The Modulatory Effect of Gender and Cytomegalovirus-Seropositivity on Circulating Inflammatory Factors and Cognitive Performance in Elderly Individuals

Abstract
1. Introduction
2. Results
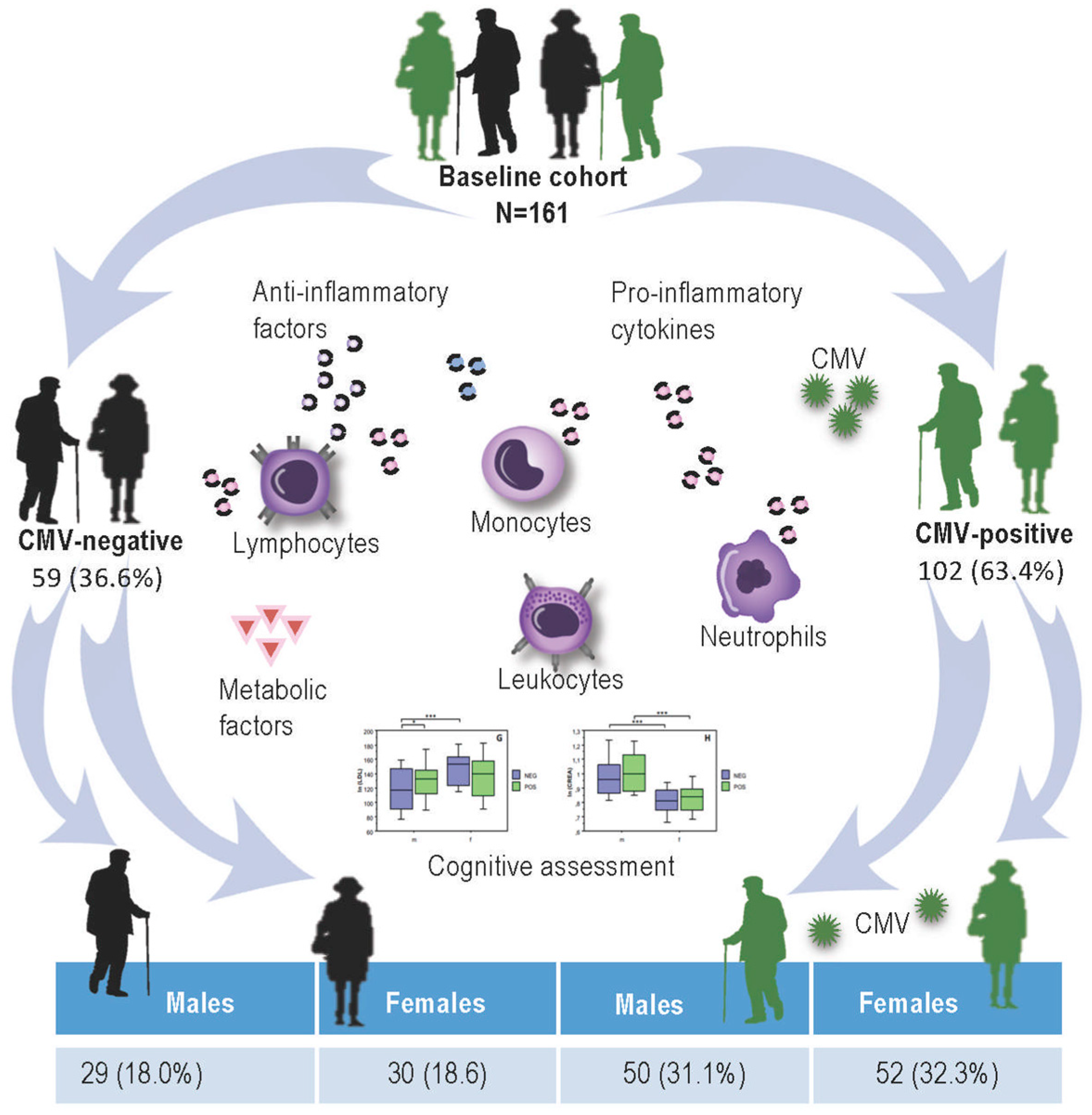
2.1. CMV-Serostatus of Study Participants
2.2. Influence of Gender and CMV-Serostatus on Circulating Levels of Pro- and Anti-Inflammatory Mediators, Immune Cells, and Metabolic Blood Values Analysed by MANOVA
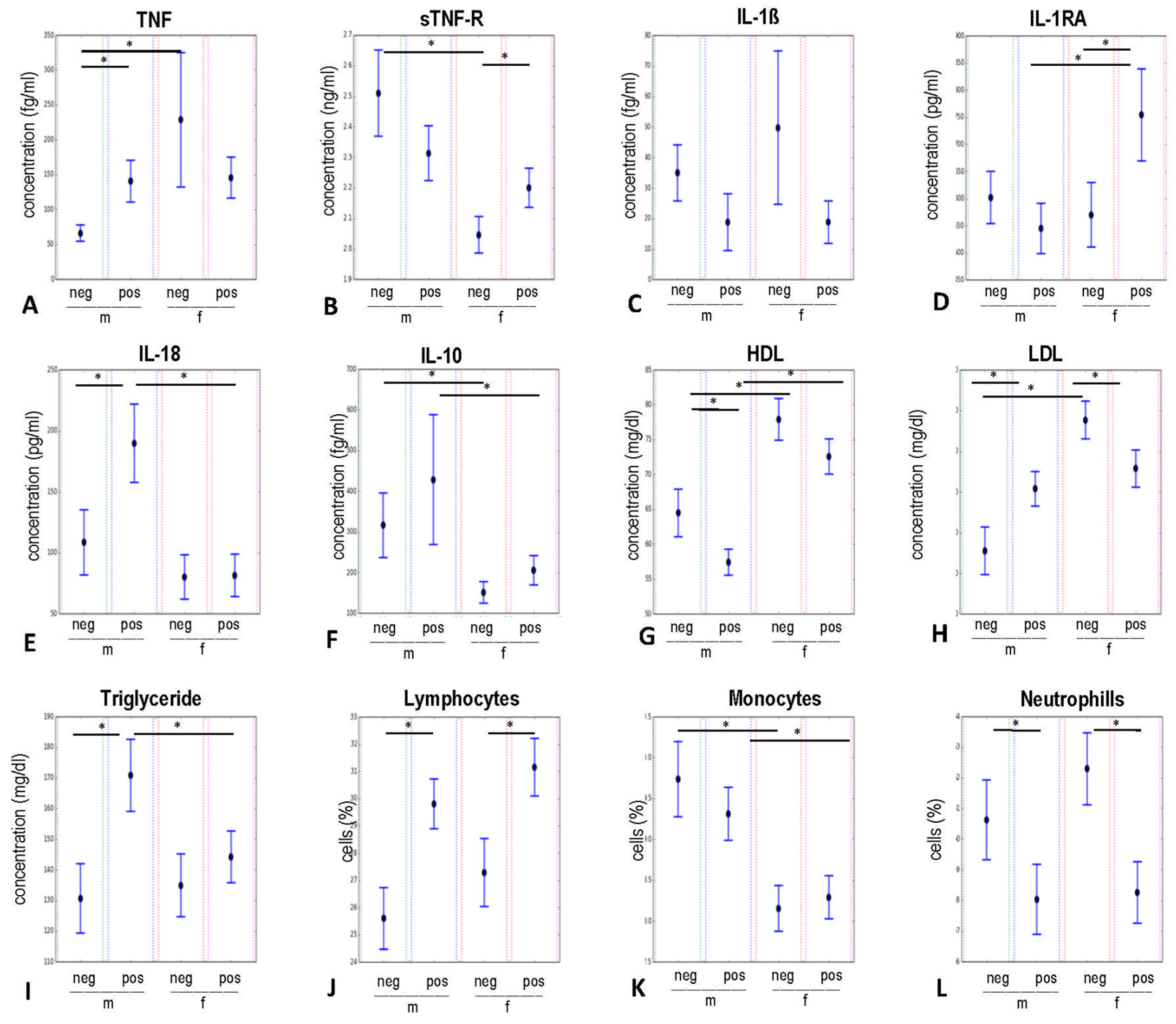
2.2.1. Pro- and Anti-Inflammatory Groups of Variables
2.2.2. Group of Metabolic Risk Variables
2.2.3. Group of Immune Cells Variables
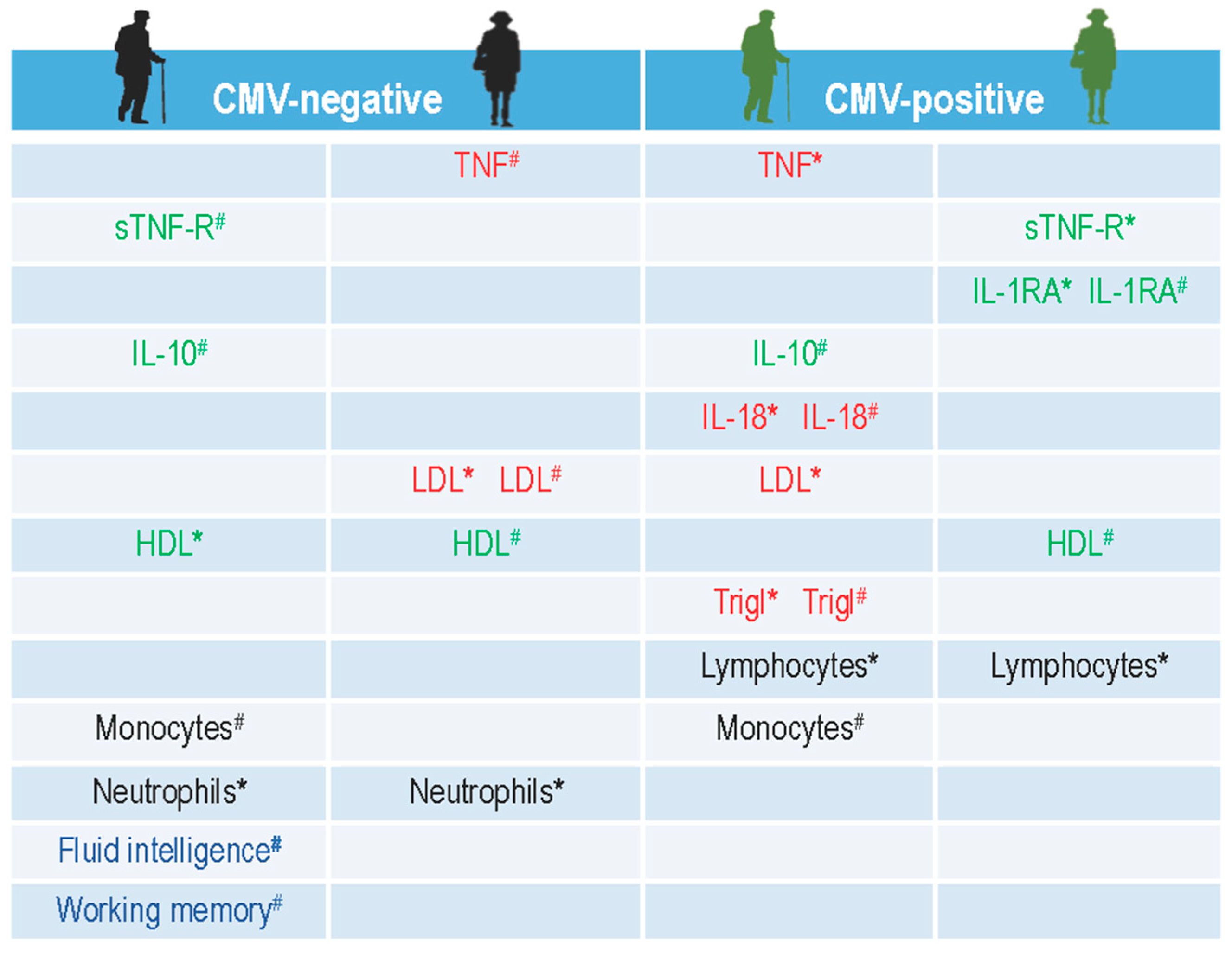
2.3. Influence of Gender and CMV-Serostatus on Circulating Levels of Pro- and Anti-Inflammatory Mediators, Immune Cells, and Metabolic Blood Values Analysed by Bootstrapping Approach
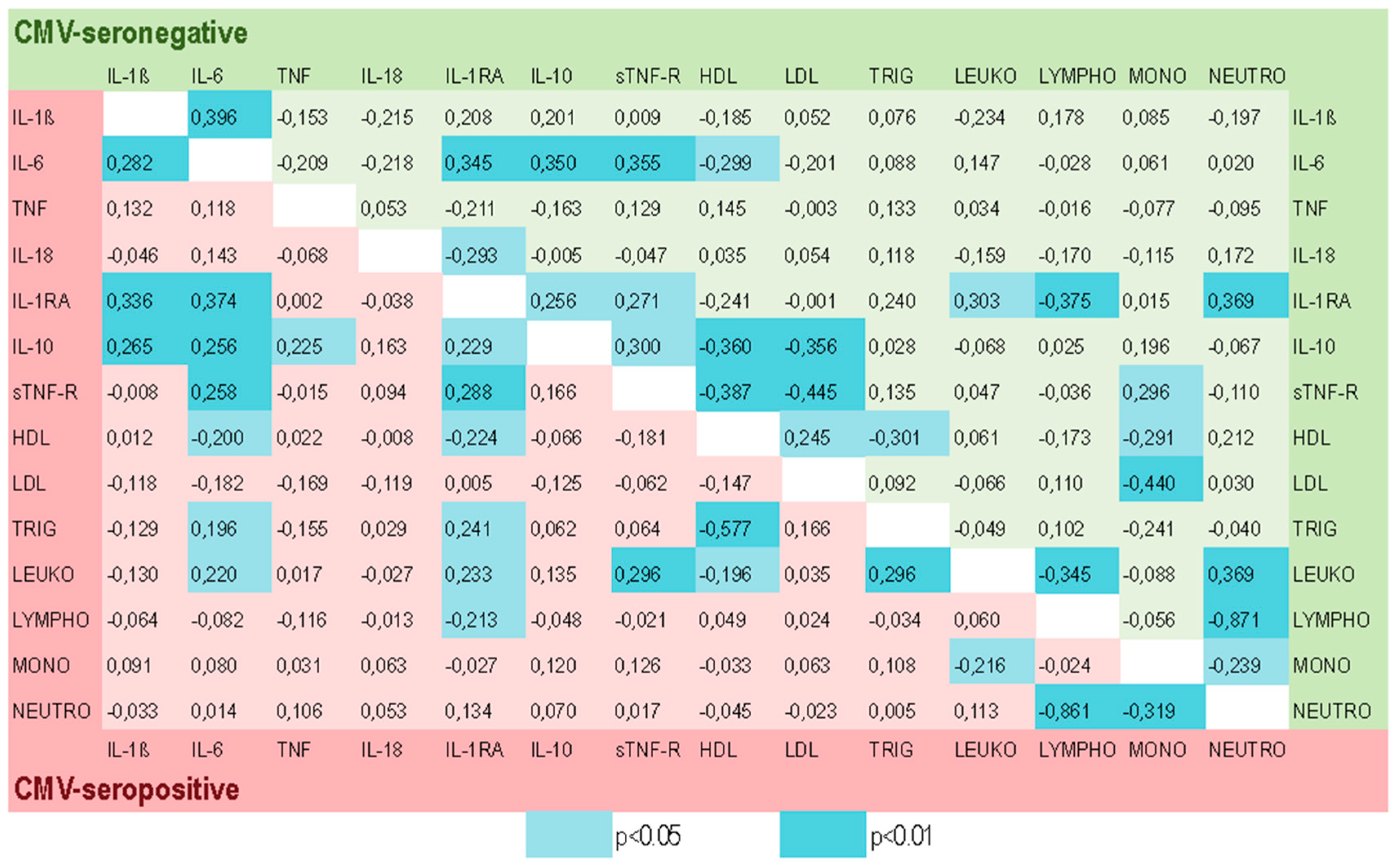
2.4. Associations Among Various Blood Biomarkers in the CMV-Negative and CMV-Positive Groups and Modulatory Effect of CMV-Infection on the Correlation Coefficients
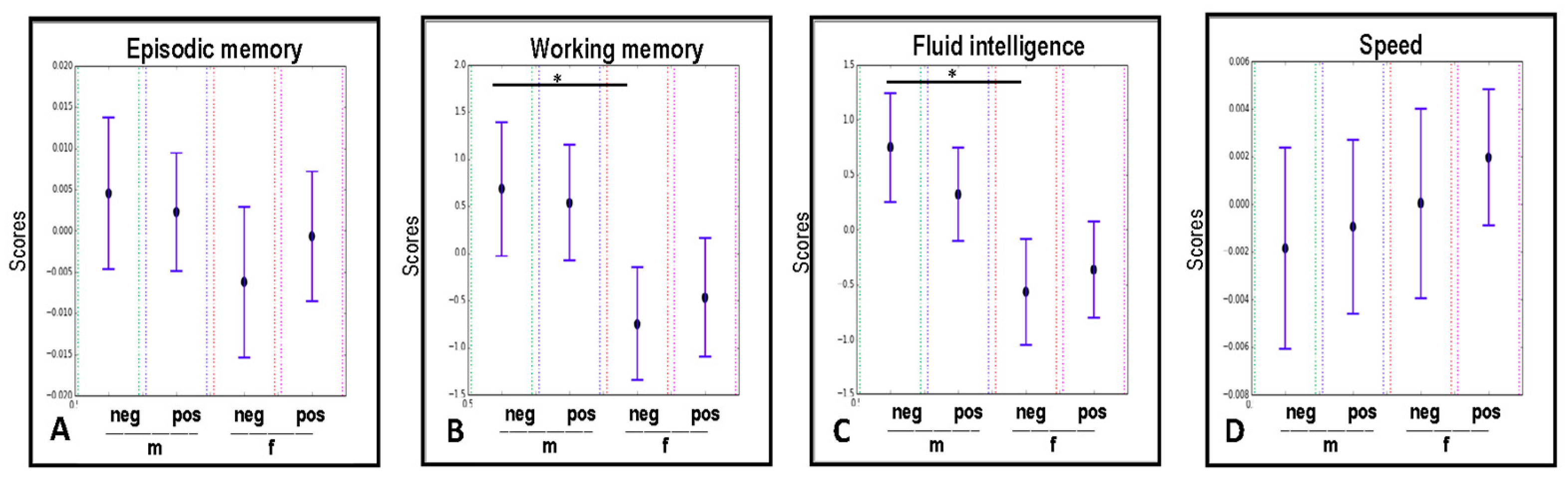
2.5. The Modulatory Effect of the CMV-serostatus and Gender on the Cognitive Abilities of Study Participants
2.6. Associations Between Cognition and Circulating Inflammatory Mediators
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Circulating Biomarkers Assessment
4.2.1. Cytokines TNF, IL-10, IL-6, and IL-1β
4.2.2. sTNF-R, IL-1RA, IL-18 Levels, and CMV-Serostatus
4.3. Cognitive Assessment
4.4. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CMV | Cytomegalovirus |
| IL | Interleukin |
| IL-1RA | Interleukin 1 receptor antagonist |
| TNF | Tumor Necrosis Factor |
| sTNF-R | Soluble Tumor Necrosis Factor receptor |
| HDL | High-density lipoprotein |
| LDL | Low-density lipoprotein |
| IgG | Immunoglobulin G |
| ANOVA | Analysis of variance |
| MANOVA | Multivariate ANOVAs |
| CI | Confidence interval |
| EM | Episodic memory |
| WM | Working memory |
| Gf | Fluid intelligence |
| CFA | Confirmatory Factor Analysis |
| IGF-1 | Insulin-like growth factor 1 |
| ELISA | Enzyme-linked Immunosorbent Assay |
| CBA | Cytometric bead array |
References
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2014, 69 (Suppl. 1), S4–S9. [Google Scholar] [CrossRef] [PubMed]
- Holmes, C.; Cunningham, C.; Zotova, E.; Woolford, J.; Dean, C.; Kerr, S.; Culliford, D.; Perry, V.H. Systemic inflammation and disease progression in Alzheimer disease. Neurology 2009, 73, 768–774. [Google Scholar] [CrossRef] [PubMed]
- King, E.; O’Brien, J.T.; Donaghy, P.; Morris, C.; Barnett, N.; Olsen, K.; Martin-Ruiz, C.; Taylor, J.P.; Thomas, A.J. Peripheral inflammation in prodromal Alzheimer’s and Lewy body dementias. J. Neurol. Neurosurg. Psychiatry 2018, 89, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Garagnani, P.; Vitale, G.; Capri, M.; Salvioli, S. Inflammaging and ‘Garb-aging’. Trends Endocrinol. Metab. 2017, 28, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, S.; Müller, L.; Wenger, E.; Duzel, S.; Pawelec, G. Contribution of neuroinflammation and immunity to brain aging and the mitigating effects of physical and cognitive interventions. Neurosci. Biobehav. Rev. 2017, 75, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.M.; Macedo-de la Concha, L.E.; Pantoja-Meléndez, C.A. Low-grade inflammation and its relation to obesity and chronic degenerative diseases. Revista Médica del Hospital General de México 2017, 80, 101–105. [Google Scholar] [CrossRef]
- Chupel, M.U.; Direito, F.; Furtado, G.E.; Minuzzi, L.G.; Pedrosa, F.M.; Colado, J.C.; Ferreira, J.P.; Filaire, E.; Teixeira, A.M. Strength Training Decreases Inflammation and Increases Cognition and Physical Fitness in Older Women with Cognitive Impairment. Front. Physiol. 2017, 8, 377. [Google Scholar] [CrossRef] [PubMed]
- Fülop, T.; Larbi, A.; Dupuis, G.; Le Page, A.; Frost, E.H.; Cohen, A.A.; Witkowski, J.M.; Franceschi, C. Immunosenescence and Inflamm-Aging As Two Sides of the Same Coin: Friends or Foes? Front. Immunol. 2017, 8, 1960. [Google Scholar] [CrossRef] [PubMed]
- Sawada, M.; Suzumura, A.; Marunouchi, T. TNF alpha induces IL-6 production by astrocytes but not by microglia. Brain Res. 1992, 583, 296–299. [Google Scholar] [CrossRef]
- Singh, T.; Newman, A.B. Inflammatory markers in population studies of aging. Ageing Res. Rev. 2011, 10, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Steensberg, A.; Fischer, C.P.; Keller, C.; Moller, K.; Pedersen, B.K. IL-6 enhances plasma IL-1ra, IL-10, and cortisol in humans. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E433–E437. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Rodriguez, L.; Lopez-Hoyos, M.; Munoz-Cacho, P.; Martinez-Taboada, V.M. Aging is associated with circulating cytokine dysregulation. Cell. Immunol. 2012, 273, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Interleukin-1 and interleukin-1 receptor antagonist. Nutrition 1995, 11, 492–494. [Google Scholar] [PubMed]
- Audet, M.C.; Anisman, H. Interplay between pro-inflammatory cytokines and growth factors in depressive illnesses. Front. Cell. Neurosci. 2013, 7, 68. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, F.; Rossetti, A.C.; Racagni, G.; Gass, P.; Riva, M.A.; Molteni, R. Brain-derived neurotrophic factor: A bridge between inflammation and neuroplasticity. Front. Cell. Neurosci. 2014, 8, 430. [Google Scholar] [CrossRef] [PubMed]
- Lutz, C.T.; Quinn, L.S. Sarcopenia, obesity, and natural killer cell immune senescence in aging: Altered cytokine levels as a common mechanism. Aging 2012, 4, 535–546. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.C.; McCusker, R.H.; Strle, K.; Johnson, R.W.; Dantzer, R.; Kelley, K.W. Regulation of IGF-I function by proinflammatory cytokines: At the interface of immunology and endocrinology. Cell. Immunol. 2008, 252, 91–110. [Google Scholar] [CrossRef] [PubMed]
- Biet, F.; Locht, C.; Kremer, L. Immunoregulatory functions of interleukin 18 and its role in defense against bacterial pathogens. J. Mol. Med. 2002, 80, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.C.; Joshi, S.; Greenwood, H.; Panda, A.; Lord, J.M. Aging of the innate immune system. Curr. Opin. Immunol. 2010, 22, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.M.; Merien, F.; Braakhuis, A.; Dulson, D. T-cells and their cytokine production: The anti-inflammatory and immunosuppressive effects of strenuous exercise. Cytokine 2017, 104, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Arend, W.P.; Malyak, M.; Guthridge, C.J.; Gabay, C. Interleukin-1 receptor antagonist: Role in biology. Annu. Rev. Immunol. 1998, 16, 27–55. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, C.; Diamond, D.J. The immune response to human CMV. Future Virol. 2012, 7, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Hamprecht, K.; Pawelec, G. The role of CMV in “immunosenescence”. In The Ageing Immune System and Health; Bueno, V., Lord, J.M., Jackson, T.A., Eds.; Springer: Cham, Switzerland, 2017; pp. 53–68. [Google Scholar]
- Pawelec, G. Immunosenenescence: Role of cytomegalovirus. Exp. Gerontol. 2014, 54, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Weltevrede, M.; Eilers, R.; de Melker, H.E.; van Baarle, D. Cytomegalovirus persistence and T-cell immunosenescence in people aged fifty and older: A systematic review. Exp. Gerontol. 2016, 77, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Nikolich-Zugich, J.; Goodrum, F.; Knox, K.; Smithey, M.J. Known unknowns: How might the persistent herpesvirome shape immunity and aging? Curr. Opin. Immunol. 2017, 48, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.E.; Redeker, A.; Arens, R.; van Baarle, D.; van den Berg, S.P.H.; Benedict, C.A.; Cicin-Sain, L.; Hill, A.B.; Wills, M.R. CMV immune evasion and manipulation of the immune system with aging. Geroscience 2017, 39, 273–291. [Google Scholar] [CrossRef] [PubMed]
- Al-Attar, A.; Presnell, S.R.; Peterson, C.A.; Thomas, D.T.; Lutz, C.T. Data correlations between gender, cytomegalovirus infection and T cells, NK cells, and soluble immune mediators in elderly humans. Data Brief 2016, 8, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.M.; Glaser, R.; Malarkey, W.B.; Beversdorf, D.Q.; Peng, J.; Kiecolt-Glaser, J.K. Inflammation and reactivation of latent herpesviruses in older adults. Brain Behav. Immun. 2012, 26, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Simanek, A.M.; Cheng, C.; Yolken, R.; Uddin, M.; Galea, S.; Aiello, A.E. Herpesviruses, inflammatory markers and incident depression in a longitudinal study of Detroit residents. Psychoneuroendocrinology 2014, 50, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.C.; Kao, W.H.; Murakami, P.; Xue, Q.L.; Chiou, R.B.; Detrick, B.; McDyer, J.F.; Semba, R.D.; Casolaro, V.; Walston, J.D.; et al. Cytomegalovirus infection and the risk of mortality and frailty in older women: A prospective observational cohort study. Am. J. Epidemiol. 2010, 171, 1144–1152. [Google Scholar] [CrossRef] [PubMed]
- Kilgour, A.H.; Firth, C.; Harrison, R.; Moss, P.; Bastin, M.E.; Wardlaw, J.M.; Deary, I.J.; Starr, J.M. Seropositivity for CMV and IL-6 levels are associated with grip strength and muscle size in the elderly. Immun. Ageing 2013, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Barnes, L.L.; Capuano, A.W.; Aiello, A.E.; Turner, A.D.; Yolken, R.H.; Torrey, E.F.; Bennett, D.A. Cytomegalovirus infection and risk of Alzheimer disease in older black and white individuals. J. Infect. Dis. 2015, 211, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Mathei, C.; Vaes, B.; Wallemacq, P.; Degryse, J. Associations between cytomegalovirus infection and functional impairment and frailty in the BELFRAIL Cohort. J. Am. Geriatr. Soc. 2011, 59, 2201–2208. [Google Scholar] [CrossRef] [PubMed]
- Aiello, A.E.; Haan, M.; Blythe, L.; Moore, K.; Gonzalez, J.M.; Jagust, W. The influence of latent viral infection on rate of cognitive decline over 4 years. J. Am. Geriatr. Soc. 2006, 54, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Gow, A.J.; Firth, C.M.; Harrison, R.; Starr, J.M.; Moss, P.; Deary, I.J. Cytomegalovirus infection and cognitive abilities in old age. Neurobiol. Aging 2013, 34, 1846–1852. [Google Scholar] [CrossRef] [PubMed]
- Ricci, G.; Pirillo, I.; Tomassoni, D.; Sirignano, A.; Grappasonni, I. Metabolic syndrome, hypertension, and nervous system injury: Epidemiological correlates. Clin. Exp. Hypertens 2017, 39, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Raz, N.; Yang, Y.; Dahle, C.L.; Land, S. Volume of white matter hyperintensities in healthy adults: Contribution of age, vascular risk factors, and inflammation-related genetic variants. Biochim. Biophys. Acta 2012, 1822, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzi, V.; Scafato, E.; Capurso, C.; D’Introno, A.; Colacicco, A.M.; Frisardi, V.; Vendemiale, G.; Baldereschi, M.; Crepaldi, G.; Di Carlo, A.; et al. Metabolic syndrome, mild cognitive impairment, and progression to dementia. The Italian Longitudinal Study on Aging. Neurobiol. Aging 2011, 32, 1932–1941. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.P.; Feng, L.; Nyunt, M.S.; Feng, L.; Gao, Q.; Lim, M.L.; Collinson, S.L.; Chong, M.S.; Lim, W.S.; Lee, T.S.; et al. Metabolic Syndrome and the Risk of Mild Cognitive Impairment and Progression to Dementia: Follow-up of the Singapore Longitudinal Ageing Study Cohort. JAMA Neurol. 2016, 73, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Freeman, R.B., Jr. The ‘indirect’ effects of cytomegalovirus infection. Am. J. Transplant. 2009, 9, 2453–2458. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Peng, G.; Bai, J.; He, B.; Huang, K.; Hu, X.; Liu, D. Cytomegalovirus Infection and Relative Risk of Cardiovascular Disease (Ischemic Heart Disease, Stroke, and Cardiovascular Death): A Meta-Analysis of Prospective Studies Up to 2016. J. Am. Heart Assoc. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Caruso, C.; Accardi, G.; Virruso, C.; Candore, G. Sex, gender and immunosenescence: A key to understand the different lifespan between men and women? Immun. Ageing 2013, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Klein, R.S. Sex Drives Dimorphic Immune Responses to Viral Infections. J. Immunol. 2017, 198, 1782–1790. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Bouman, A.; Schipper, M.; Heineman, M.J.; Faas, M.M. Gender difference in the non-specific and specific immune response in humans. Am. J. Reprod. Immunol. 2004, 52, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Garcia Verdecia, B.; Saavedra Hernandez, D.; Lorenzo-Luaces, P.; de Jesus Badia Alvarez, T.; Leonard Rupale, I.; Mazorra Herrera, Z.; Crombet Ramos, T.; Lage Davila, A. Immunosenescence and gender: A study in healthy Cubans. Immun. Ageing 2013, 10, 16. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, K.; Utsuyama, M.; Hayashi, Y.; Kitagawa, M.; Makinodan, T.; Fulop, T. Slower immune system aging in women versus men in the Japanese population. Immun. Ageing 2013, 10, 19. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Greer, J.M.; Hull, R.; O’Sullivan, J.D.; Henderson, R.D.; Read, S.J.; McCombe, P.A. The effect of ageing on human lymphocyte subsets: Comparison of males and females. Immun. Ageing 2010, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, S.; Derhovanessian, E.; Steinhagen-Thiessen, E.; Goldeck, D.; Muller, L.; Pawelec, G. Impact of age, sex and CMV-infection on peripheral T cell phenotypes: Results from the Berlin BASE-II Study. Biogerontology 2015, 16, 631–643. [Google Scholar] [CrossRef] [PubMed]
- van der Heiden, M.; van Zelm, M.C.; Bartol, S.J.; de Rond, L.G.; Berbers, G.A.; Boots, A.M.; Buisman, A.M. Differential effects of Cytomegalovirus carriage on the immune phenotype of middle-aged males and females. Sci. Rep. 2016, 6, 26892. [Google Scholar] [CrossRef] [PubMed]
- Althouse, A.D. Adjust for Multiple Comparisons? It’s Not That Simple. Ann. Thorac. Surg. 2016, 101, 1644–1645. [Google Scholar] [CrossRef] [PubMed]
- Goetzl, E.J.; Huang, M.C.; Kon, J.; Patel, K.; Schwartz, J.B.; Fast, K.; Ferrucci, L.; Madara, K.; Taub, D.D.; Longo, D.L. Gender specificity of altered human immune cytokine profiles in aging. FASEB J. 2010, 24, 3580–3589. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, M.P.; Meydani, M.; Lichtenstein, A.H.; Schaefer, E.J.; Dillard, A.; Lamon-Fava, S. Sex hormone modulation of proinflammatory cytokine and C-reactive protein expression in macrophages from older men and postmenopausal women. J. Endocrinol. 2010, 206, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Malkin, C.J.; Pugh, P.J.; Jones, R.D.; Kapoor, D.; Channer, K.S.; Jones, T.H. The effect of testosterone replacement on endogenous inflammatory cytokines and lipid profiles in hypogonadal men. J. Clin. Endocrinol. Metab. 2004, 89, 3313–3318. [Google Scholar] [CrossRef] [PubMed]
- Accardi, G.; Caruso, C. Immune-inflammatory responses in the elderly: An update. Immun. Ageing 2018, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Al-Attar, A.; Presnell, S.R.; Peterson, C.A.; Thomas, D.T.; Lutz, C.T. The effect of sex on immune cells in healthy aging: Elderly women have more robust natural killer lymphocytes than do elderly men. Mech. Ageing Dev. 2016, 156, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Gubbels Bupp, M.R. Sex, the aging immune system, and chronic disease. Cell. Immunol. 2015, 294, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Gubbels Bupp, M.R.; Potluri, T.; Fink, A.L.; Klein, S.L. The Confluence of Sex Hormones and Aging on Immunity. Front. Immunol. 2018, 9, 1269. [Google Scholar] [CrossRef] [PubMed]
- Ostan, R.; Monti, D.; Gueresi, P.; Bussolotto, M.; Franceschi, C.; Baggio, G. Gender, aging and longevity in humans: An update of an intriguing/neglected scenario paving the way to a gender-specific medicine. Clin. Sci. 2016, 130, 1711–1725. [Google Scholar] [CrossRef] [PubMed]
- Villacres, M.C.; Longmate, J.; Auge, C.; Diamond, D.J. Predominant type 1 CMV-specific memory T-helper response in humans: Evidence for gender differences in cytokine secretion. Hum. Immunol. 2004, 65, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Kovats, S. Estrogen receptors regulate innate immune cells and signaling pathways. Cell. Immunol. 2015, 294, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Nakagomi, A.; Seino, Y.; Noma, S.; Kohashi, K.; Kosugi, M.; Kato, K.; Kusama, Y.; Atarashi, H.; Shimizu, W. Relationships between the Serum Cholesterol Levels, Production of Monocyte Proinflammatory Cytokines and Long-term Prognosis in Patients with Chronic Heart Failure. Intern. Med. 2014, 53, 2415–2424. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R.; Grunfeld, C. Role of cytokines in inducing hyperlipidemia. Diabetes 1992, 41 (Suppl. 2), 97–101. [Google Scholar] [CrossRef] [PubMed]
- Odegaard, J.I.; Chawla, A. Old HDL learns a new (anti-inflammatory) trick. Nat. Immunol. 2014, 15, 138–139. [Google Scholar] [CrossRef] [PubMed]
- Khovidhunkit, W.; Kim, M.S.; Memon, R.A.; Shigenaga, J.K.; Moser, A.H.; Feingold, K.R.; Grunfeld, C. Effects of infection and inflammation on lipid and lipoprotein metabolism: Mechanisms and consequences to the host. J. Lipid Res. 2004, 45, 1169–1196. [Google Scholar] [CrossRef] [PubMed]
- Catapano, A.L.; Pirillo, A.; Norata, G.D. Vascular inflammation and low-density lipoproteins: Is cholesterol the link? A lesson from the clinical trials. Br. J. Pharmacol. 2017, 174, 3973–3985. [Google Scholar] [CrossRef] [PubMed]
- Orsini, A.; Chiacchio, L.; Cinque, M.; Cocchiaro, C.; Schiappa, O.; Grossi, D. Effects of age, education and sex on two tests of immediate memory: A study of normal subjects from 20 to 99 years of age. Percept. Mot. Skills 1986, 63, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, W.; Wang, L.; Zhang, X. Harvard Aging Brain Study. Gender differences of neuropsychological profiles in cognitively normal older people without amyloid pathology. Compr. Psychiatry 2017, 75, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Munro, C.A.; Winicki, J.M.; Schretlen, D.J.; Gower, E.W.; Turano, K.A.; Munoz, B.; Keay, L.; Bandeen-Roche, K.; West, S.K. Sex differences in cognition in healthy elderly individuals. Neuropsychol. Dev. Cogn. B Aging Neuropsychol. Cogn. 2012, 19, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Al-Delaimy, W.K.; von Muhlen, D.; Barrett-Connor, E. Insulinlike growth factor-1, insulinlike growth factor binding protein-1, and cognitive function in older men and women. J. Am. Geriatr. Soc. 2009, 57, 1441–1446. [Google Scholar] [CrossRef] [PubMed]
- Perice, L.; Barzilai, N.; Verghese, J.; Weiss, E.F.; Holtzer, R.; Cohen, P.; Milman, S. Lower circulating insulin-like growth factor-I is associated with better cognition in females with exceptional longevity without compromise to muscle mass and function. Aging 2016, 8, 2414–2424. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.J.; Finch, C.E.; Cohen, H.J. Cytokines and cognition--the case for a head-to-toe inflammatory paradigm. J. Am. Geriatr. Soc. 2002, 50, 2041–2056. [Google Scholar] [CrossRef] [PubMed]
- Shields, G.S.; Moons, W.G.; Slavich, G.M. Inflammation, Self-Regulation, and Health: An Immunologic Model of Self-Regulatory Failure. Perspect. Psychol. Sci. 2017, 12, 588–612. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.T.; Collins, J.S.; Wiener, H.; Acton, R.; Go, R.C. The role of TNF and its receptors in Alzheimer’s disease. Neurobiol. Aging 2001, 22, 873–883. [Google Scholar] [CrossRef]
- Fiore, M.; Angelucci, F.; Alleva, E.; Branchi, I.; Probert, L.; Aloe, L. Learning performances, brain NGF distribution and NPY levels in transgenic mice expressing TNF-alpha. Behav. Brain Res. 2000, 112, 165–175. [Google Scholar] [CrossRef]
- Rizzo, F.R.; Musella, A.; De Vito, F.; Fresegna, D.; Bullitta, S.; Vanni, V.; Guadalupi, L.; Stampanoni Bassi, M.; Buttari, F.; Mandolesi, G.; et al. Tumor Necrosis Factor and Interleukin-1beta Modulate Synaptic Plasticity during Neuroinflammation. Neural Plast. 2018, 2018, 8430123. [Google Scholar] [CrossRef] [PubMed]
- Yirmiya, R.; Goshen, I. Immune modulation of learning, memory, neural plasticity and neurogenesis. Brain Behav. Immun. 2011, 25, 181–213. [Google Scholar] [CrossRef] [PubMed]
- Lobo-Silva, D.; Carriche, G.M.; Castro, A.G.; Roque, S.; Saraiva, M. Balancing the immune response in the brain: IL-10 and its regulation. J. Neuroinflamm. 2016, 13, 297. [Google Scholar] [CrossRef] [PubMed]
- Morrisette-Thomas, V.; Cohen, A.A.; Fulop, T.; Riesco, E.; Legault, V.; Li, Q.; Milot, E.; Dusseault-Belanger, F.; Ferrucci, L. Inflamm-aging does not simply reflect increases in pro-inflammatory markers. Mech. Ageing Dev. 2014, 139, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Tegeler, C.; O’Sullivan, J.L.; Bucholtz, N.; Goldeck, D.; Pawelec, G.; Steinhagen-Thiessen, E.; Demuth, I. The inflammatory markers CRP, IL-6, and IL-10 are associated with cognitive function--data from the Berlin Aging Study II. Neurobiol. Aging 2016, 38, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Marsland, A.L.; Gianaros, P.J.; Abramowitch, S.M.; Manuck, S.B.; Hariri, A.R. Interleukin-6 covaries inversely with hippocampal grey matter volume in middle-aged adults. Biol. Psychiatry 2008, 64, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, A.L.; Massaro, J.M.; Beiser, A.S.; Seshadri, S.; Larson, M.G.; Wolf, P.A.; Au, R.; Benjamin, E.J. Inflammatory markers and neuropsychological functioning: The Framingham Heart Study. Neuroepidemiology 2011, 37, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.E.; Hodkinson, C.F.; Maylor, E.A.; McCormack, J.M.; Rae, G.; Strain, S.; Alexander, H.D.; Wallace, J.M. Intracellular cytokine production and cognition in healthy older adults. Psychoneuroendocrinology 2013, 38, 2196–2208. [Google Scholar] [CrossRef] [PubMed]
- Trompet, S.; de Craen, A.J.; Slagboom, P.; Shepherd, J.; Blauw, G.J.; Murphy, M.B.; Bollen, E.L.; Buckley, B.M.; Ford, I.; Gaw, A.; et al. Genetic variation in the interleukin-1 beta-converting enzyme associates with cognitive function. The PROSPER study. Brain 2008, 131, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Whiting, C.C.; Siebert, J.; Newman, A.M.; Du, H.W.; Alizadeh, A.A.; Goronzy, J.; Weyand, C.M.; Krishnan, E.; Fathman, C.G.; Maecker, H.T. Large-Scale and Comprehensive Immune Profiling and Functional Analysis of Normal Human Aging. PLoS ONE 2015, 10, e0133627. [Google Scholar] [CrossRef] [PubMed]
- de Jager, W.; Bourcier, K.; Rijkers, G.T.; Prakken, B.J.; Seyfert-Margolis, V. Prerequisites for cytokine measurements in clinical trials with multiplex immunoassays. BMC Immunol. 2009, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Goldeck, D.; Pawelec, G.; Norman, K.; Steinhagen-Thiessen, E.; Oettinger, L.; Haehnel, K.; Demuth, I. No strong correlations between serum cytokine levels, CMV serostatus and hand-grip strength in older subjects in the Berlin BASE-II cohort. Biogerontology 2016, 17, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Kern, P.A.; Ranganathan, S.; Li, C.; Wood, L.; Ranganathan, G. Adipose tissue tumor necrosis factor and interleukin-6 expression in human obesity and insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E745–E751. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.; Ward, G. Interferences in immunoassay. Clin. Biochem. Rev. 2004, 25, 105–120. [Google Scholar] [PubMed]
- Rios-Lugo, M.J.; Martin, C.; Alarcon, J.A.; Esquifino, A.; Barbieri, G.; Solano, P.; Sanz, M. Optimization of buffer solutions to analyze inflammatory cytokines in gingival crevicular fluid by multiplex flow cytometry. Med. Oral. Patol. Oral Cir. Bucal 2015, 20, e13–e16. [Google Scholar] [CrossRef] [PubMed]
- Chavance, M.; Escolano, S.; Romon, M.; Basdevant, A.; de Lauzon-Guillain, B.; Charles, M.A. Latent variables and structural equation models for longitudinal relationships: An illustration in nutritional epidemiology. BMC Med. Res. Methodol. 2010, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Düzel, S.; Buchmann, N.; Drewelies, J.; Gerstorf, D.; Lindenberger, U.; Steinhagen-Thiessen, E.; Norman, K.; Demuth, I. Validation of a single factor representing the indicators of metabolic syndrome as a continuous measure of metabolic load and its association with health and cognitive function. PLoS ONE 2018, 13, e0208231. [Google Scholar] [CrossRef] [PubMed]
- Helmstaedter, C.; Durwen, H.F. The Verbal Learning and Retention Test. A useful and differentiated tool in evaluating verbal memory performance. Schweiz Arch. Neurol. Psychiatr. (1985) 1990, 141, 21–30. [Google Scholar]
- Lindenberger, U.; Mayr, U.; Kliegl, R. Speed and intelligence in old age. Psychol. Aging 1993, 8, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Schmiedek, F.; Lovden, M.; Lindenberger, U. Hundred Days of Cognitive Training Enhance Broad Cognitive Abilities in Adulthood: Findings from the COGITO Study. Front. Aging Neurosci. 2010, 2. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, M.M.; Verma, D.K. Exposure estimation in the presence of nondetectable values: Another look. AIHAJ 2001, 62, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Hewett, P.; Ganser, G.H. A comparison of several methods for analyzing censored data. Ann. Occup. Hyg. 2007, 51, 611–632. [Google Scholar] [CrossRef] [PubMed]
- Ogden, T.L. Handling results below the level of detection. Ann. Occup. Hyg. 2010, 54, 255–256. [Google Scholar] [CrossRef] [PubMed]
- Hesterberg, T.; Monaghan, S.; Moore, D.S.; Clipson, A.; Epstein, R. Bootstrap Methods and Permutation Tests; W.H. Freeman and Company: New York, NY, USA, 2003. [Google Scholar]
- Steiger, J.H. Testing Pattern Hypotheses On Correlation Matrices: Alternative Statistics And Some Empirical Results. Multivar. Behav. Res. 1980, 15, 335–352. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | r | p-Value |
|---|---|---|
| CMV-negative group (n = 59): | ||
| EM 1 vs. TNF | −0.330 | 0.010 |
| EM vs. IL-10 | 0.282 | 0.029 |
| WM vs. TNF | −0.334 | 0.009 |
| Gf vs. TNF | −0.415 | 0.001 |
| CMV-positive group (n = 102): | ||
| EM vs. IL-6 | −0.201 | 0.042 |
| EM vs. IL-1RA | −0.214 | 0.030 |
| WM vs. IL-1RA | −0.218 | 0.028 |
| Gf vs. IL-6 | −0.205 | 0.039 |
| Gf vs. IL-RA | −0.272 | 0.005 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Benedetto, S.; Gaetjen, M.; Müller, L. The Modulatory Effect of Gender and Cytomegalovirus-Seropositivity on Circulating Inflammatory Factors and Cognitive Performance in Elderly Individuals. Int. J. Mol. Sci. 2019, 20, 990. https://doi.org/10.3390/ijms20040990
Di Benedetto S, Gaetjen M, Müller L. The Modulatory Effect of Gender and Cytomegalovirus-Seropositivity on Circulating Inflammatory Factors and Cognitive Performance in Elderly Individuals. International Journal of Molecular Sciences. 2019; 20(4):990. https://doi.org/10.3390/ijms20040990
Chicago/Turabian StyleDi Benedetto, Svetlana, Marcel Gaetjen, and Ludmila Müller. 2019. "The Modulatory Effect of Gender and Cytomegalovirus-Seropositivity on Circulating Inflammatory Factors and Cognitive Performance in Elderly Individuals" International Journal of Molecular Sciences 20, no. 4: 990. https://doi.org/10.3390/ijms20040990
APA StyleDi Benedetto, S., Gaetjen, M., & Müller, L. (2019). The Modulatory Effect of Gender and Cytomegalovirus-Seropositivity on Circulating Inflammatory Factors and Cognitive Performance in Elderly Individuals. International Journal of Molecular Sciences, 20(4), 990. https://doi.org/10.3390/ijms20040990