NPP3 in the endometrial stromal cells of eutopic endometria and in all three entities of endometriosis can be used as a histopathologic marker of endometriosis disease. In addition to its role in the control of extracellular ATP levels, NPP3 might well play a role in the invasive capacity of the stromal endometrial cells in endometriosis since it is known that overexpression of NPP3 in murine fibroblasts stimulates the motility and the invasiveness of these cells [
35]. Our finding of de novo NPP3 in stromal cells, with greater expression in the functional layer which is shed during menses, and the relation of NPP3 with the cell motility and invasion, suggest involvement of NPP3 in the formation and progression of endometriotic lesions based on the retrograde menstruation theory [
29]. Besides the importance of NPP3 as histopathological marker, additional studies are needed to determine the precise role of NPP3 in the pathogenesis and progression of endometriosis. It might well be a new target for pharmacological therapy of endometriosis. Indeed, targeting NPP3 is feasible since phase 1 trials using an antibody drug conjugate targeting this protein have been completed in patients with advanced metastatic renal cell carcinoma with promising antitumor results [
36].
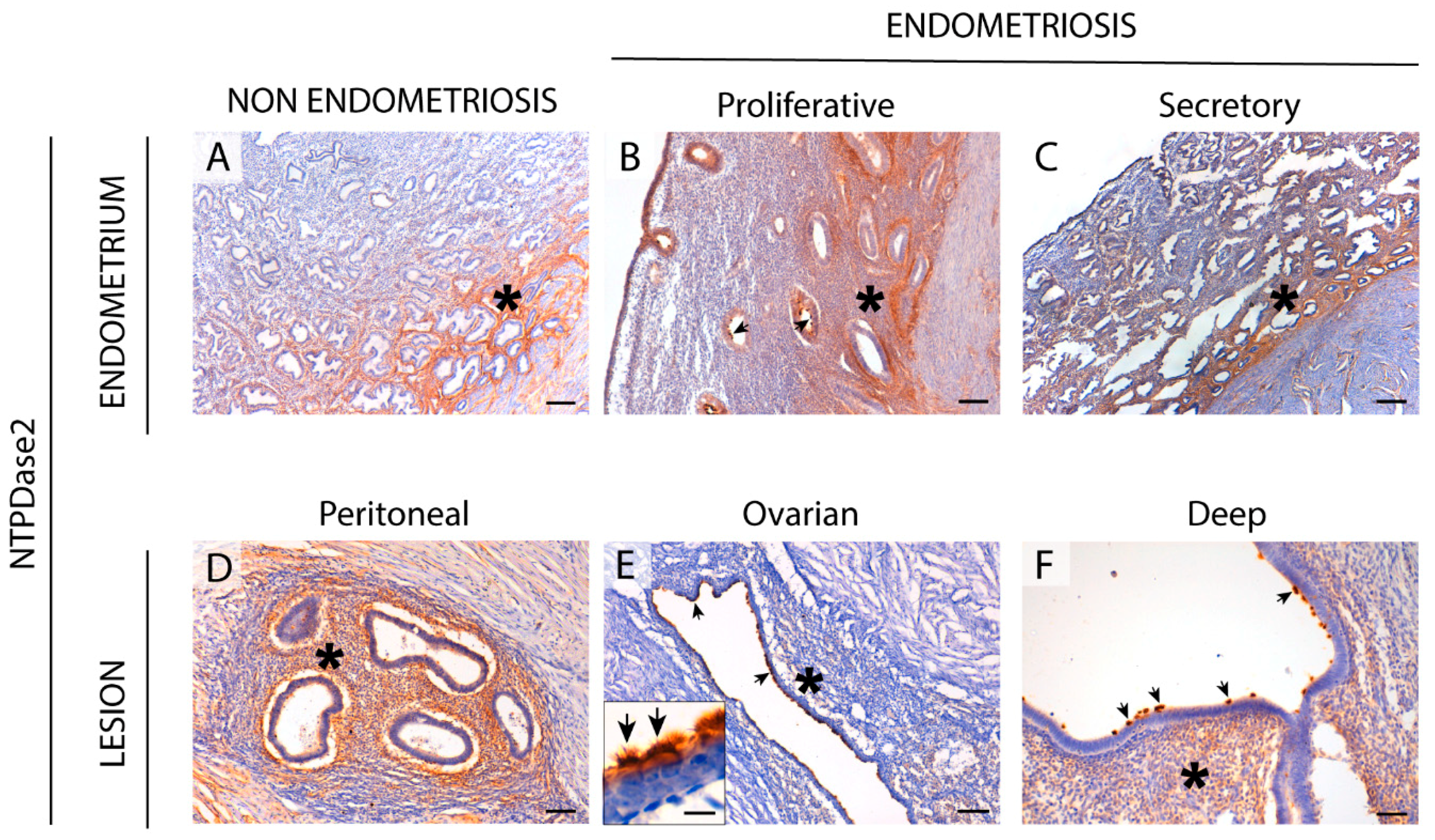
We found NTPDase2 expression in the same cell types and structures as in the non-endometriosis condition. Moreover, NTPDase2 was expressed by perivascular cells in some lesions with colocalization with the eMSCs marker SUSD2. Functional studies are needed to determinate whether NTPDase2+ SUSD2+ cells are eMSCs as in the eutopic endometrium. Retrograde shedding of stem cells into the pelvic cavity without immune clearance is thought to be lesion-initiating. Therefore, it would be of interest to compare lesions containing the NTPDase2+ SUSD2+ cell population with lesions without it.
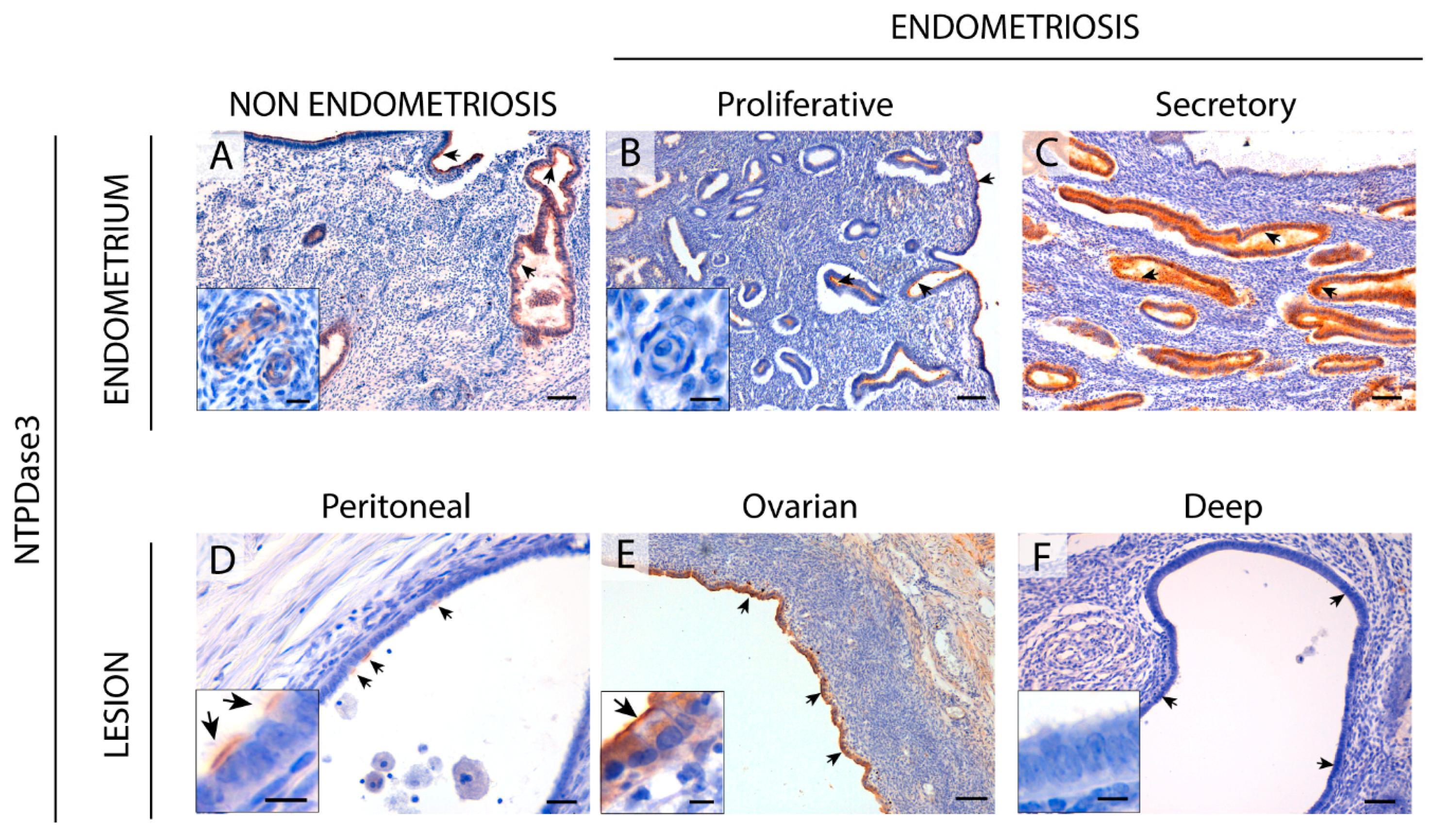
NTPDase3 was described in epithelial cells and spiral arteries in healthy endometria. In fact, NTPDase3 has been considered a spiral artery marker [
8]. But we did not find NTPDase3 labeling in spiral arteries of women with endometriosis. Spiral artery remodeling plays a central role in establishing and maintaining a normal pregnancy, and impaired remodeling is involved in common pregnancy disorders. This might be also one of the mechanisms underlying the decreased pregnancy rates in women with endometriosis. It is important to highlight the loss of NTPDase3 in the epithelial cells of deep infiltrating lesions. Although NTPDase3 has been little explored in pathological conditions, a decrease in
ntpdase3 expression has been described during the induction of mouse bladder cancer, suggesting its participation in cancer establishment and progression [
37]. This result, together with the loss of NTPDase3 in the epithelial cells of the most severe form of endometriosis, provides further evidence of the need to study its role in the pathophysiology of endometriosis and cancer.
CD26 or dipeptidyl peptidase IV (DPPIV) is a membrane glycoprotein that binds, among other peptides, the ectoenzyme ADA in humans. It is involved in the protection of the tissue against local inflammation and in intracellular signaling. CD26 has been described as a cancer stem cell marker and tumor suppressor protein in certain types of cancer. By contrast, CD26 overexpression promotes cell proliferation, invasion, and tumorigenesis in endometrial carcinoma cells [
38]. In endometriosis, Tan et al. [
39] described the increase of endometrial stromal cell migration and invasion in part by reduced expression of CD26 under hypoxia conditions and also by CD26 inhibition. Other studies performed in tissue, including ours, have not matched these in vitro results with cell culture since we were not able to detect CD26 in endometrial stroma, but only in epithelial glandular cells. This might be due to the differing behavior of cells in vitro or even to technical reasons. Here, we show high expression of CD26 in the epithelial cells of eutopic endometrium and in ectopic tissue. The difference with the endometrial expression in women without endometriosis is that CD26 expression in endometriosis is constant throughout the cycle. It would be interesting to see whether the high expression of CD26 in ectopic epithelial cells has a similar effect to that of endometrial carcinoma cells on cell migration and invasion ability. In relation to the ATP metabolism, knowing the levels of ADA, the soluble enzyme that hydrolyses the extracellular adenosine to control the immunosuppressive milieu, is key to understanding what is happening in endometriosis. In a previous study, high levels of ADA were found in the contents of ovarian endometriomas [
16]. We were, however, unable to detect ADA by immunostaining due to the technical limitations of the antibodies available, and we cannot be certain whether high levels of CD26 in tissue is related to an increase in ADA activity.
The changes in the expression of the ectonucleotidases described here in eutopic and ectopic endometrium argue for extracellular ATP accumulation. The greatest loss of ectonucleotidase expression was found in the deep infiltrating endometriosis, the most severe endometriosis subtype [
40,
41]. Our results, together with the role of ATP in pain [
25,
42], lend support to the involvement of ectonucleotidase expression changes with the severity of endometriosis. Moreover, our results reinforce the relevance of the stroma and tissue microenvironment in the etiopathology and progression of endometriosis disease. Future studies on the role of purinergic signaling in endometriosis are needed to identify biomarkers of the disease and to develop new therapeutic strategies that would allow for earlier detection and respect for the reproductive wishes of women with endometriosis. However, unlike in cancer, where ectonucleotidase blockade is a therapeutic tool, in endometriosis the use of inhibitors of ectonucleotidases does not seem to represent an appropriate strategy. On the contrary, increasing the ATPase activity would combat the eventual ATP accumulation of endometrial microenvironment. In line with this, the use of A-317491, an antagonist of the ATP receptor P2X3, relieves pain in endometriosis [
26]. Administration of soluble CD39 is known to be safe and is well studied in the context of cardiovascular diseases where it is known to prevent thrombus formation (reviewed in [
43]).
4. Materials and Methods
4.1. Samples
The ethical principles of this study adhere to the Helsinki Declaration, and all the procedures were approved by the ethics committee for clinical investigation of Bellvitge Hospital (project identification code PR090/15, Acta 21/16, 12/2016). All the patients included gave written informed consent. Fifty-seven patients with endometriotic lesions (ectopic endometrial tissue) were recruited for the study by the Gynecology Service of Bellvitge Hospital (Barcelona, Spain) between March 2016 and July 2019, and by the Gynecology Service of Dexeus Institute (Barcelona, Spain) between October 2016 and March 2018. Thirty-four endometrium samples were obtained by the Gynecology Service of Bellvitge Hospital from women without endometriosis or endometrial malignancy as a control group (including 10 proliferative, 4 secretory, and 20 atrophic endometria; age mean of patients = 55.03 years, standard derivation = 11.83).
Human endometrial samples from women with endometriosis (
n = 25) were obtained from hysterectomy specimens without endometrial malignancy at the pathology services of Bellvitge Hospital and Dexeus Hospital. Peritoneal endometriosis (
n = 7), ovarian endometriosis (
n = 27), and/or deep endometriosis (
n = 28) were surgically removed in the gynecology services of the same hospitals. Demographic description of the samples from the women with endometriosis are summarized in
Table 3. Endometrial dating was carried out by the pathology services.
Excised tissue samples were fixed with 4% paraformaldehyde, cryoprotected by introducing them into a 30% (w/v) sucrose solution at 4 °C for 24 h, and then embedded in O.C.T freezing media (Tissue-Tek®; Sakura Finetk, Zoeterwoude, Netherlands). Fifteen μm sections were obtained using a Cryostat Leica CM1950 (Leica, Wetzlar, Germany). Sections were put onto poly-l-lysine coated glass slides and stored at −20 °C until use. Routine haematoxylin and eosin staining was performed.
4.2. Antibodies
Primary antibodies used in this study are listed in
Table 4. Secondary antibodies used for immunohistochemistry were horseradish peroxidase (HRP)-conjugated goat anti-mouse (EnVision™ + System; DAKO, Carpinteria, CA, USA) and HRP-conjugated goat anti-rabbit (EnVision™ + System).
Secondary antibodies used for immunofluorescence assays were Alexa Fluor 488 goat anti-mouse and Alexa Fluor 647 goat anti-rabbit (Thermo Fisher Scientific, Rockford, Illinois, USA). Secondary antibodies were used at 1:500 and dilutions were made in PBS.
4.3. Immunolabeling Experiments
Slices were washed twice with PBS to remove the O.C.T freezing media and then pre-incubated for 1 h at room temperature (RT) with PBS containing 20% normal goat serum (NGS, Gibco, Paisley, UK), 0.2% Triton and 0.2% gelatin (Merck, Darmstadt, Germany). For immunohistochemistry experiments a previous blocking of endogenous peroxidase activity was performed with 10% methanol (
v/v) and 2% H2O2 (
v/v) in PBS for 30 min. Slices were then incubated overnight (O/N) at 4 °C with the primary antibodies (listed in
Table 2) diluted in PBS. After three washes in PBS, tissue sections were incubated with the appropriate secondary antibodies for 1 h at RT, except HRP-goat anti-mouse and HRP-goat anti-rabbit, which were incubated for 30 min at RT. Secondary antibodies alone were routinely included as controls for the experiments.
For immunohistochemistry, the peroxidase reaction was performed in a solution containing 0.6 mg/mL 3, 3′-diaminobenzidine substrate (DAB; D-5637, Sigma-Aldrich, Saint Louis, MO, USA) and 0.5 µL/mL H2O2 in PBS for 10 min, and stopped with PBS. Nuclei were counterstained with haematoxylin and slides were then dehydrated and mounted with DPX mounting medium. Samples were observed under light Nikon Eclipse E200 and photographed under a light Leica DMD 108 microscope. In fluorescence assays, for nuclei labeling, slides were mounted with aqueous mounting medium with DAPI (ProLong™ Gold antifade reagent with DAPI, Life Technologies, Paisley, UK). Samples were then observed and photographed under a Zeiss LSM 880 Confocal Laser Scanning Microscope. Fluorescence images were processed with the software ZEN 2.3 SP1 (Zeiss, Oberkochen, Germany).
Immunohistochemical staining was independently evaluated by two observers. Staining distribution was recorded. Label intensity was scored as negative (-), weak (+), intermediate (++), or strongly positive (+++).
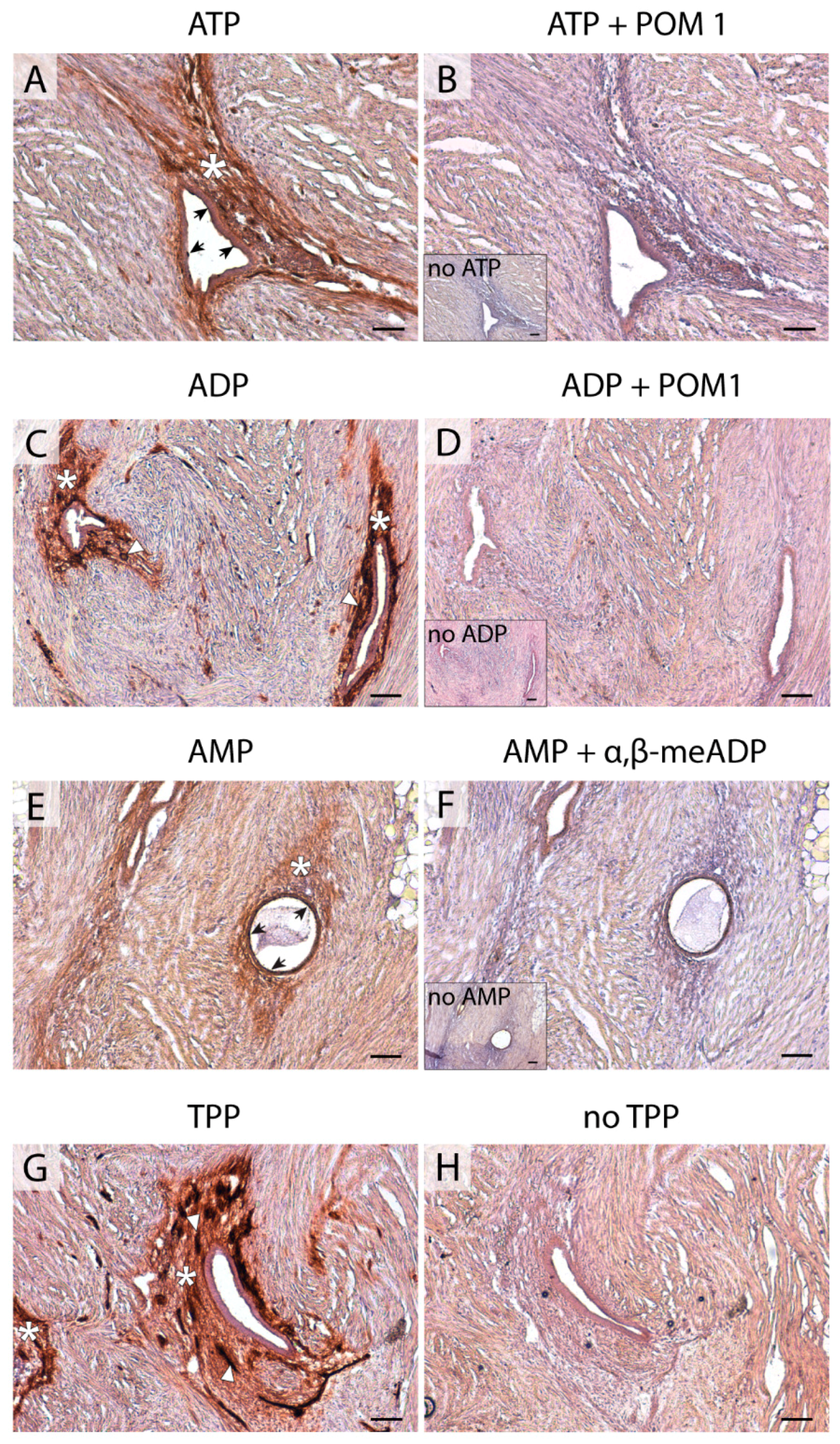
4.4. In situ ATPase, ADPase, AMPase, and TPPase Activity Experiments
A protocol based on the Wachstein/Meisel lead phosphate method was used [
8,
11,
44,
45]. The sections were washed twice with 50 mM Tris-maleate buffer pH 7.4 and pre-incubated for 30 min at RT with 50 mM Tris-maleate buffer pH 7.4 containing 2 mM MgCl
2 and 0.25 mM sucrose. The enzymatic reaction was carried out by incubating tissue sections for 1 h at 37 °C with 50 mM Tris-maleate buffer pH 7.4 supplemented with 0.25 mM sucrose, 2 mM MgCl
2, 5 mM MnCl
2, 3 % Dextran, 2 mM Pb(NO
3)
2, and 2 mM CaCl
2. All experiments were performed in the presence of 2.5 mM levamisole, as an inhibitor of alkaline phosphatase (AP) activity, and in the presence of 1 mM AMP, ADP, ATP, or TPP as a substrate. TPP is a false substrate, which can be cleaved by the pyrophosphatase activity of E-NPPs. Control assays were performed in the absence of nucleotide. For E-NTPDase inhibition experiments, 1 mM POM 1 was added to pre-incubation and enzymatic reaction buffers. For CD73 inhibition experiments, 1 mM α, β-meADP was added to pre-incubation and enzymatic reaction buffers. The reaction was revealed by incubation with 1% (NH
4)
2S (
v/v) for exactly 1 min. Nuclei were counterstained with haematoxylin. Samples were mounted with aqueous mounting medium (FluoromountTM, Sigma-Aldrich), observed under a light Nikon Eclipse E200 microscope, and photographed under a light Leica DMD 108 microscope.
4.5. Statistical Analysis
The predictive analytics software IBM SPSS Statistics v22 (IBM Corp., Armonk, NY, USA) was used for the creation of frequency tables with the distribution of ectonucleotidases in each endometrial component as well as the label intensity score in each case.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







