Zebrafish Models of Neurodevelopmental Disorders: Limitations and Benefits of Current Tools and Techniques


Abstract
:1. Introduction
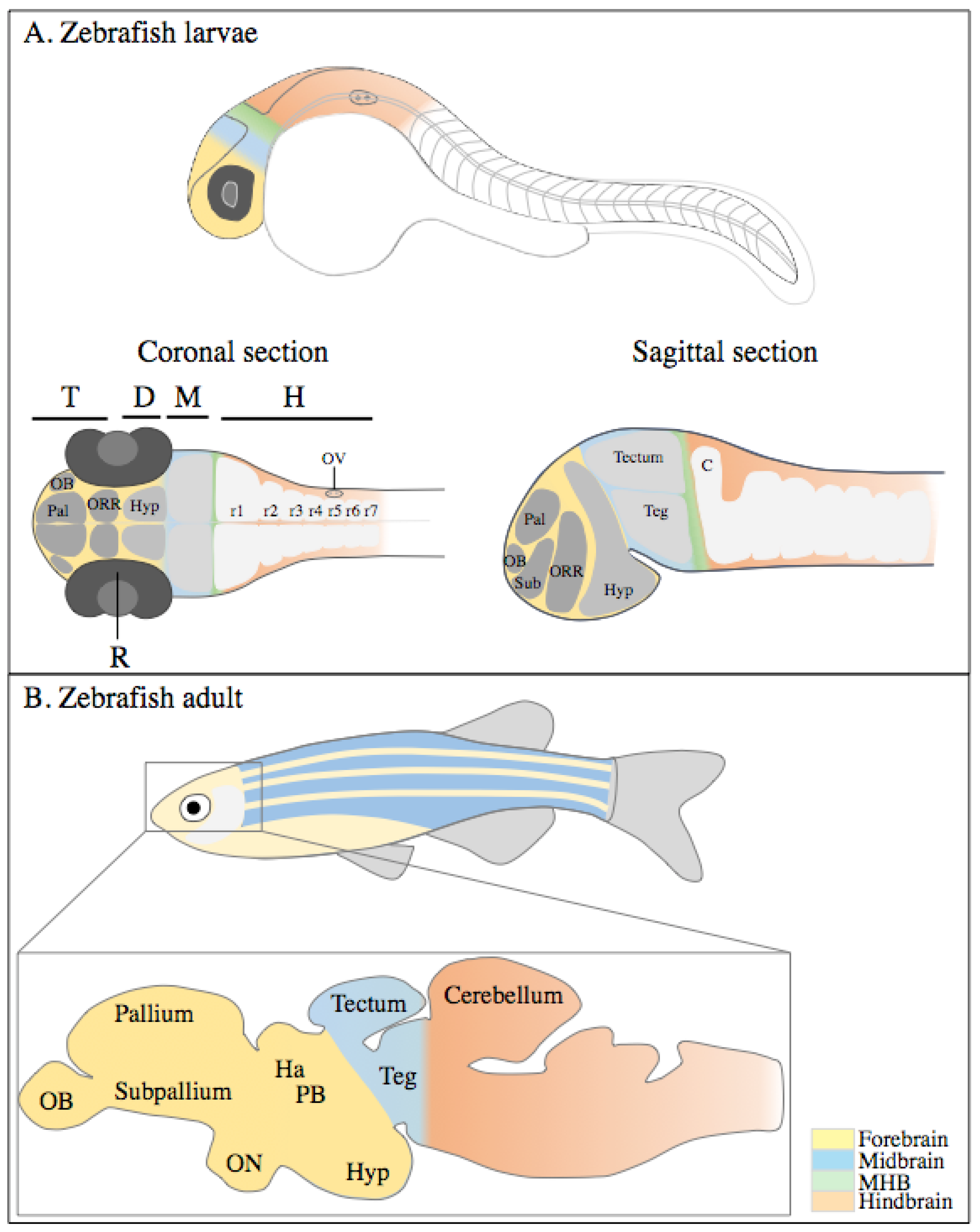
2. Zebrafish Central Nervous System Development and Organization
2.1. CNS Morphogenesis
2.2. Neuronal Subclasses
3. Genetic Tools for Investigating Neuronal Development and Function
3.1. Knockdown Techniques
3.2. Transgene Overexpression
3.3. Stable Mutagenesis
4. Assays
4.1. Cellular Characterization
4.2. Behavior Characterization
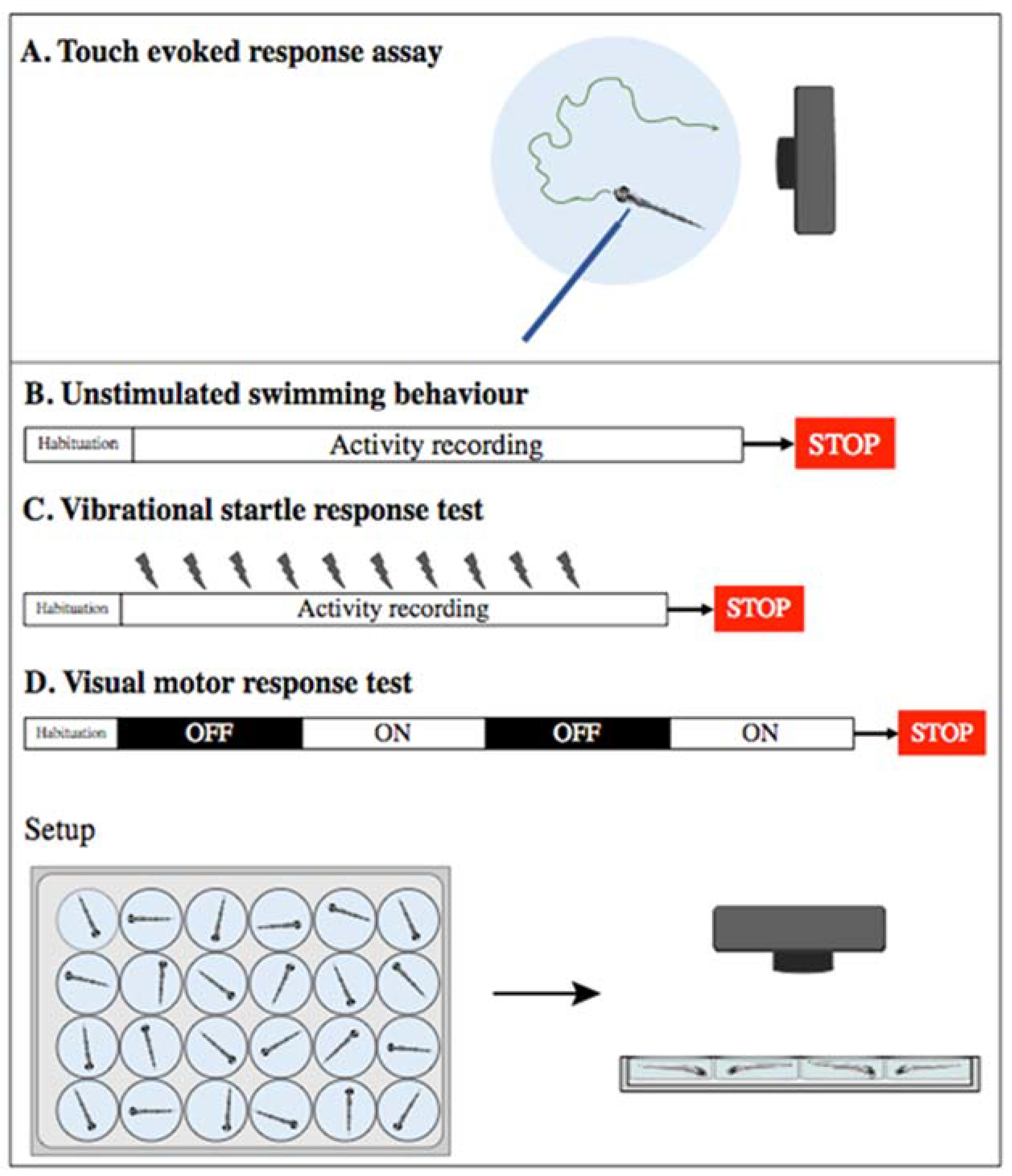
4.2.1. Larvae Assays
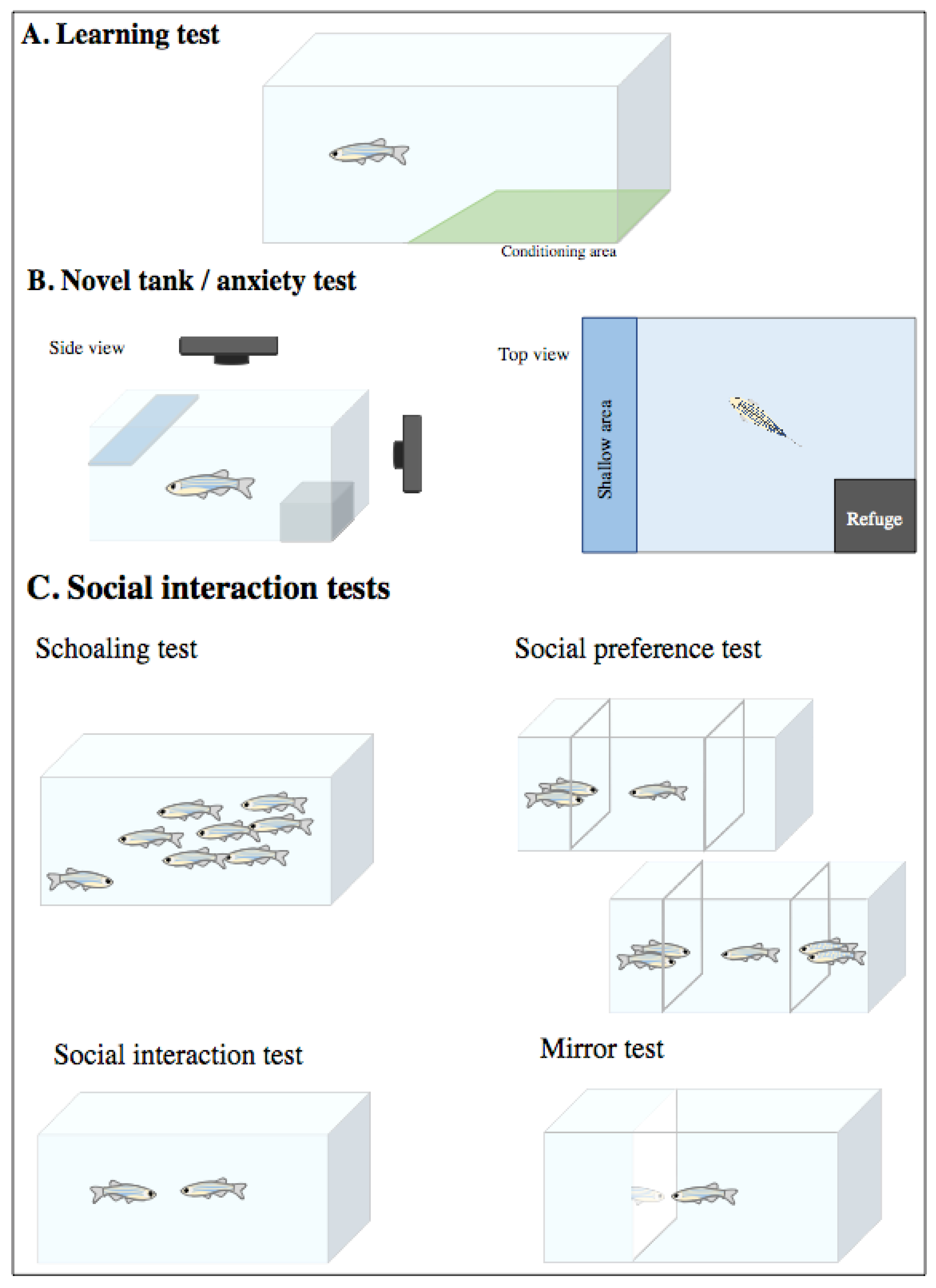
4.2.2. Adult Assays
Learning Tests
Anxiety/Fear Tests
Social Interaction Tests
5. Zebrafish Models of Human Neurodevelopmental Disorders
6. Therapies
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADHD | attention deficit hyperactivity disorder |
| ASD | autism spectrum disorder |
| C | cerebellum |
| CAM | cameleon |
| CNS | central nervous system |
| CNV | copy number variation |
| CRISPR | clustered regularly interspaced short palindromic repeats |
| D | diencephalon |
| DD | developmental delay |
| DPF | days post fertilization |
| GFP | green fluorescent protein |
| GLRA2 | glycine receptor alpha 2 |
| GlyR | glycine receptor |
| ID | intellectual disability |
| M | midbrain |
| MHB | midbrain-hindbrain boundary |
| MO | morpholino |
| MPEP | 2-methyl-6-(phenylethynyl)pyridine |
| MPH | methylphenidate |
| H: | hindbrain |
| HA | habenula |
| HPF | hours post fertilization |
| HYP | hypothalamus |
| OB | olfactory bulb |
| ON | optic nerve |
| ORR | optic recess region |
| OV | otic vesicle |
| PAL | pallium |
| PB | pineal body |
| PFOS | perfluorooctane sulfonate |
| PTZ | pentylenetetrazol |
| R | retina |
| RFP | red fluorescent protein |
| r1–r7 | rhombomeres 1 to 7 |
| SUB | sub-pallium |
| T | telencephalon |
| TEG | tegmentum |
| UV | ultra violet |
| VPA | valproic acid |
| YFP | yellow fluorescent protein |
References
- Happé, F.; Ronald, A. The ‘Fractionable Autism Triad’: A Review of Evidence from Behavioural, Genetic, Cognitive and Neural Research. Neuropsychol. Rev. 2008, 18, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Couture, J. A Review of the Pathophysiology, Etiology, and Treatment of Attention-Deficit Hyperactivity Disorder (ADHD). Ann. Pharmacother. 2014, 48, 209–225. [Google Scholar] [CrossRef] [PubMed]
- McDougle, C.J.; Erickson, C.A.; Stigler, K.A.; Posey, D.J. Neurochemistry in the Pathophysiology of Autism. J. Clin. Psychiatry 2005, 66 (Suppl. 10), 9–18. [Google Scholar]
- Geschwind, D.H. Genetics of autism spectrum disorders. Trends Cogn. Sci. 2011, 15, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Norton, W.H.J. Toward developmental models of psychiatric disorders in zebrafish. Front. Neural Circuits 2013, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pilorge, M.; Fassier, C.; Le Corronc, H.; Potey, A.; Bai, J.; De Gois, S.; Delaby, E.; Assouline, B.; Guinchat, V.; Devillard, F.; et al. Genetic and functional analyses demonstrate a role for abnormal glycinergic signaling in autism. Mol. Psychiatry 2016, 21, 936–945. [Google Scholar] [CrossRef]
- Rubia, K. “Cool” Inferior Frontostriatal Dysfunction in Attention-Deficit/Hyperactivity Disorder Versus “Hot” Ventromedial Orbitofrontal-Limbic Dysfunction in Conduct Disorder: A Review. Biol. Psychiatry 2011, 69, e69–e87. [Google Scholar] [CrossRef]
- Kozol, R.A.; Abrams, A.J.; James, D.M.; Buglo, E.; Yan, Q.; Dallman, J.E. Function Over Form: Modeling Groups of Inherited Neurological Conditions in Zebrafish. Front. Mol. Neurosci. 2016, 9, 1–15. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Chakraborty, C.; Hsu, C.; Wen, Z.; Lin, C.; Agoramoorthy, G. Zebrafish: A Complete Animal Model for In Vivo Drug Discovery and Development. Curr. Drug Metab. 2009, 10, 116–124. [Google Scholar] [CrossRef]
- Ali, S.; Champagne, D.L.; Richardson, M.K. Behavioral profiling of zebrafish embryos exposed to a panel of 60 water-soluble compounds. Behav. Brain Res. 2012, 228, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Woo, K.; Fraser, S.E. Order and coherence in the fate map of the zebrafish nervous system. Development 1995, 121, 2595–2609. [Google Scholar] [PubMed]
- Friedrich, R.W.; Korsching, S.I. Combinatorial and Chemotopic Odorant Coding in the Zebrafish Olfactory Bulb Visualized by Optical Imaging. Neuron 1997, 18, 737–752. [Google Scholar] [CrossRef]
- Miyasaka, N.; Morimoto, K.; Tsubokawa, T.; Higashijima, S.-i.; Okamoto, H.; Yoshihara, Y. From the Olfactory Bulb to Higher Brain Centers: Genetic Visualization of Secondary Olfactory Pathways in Zebrafish. J. Neurosci. 2009, 29, 4756–4767. [Google Scholar] [CrossRef]
- Shinozuka, K.; Watanabe, S. Effects of telencephalic ablation on shoaling behavior in goldfish. Physiol. Behav. 2004, 81, 141–148. [Google Scholar] [CrossRef]
- Teles, M.C.; Cardoso, S.D.; Oliveira, R.F. Social Plasticity Relies on Different Neuroplasticity Mechanisms across the Brain Social Decision-Making Network in Zebrafish. Front. Behav. Neurosci. 2016, 10, 1–12. [Google Scholar] [CrossRef]
- Stednitz, S.J.; McDermott, E.M.; Ncube, D.; Tallafuss, A.; Eisen, J.S.; Washbourne, P. Forebrain Control of Behaviorally Driven Social Orienting in Zebrafish. Curr. Biol. 2018, 28, 2445–2451.e3. [Google Scholar] [CrossRef]
- Mueller, T. What is the Thalamus in Zebrafish? Front. Neurosci. 2012, 6, 1–14. [Google Scholar] [CrossRef]
- Lin, Q.; Jesuthasan, S. Masking of a circadian behavior in larval zebrafish involves the thalamo-habenula pathway. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Wullimann, M.F. Secondary neurogenesis and telencephalic organization in zebrafish and mice: A brief review. Integr. Zool. 2009, 4, 123–133. [Google Scholar] [CrossRef]
- Mueller, T.; Dong, Z.; Berberoglu, M.A.; Guo, S. The dorsal pallium in zebrafish, Danio rerio (Cyprinidae, Teleostei). Brain Res. 2011, 1381, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.-K.; Jesuthasan, S.J.; Penney, T.B. Zebrafish forebrain and temporal conditioning. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20120462. [Google Scholar] [CrossRef] [PubMed]
- Korzh, V. Development of brain ventricular system. Cell. Mol. Life Sci. 2018, 75, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Karlstrom, R. Genetic analysis of axon guidance and mapping in the zebrafish. Trends Neurosci. 1997, 20, 3–8. [Google Scholar] [CrossRef]
- Portugues, R.; Engert, F. The neural basis of visual behaviors in the larval zebrafish. Curr. Opin. Neurobiol. 2009, 19, 644–647. [Google Scholar] [CrossRef] [PubMed]
- Kita, E.M.; Scott, E.K.; Goodhill, G.J. Topographic wiring of the retinotectal connection in zebrafish: Retinotectal Connection in Zebrafish. Dev. Neurobiol. 2015, 75, 542–556. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Li, X.; Du, J. Optic tectal superficial interneurons detect motion in larval zebrafish. Protein Cell 2018. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, J.P.; Trivedi, C.A.; Maurer, C.M.; Ryu, S.; Bollmann, J.H. Layer-Specific Targeting of Direction-Selective Neurons in the Zebrafish Optic Tectum. Neuron 2012, 76, 1147–1160. [Google Scholar] [CrossRef]
- Galant, S.; Furlan, G.; Coolen, M.; Dirian, L.; Foucher, I.; Bally-Cuif, L. Embryonic origin and lineage hierarchies of the neural progenitor subtypes building the zebrafish adult midbrain. Dev. Biol. 2016, 420, 120–135. [Google Scholar] [CrossRef]
- Lodge, D.J.; Grace, A.A. The laterodorsal tegmentum is essential for burst firing of ventral tegmental area dopamine neurons. Proc. Natl. Acad. Sci. USA 2006, 103, 5167–5172. [Google Scholar] [CrossRef]
- Redila, V.; Kinzel, C.; Jo, Y.S.; Puryear, C.B.; Mizumori, S.J.Y. A role for the lateral dorsal tegmentum in memory and decision neural circuitry. Neurobiol. Learn. Mem. 2015, 117, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.; Heisenberg, C.-P.; Jiang, Y.-J.; Beuchle, D.; Lun, K.; Furutani-Seiki, M.; Granato, M.; Haffter, P.; Hammerschmidt, M.; Kane, D.A.; et al. Mutations in zebrafish genes affecting the formation of the boundary between midbrain and hindbrain. Development 1996, 123, 179–190. [Google Scholar] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Moens, C.B.; Prince, V.E. Constructing the hindbrain: Insights from the zebrafish. Dev. Dyn. 2002, 224, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Oxtoby, E.; Jowett, T. Cloning of the zebrafish krox-20 gene (krx-20) and its expression during hindbrain development. Nucleic Acids Res. 1993, 21, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Köster, R.W.; Fraser, S.E. Direct imaging of in vivo neuronal migration in the developing cerebellum. Curr. Biol. 2001, 11, 1858–1863. [Google Scholar] [CrossRef]
- Bae, Y.-K.; Kani, S.; Shimizu, T.; Tanabe, K.; Nojima, H.; Kimura, Y.; Higashijima, S.; Hibi, M. Anatomy of zebrafish cerebellum and screen for mutations affecting its development. Dev. Biol. 2009, 330, 406–426. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, R.; Botez, M.I. The cerebellum and learning processes in animals. Brain Res. Rev. 1990, 15, 325–332. [Google Scholar] [CrossRef]
- Rodríguez, F.; Durán, E.; Gómez, A.; Ocaña, F.M.; Álvarez, E.; Jiménez-Moya, F.; Broglio, C.; Salas, C. Cognitive and emotional functions of the teleost fish cerebellum. Brain Res. Bull. 2005, 66, 365–370. [Google Scholar] [CrossRef]
- Hibi, M.; Matsuda, K.; Takeuchi, M.; Shimizu, T.; Murakami, Y. Evolutionary mechanisms that generate morphology and neural-circuit diversity of the cerebellum. Dev. Growth Differ. 2017, 59, 228–243. [Google Scholar] [CrossRef]
- Ikenaga, T.; Yoshida, M.; Uematsu, K. Cerebellar efferent neurons in teleost fish. Cerebellum 2006, 5, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Rink, E.; Wullimann, M.F. Development of the catecholaminergic system in the early zebrafish brain: An immunohistochemical study. Dev. Brain Res. 2002, 137, 89–100. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-Sequencing Transcriptome and Splicing Database of Glia, Neurons, and Vascular Cells of the Cerebral Cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef] [PubMed]
- Tasic, B.; Menon, V.; Nguyen, T.N.; Kim, T.K.; Jarsky, T.; Yao, Z.; Levi, B.; Gray, L.T.; Sorensen, S.A.; Dolbeare, T.; et al. Adult mouse cortical cell taxonomy revealed by single cell transcriptomics. Nat. Neurosci. 2016, 19, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Hibi, M.; Shimizu, T. Development of the cerebellum and cerebellar neural circuits. Dev. Neurobiol. 2012, 72, 282–301. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T.; Guo, S. The distribution of GAD67-mRNA in the adult zebrafish (teleost) forebrain reveals a prosomeric pattern and suggests previously unidentified homologies to tetrapods. J. Comp. Neurol. 2009, 516, 553–568. [Google Scholar] [CrossRef]
- Kastenhuber, E.; Kratochwil, C.F.; Ryu, S.; Schweitzer, J.; Driever, W. Genetic dissection of dopaminergic and noradrenergic contributions to catecholaminergic tracts in early larval zebrafish. J. Comp. Neurol. 2010, 518, 439–458. [Google Scholar] [CrossRef]
- Filippi, A.; Mahler, J.; Schweitzer, J.; Driever, W. Expression of the paralogous tyrosine hydroxylase encoding genes th1 and th2 reveals the full complement of dopaminergic and noradrenergic neurons in zebrafish larval and juvenile brain. J. Comp. Neurol. 2010, 518, 423–438. [Google Scholar] [CrossRef]
- McLean, D.L.; Fetcho, J.R. Ontogeny and innervation patterns of dopaminergic, noradrenergic, and serotonergic neurons in larval zebrafish. J. Comp. Neurol. 2004, 480, 38–56. [Google Scholar] [CrossRef]
- Higashijima, S.-I.; Mandel, G.; Fetcho, J.R. Distribution of prospective glutamatergic, glycinergic, and GABAergic neurons in embryonic and larval zebrafish. J. Comp. Neurol. 2004, 480, 1–18. [Google Scholar] [CrossRef]
- Robles, E.; Smith, S.J.; Baier, H. Characterization of Genetically Targeted Neuron Types in the Zebrafish Optic Tectum. Front. Neural Circuits 2011, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Partridge, M.; Vincent, A.; Matthews, P.; Puma, J.; Stein, D.; Summerton, J. A Simple Method for Delivering Morpholino Antisense Oligos into the Cytoplasm of Cells. Antisense Nucleic Acid Drug Dev. 1996, 6, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Nasevicius, A.; Ekker, S.C. Effective targeted gene ‘knockdown’ in zebrafish. Nat. Genet. 2000, 26, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Lykke-Andersen, S.; Jensen, T.H. Nonsense-mediated mRNA decay: An intricate machinery that shapes transcriptomes. Nat. Rev. Mol. Cell Biol. 2015, 16, 665–677. [Google Scholar] [CrossRef]
- Ekker, S.C.; Larson, J.D. Morphant technology in model developmental systems. genesis 2001, 30, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Robu, M.E.; Larson, J.D.; Nasevicius, A.; Beiraghi, S.; Brenner, C.; Farber, S.A.; Ekker, S.C. p53 Activation by Knockdown Technologies. PLoS Genet. 2007, 3, 0787–0801. [Google Scholar] [CrossRef] [PubMed]
- Danilova, N.; Kumagai, A.; Lin, J. p53 Upregulation Is a Frequent Response to Deficiency of Cell-Essential Genes. PLoS ONE 2010, 5, e15938. [Google Scholar] [CrossRef] [PubMed]
- Gerety, S.S.; Wilkinson, D.G. Morpholino artifacts provide pitfalls and reveal a novel role for pro-apoptotic genes in hindbrain boundary development. Dev. Biol. 2011, 350, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Stainier, D.Y.R.; Kontarakis, Z.; Rossi, A. Making Sense of Anti-Sense Data. Dev. Cell 2015, 32, 7–8. [Google Scholar] [CrossRef]
- Kwan, K.M.; Fujimoto, E.; Grabher, C.; Mangum, B.D.; Hardy, M.E.; Campbell, D.S.; Parant, J.M.; Yost, H.J.; Kanki, J.P.; Chien, C.-B. The Tol2kit: A multisite gateway-based construction kit forTol2 transposon transgenesis constructs. Dev. Dyn. 2007, 236, 3088–3099. [Google Scholar] [CrossRef]
- Bussmann, J.; Schulte-Merker, S. Rapid BAC selection for tol2-mediated transgenesis in zebrafish. Development 2011, 138, 4327–4332. [Google Scholar] [CrossRef]
- Hans, S.; Kaslin, J.; Freudenreich, D.; Brand, M. Temporally-Controlled Site-Specific Recombination in Zebrafish. PLoS ONE 2009, 4, e4640. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.A.; Giniger, E.; Maniatis, T.; Ptashne, M. GAL4 activates transcription in Drosophila. Nature 1988, 332, 853–856. [Google Scholar] [CrossRef]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 1993, 118, 401–415. [Google Scholar] [PubMed]
- Asakawa, K.; Kawakami, K. Targeted gene expression by the Gal4-UAS system in zebrafish. Dev. Growth Differ. 2008, 50, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, K.; Suster, M.L.; Mizusawa, K.; Nagayoshi, S.; Kotani, T.; Urasaki, A.; Kishimoto, Y.; Hibi, M.; Kawakami, K. Genetic dissection of neural circuits by Tol2 transposon-mediated Gal4 gene and enhancer trapping in zebrafish. Proc. Natl. Acad. Sci. USA 2008, 105, 1255–1260. [Google Scholar] [CrossRef]
- Köster, R.W.; Fraser, S.E. Tracing Transgene Expression in Living Zebrafish Embryos. Dev. Biol. 2001, 233, 329–346. [Google Scholar] [CrossRef] [PubMed]
- Scott, E.K.; Mason, L.; Arrenberg, A.B.; Ziv, L.; Gosse, N.J.; Xiao, T.; Chi, N.C.; Asakawa, K.; Kawakami, K.; Baier, H. Targeting neural circuitry in zebrafish using GAL4 enhancer trapping. Nat. Methods 2007, 4, 323–326. [Google Scholar] [CrossRef] [PubMed]
- Distel, M.; Wullimann, M.F.; Koster, R.W. Optimized Gal4 genetics for permanent gene expression mapping in zebrafish. Proc. Natl. Acad. Sci. USA 2009, 106, 13365–13370. [Google Scholar] [CrossRef] [PubMed]
- Scheer, N.; Campos-Ortega, J.A. Use of the Gal4-UAS technique for targeted gene expression in the zebrafish. Mech. Dev. 1999, 80, 153–158. [Google Scholar] [CrossRef]
- Kimura, Y. alx, a Zebrafish Homolog of Chx10, Marks Ipsilateral Descending Excitatory Interneurons That Participate in the Regulation of Spinal Locomotor Circuits. J. Neurosci. 2006, 26, 5684–5697. [Google Scholar] [CrossRef] [PubMed]
- Kani, S.; Bae, Y.-K.; Shimizu, T.; Tanabe, K.; Satou, C.; Parsons, M.J.; Scott, E.; Higashijima, S.; Hibi, M. Proneural gene-linked neurogenesis in zebrafish cerebellum. Dev. Biol. 2010, 343, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kinkhabwala, A.; Riley, M.; Koyama, M.; Monen, J.; Satou, C.; Kimura, Y.; Higashijima, S.-i.; Fetcho, J. A structural and functional ground plan for neurons in the hindbrain of zebrafish. Proc. Natl. Acad. Sci. USA 2011, 108, 1164–1169. [Google Scholar] [CrossRef] [PubMed]
- Satou, C.; Kimura, Y.; Higashijima, S.-i. Generation of Multiple Classes of V0 Neurons in Zebrafish Spinal Cord: Progenitor Heterogeneity and Temporal Control of Neuronal Diversity. J. Neurosci. 2012, 32, 1771–1783. [Google Scholar] [CrossRef] [PubMed]
- Higashijima, S.-I. Engrailed-1 Expression Marks a Primitive Class of Inhibitory Spinal Interneuron. J. Neurosci. 2004, 24, 5827–5839. [Google Scholar] [CrossRef] [PubMed]
- Stil, A.; Drapeau, P. Neuronal labeling patterns in the spinal cord of adult transgenic Zebrafish: Spinal Neuronal Patterns in the Adult Zebrafish. Dev. Neurobiol. 2016, 76, 642–660. [Google Scholar] [CrossRef] [PubMed]
- Satou, C.; Kimura, Y.; Hirata, H.; Suster, M.L.; Kawakami, K.; Higashijima, S.-I. Transgenic tools to characterize neuronal properties of discrete populations of zebrafish neurons. Development 2013, 140, 3927–3931. [Google Scholar] [CrossRef]
- Bernardos, R.L.; Raymond, P.A. GFAP transgenic zebrafish. Gene Expr. Patterns 2006, 6, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- McLean, D.L.; Fan, J.; Higashijima, S.; Hale, M.E.; Fetcho, J.R. A topographic map of recruitment in spinal cord. Nature 2007, 446, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Takahoko, M.; Okamoto, H. HuC:Kaede, a useful tool to label neural morphologies in networks in vivo. genesis 2006, 44, 136–142. [Google Scholar] [CrossRef]
- Park, H.-C.; Kim, C.-H.; Bae, Y.-K.; Yeo, S.-Y.; Kim, S.-H.; Hong, S.-K.; Shin, J.; Yoo, K.-W.; Hibi, M.; Hirano, T.; et al. Analysis of Upstream Elements in the HuC Promoter Leads to the Establishment of Transgenic Zebrafish with Fluorescent Neurons. Dev. Biol. 2000, 227, 279–293. [Google Scholar] [CrossRef] [PubMed]
- St John, J.A.; Key, B. HuC–eGFP mosaic labelling of neurons in zebrafish enables in vivo live cell imaging of growth cones. J. Mol. Histol. 2012, 43, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, C.; Hu, C.; Wang, Y.; Lin, J.; Jiang, Y.; Li, Q.; Xu, X. CRISPR/Cas9-induced shank3b mutant zebrafish display autism-like behaviors. Mol. Autism 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Higashijima, S.; Hotta, Y.; Okamoto, H. Visualization of Cranial Motor Neurons in Live Transgenic Zebrafish Expressing Green Fluorescent Protein Under the Control of the Islet-1 Promoter/Enhancer. J. Neurosci. 2000, 20, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, K.; Meza Santoscoy, P.; Lygdas, K.; Nikolaeva, Y.; MacDonald, R.; Cunliffe, V.; Nikolaev, A. Imaging Neuronal Activity in the Optic Tectum of Late Stage Larval Zebrafish. J. Dev. Biol. 2018, 6, 6. [Google Scholar] [CrossRef]
- Lam, C.S.; März, M.; Strähle, U. gfap and nestin reporter lines reveal characteristics of neural progenitors in the adult zebrafish brain. Dev. Dyn. 2009, 238, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Park, H.-C.; Topczewska, J.M.; Mawdsley, D.J.; Appel, B. Neural cell fate analysis in zebrafish using olig2 BAC transgenics. Methods Cell Sci. 2003, 25, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Kesavan, G.; Chekuru, A.; Machate, A.; Brand, M. CRISPR/Cas9-Mediated Zebrafish Knock-in as a Novel Strategy to Study Midbrain-Hindbrain Boundary Development. Front. Neuroanat. 2017, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kettleborough, R.N.W.; Busch-Nentwich, E.M.; Harvey, S.A.; Dooley, C.M.; de Bruijn, E.; van Eeden, F.; Sealy, I.; White, R.J.; Herd, C.; Nijman, I.J.; et al. A systematic genome-wide analysis of zebrafish protein-coding gene function. Nature 2013, 496, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Dooley, C.M.; Scahill, C.; Fényes, F.; Kettleborough, R.N.W.; Stemple, D.L.; Busch-Nentwich, E.M. Multi-allelic phenotyping—A systematic approach for the simultaneous analysis of multiple induced mutations. Methods 2013, 62, 197–206. [Google Scholar] [CrossRef]
- Solnica-Krezel, L.; Schier, A.F.; Driever, W. Efficient Recovery of ENU-Induced Mutations from the Zebrafish Germline. Genetics 1994, 136, 1401–1420. [Google Scholar] [PubMed]
- Mullins, M.C.; Hammerschmidt, M.; Haffter, P.; Nüsslein-Volhard, C. Large-scale mutagenesis in the zebrafish: In search of genes controlling development in a vertebrate. Curr. Biol. 1994, 4, 189–202. [Google Scholar] [CrossRef]
- Amsterdam, A.; Burgess, S.; Golling, G.; Chen, W.; Sun, Z.; Townsend, K.; Farrington, S.; Haldi, M.; Hopkins, N. A large-scale insertional mutagenesis screen in zebrafish. Genes Dev. 1999, 13, 2713–2724. [Google Scholar] [CrossRef]
- Simone, B.W.; Martínez-Gálvez, G.; WareJoncas, Z.; Ekker, S.C. Fishing for understanding: Unlocking the zebrafish gene editor’s toolbox. Methods 2018, 150, 3–10. [Google Scholar] [CrossRef]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Ma, Y.; Huang, T.; Chen, Y.; Peng, Y.; Li, B.; Li, J.; Zhang, Y.; Song, B.; Sun, X.; et al. Genetic Modulation of RNA Splicing with a CRISPR-Guided Cytidine Deaminase. Mol. Cell 2018, 72, 380–394.e7. [Google Scholar] [CrossRef]
- Choudhury, S.R.; Cui, Y.; Lubecka, K.; Stefanska, B.; Irudayaraj, J. CRISPR-dCas9 mediated TET1 targeting for selective DNA demethylation at BRCA1 promoter. Oncotarget 2016, 7, 46545–46556. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.S.; Wu, H.; Ji, X.; Stelzer, Y.; Wu, X.; Czauderna, S.; Shu, J.; Dadon, D.; Young, R.A.; Jaenisch, R. Editing DNA Methylation in the Mammalian Genome. Cell 2016, 167, 233–247.e17. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.I.; Celik, H.; Rois, L.E.; Fishberger, G.; Fowler, T.; Rees, R.; Kramer, A.; Martens, A.; Edwards, J.R.; Challen, G.A. Reprogrammable CRISPR/Cas9-based system for inducing site-specific DNA methylation. Biol. Open 2016, 5, 866–874. [Google Scholar] [CrossRef]
- Morita, S.; Noguchi, H.; Horii, T.; Nakabayashi, K.; Kimura, M.; Okamura, K.; Sakai, A.; Nakashima, H.; Hata, K.; Nakashima, K.; et al. Targeted DNA demethylation in vivo using dCas9–peptide repeat and scFv–TET1 catalytic domain fusions. Nat. Biotechnol. 2016, 34, 1060–1065. [Google Scholar] [CrossRef]
- Xu, X.; Tao, Y.; Gao, X.; Zhang, L.; Li, X.; Zou, W.; Ruan, K.; Wang, F.; Xu, G.; Hu, R. A CRISPR-based approach for targeted DNA demethylation. Cell Discov. 2016, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.S.; Lam, I.I.; Clay, H.; Duong, D.N.; Deo, R.C.; Coughlin, S.R. A Rapid Method for Directed Gene Knockout for Screening in G0 Zebrafish. Dev. Cell 2018, 46, 112–125.e4. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Kontarakis, Z.; Gerri, C.; Nolte, H.; Hölper, S.; Krüger, M.; Stainier, D.Y.R. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature 2015, 524, 230–233. [Google Scholar] [CrossRef] [PubMed]
- El-Brolosy, M.A.; Stainier, D.Y.R. Genetic compensation: A phenomenon in search of mechanisms. PLoS Genet. 2017, 13, e1006780. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, V.; Choi, H.M.T.; Fraser, S.E.; Pierce, N.A. Multidimensional quantitative analysis of mRNA expression within intact vertebrate embryos. Development 2018, 145, dev156869. [Google Scholar] [CrossRef] [PubMed]
- Helmchen, F.; Denk, W. Deep tissue two-photon microscopy. Nat. Methods 2005, 2, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Leung, L.C.; Wang, G.X.; Mourrain, P. Imaging zebrafish neural circuitry from whole brain to synapse. Front. Neural Circuits 2013, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Renninger, S.L.; Orger, M.B. Two-photon imaging of neural population activity in zebrafish. Methods 2013, 62, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Mickoleit, M.; Huisken, J. Light sheet microscopy. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 123, pp. 193–215. ISBN 978-0-12-420138-5. [Google Scholar]
- Lemon, W.C.; Keller, P.J. Live imaging of nervous system development and function using light-sheet microscopy. Mol. Reprod. Dev. 2015, 82, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Royer, L.A.; Lemon, W.C.; Chhetri, R.K.; Wan, Y.; Coleman, M.; Myers, E.W.; Keller, P.J. Adaptive light-sheet microscopy for long-term, high-resolution imaging in living organisms. Nat. Biotechnol. 2016, 34, 1267–1278. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, K.; Pratiwi, F.W.; Wu, F.C.M.; Chen, P.; Chen, B.-C. Recent Progress in Light Sheet Microscopy for Biological Applications. Appl. Spectrosc. 2018, 72, 1137–1169. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, A.; Llopis, J.; Heim, R.; McCaffery, J.M.; Adams, J.A.; Ikura, M.; Tsien, R.Y. Fluorescent indicators for Ca2+based on green fluorescent proteins and calmodulin. Nature 1997, 388, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Arrenberg, A.B.; Del Bene, F.; Baier, H. Optical control of zebrafish behavior with halorhodopsin. Proc. Natl. Acad. Sci. USA 2009, 106, 17968–17973. [Google Scholar] [CrossRef]
- Zhu, P.; Narita, Y.; Bundschuh, S.T.; Fajardo, O.; Schärer, Y.-P.Z.; Chattopadhyaya, B.; Bouldoires, E.A.; Stepien, A.E.; Deisseroth, K.; Arber, S.; et al. Optogenetic dissection of neuronal circuits in zebrafish using viral gene transfer and the Tet system. Front. Neural Circuits 2009, 3. [Google Scholar] [CrossRef]
- Reade, A.; Motta-Mena, L.B.; Gardner, K.H.; Stainier, D.Y.; Weiner, O.D.; Woo, S. TAEL: A zebrafish-optimized optogenetic gene expression system with fine spatial and temporal control. Development 2017, 144, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Dal Maschio, M.; Donovan, J.C.; Helmbrecht, T.O.; Baier, H. Linking Neurons to Network Function and Behavior by Two-Photon Holographic Optogenetics and Volumetric Imaging. Neuron 2017, 94, 774–789.e5. [Google Scholar] [CrossRef] [PubMed]
- Saint-Amant, L.; Drapeau, P. Time course of the development of motor behaviors in the zebrafish embryo. J. Neurobiol. 1998, 37, 622–632. [Google Scholar] [CrossRef]
- Saint-Amant, L.; Drapeau, P. Synchronization of an Embryonic Network of Identified Spinal Interneurons Solely by Electrical Coupling. Neuron 2001, 31, 1035–1046. [Google Scholar] [CrossRef]
- Legradi, J.; el Abdellaoui, N.; van Pomeren, M.; Legler, J. Comparability of behavioural assays using zebrafish larvae to assess neurotoxicity. Environ. Sci. Pollut. Res. 2015, 22, 16277–16289. [Google Scholar] [CrossRef] [PubMed]
- Tzschentke, T.M. Measuring reward with the conditioned place preference paradigm: A comprehensive review of drug effects, recent progress and new issues. Prog. Neurobiol. 1998, 56, 613–672. [Google Scholar] [CrossRef]
- Williams, F.E.; White, D.; Messer, W.S. A simple spatial alternation task for assessing memory function in zebrafish. Behav. Process. 2002, 58, 125–132. [Google Scholar] [CrossRef]
- Blaser, R.E.; Vira, D.G. Experiments on learning in zebrafish (Danio rerio): A promising model of neurocognitive function. Neurosci. Biobehav. Rev. 2014, 42, 224–231. [Google Scholar] [CrossRef]
- Wong, K.; Elegante, M.; Bartels, B.; Elkhayat, S.; Tien, D.; Roy, S.; Goodspeed, J.; Suciu, C.; Tan, J.; Grimes, C.; et al. Analyzing habituation responses to novelty in zebrafish (Danio rerio). Behav. Brain Res. 2010, 208, 450–457. [Google Scholar] [CrossRef]
- Wang, Y.; Zhong, H.; Wang, C.; Gao, D.; Zhou, Y.; Zuo, Z. Maternal exposure to the water soluble fraction of crude oil, lead and their mixture induces autism-like behavioral deficits in zebrafish (Danio rerio) larvae. Ecotoxicol. Environ. Saf. 2016, 134, 23–30. [Google Scholar] [CrossRef]
- Miller, N.; Gerlai, R. From Schooling to Shoaling: Patterns of Collective Motion in Zebrafish (Danio rerio). PLoS ONE 2012, 7, e48865. [Google Scholar] [CrossRef]
- Cachat, J.; Kyzar, E.J.; Collins, C.; Gaikwad, S.; Green, J.; Roth, A.; El-Ounsi, M.; Davis, A.; Pham, M.; Landsman, S.; et al. Unique and potent effects of acute ibogaine on zebrafish: The developing utility of novel aquatic models for hallucinogenic drug research. Behav. Brain Res. 2013, 236, 258–269. [Google Scholar] [CrossRef]
- Ariyasiri, K.; Choi, T.-I.; Kim, O.-H.; Hong, T.I.; Gerlai, R.; Kim, C.-H. Pharmacological (ethanol) and mutation (sam2 KO) induced impairment of novelty preference in zebrafish quantified using a new three-chamber social choice task. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 88, 53–65. [Google Scholar] [CrossRef]
- Kim, L.; He, L.; Maaswinkel, H.; Zhu, L.; Sirotkin, H.; Weng, W. Anxiety, hyperactivity and stereotypy in a zebrafish model of fragile X syndrome and autism spectrum disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 55, 40–49. [Google Scholar] [CrossRef]
- Maaswinkel, H.; Zhu, L.; Weng, W. Assessing Social Engagement in Heterogeneous Groups of Zebrafish: A New Paradigm for Autism-Like Behavioral Responses. PLoS ONE 2013, 8, e75955. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a Comprehensive Catalog of Zebrafish Behavior 1.0 and Beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Van Meer, P.; Raber, J. Mouse behavioural analysis in systems biology. Biochem. J. 2005, 389, 593–610. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Stewart, A.M.; Gerlai, R. Zebrafish as an emerging model for studying complex brain disorders. Trends Pharmacol. Sci. 2014, 35, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Shiotsuki, H.; Yoshimi, K.; Shimo, Y.; Funayama, M.; Takamatsu, Y.; Ikeda, K.; Takahashi, R.; Kitazawa, S.; Hattori, N. A rotarod test for evaluation of motor skill learning. J. Neurosci. Methods 2010, 189, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Tierney, K.B. Swimming Performance Assessment in Fishes. J. Vis. Exp. 2011, 51, e2572. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.J.H.; Zerulla, T.C.; Tierney, K.B. Zebrafish (Danio rerio) as a model for the study of aging and exercise: Physical ability and trainability decrease with age. Exp. Gerontol. 2014, 50, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Fromme, H.; Tittlemier, S.A.; Völkel, W.; Wilhelm, M.; Twardella, D. Perfluorinated compounds—Exposure assessment for the general population in western countries. Int. J. Hyg. Environ. Health 2009, 212, 239–270. [Google Scholar] [CrossRef]
- Hoffman, K.; Webster, T.F.; Weisskopf, M.G.; Weinberg, J.; Vieira, V.M. Exposure to Polyfluoroalkyl Chemicals and Attention Deficit/Hyperactivity Disorder in U.S. Children 12–15 Years of Age. Environ. Health Perspect. 2010, 118, 1762–1767. [Google Scholar] [CrossRef]
- Gump, B.B.; Wu, Q.; Dumas, A.K.; Kannan, K. Perfluorochemical (PFC) Exposure in Children: Associations with Impaired Response Inhibition. Environ. Sci. Technol. 2011, 45, 8151–8159. [Google Scholar] [CrossRef]
- Spulber, S.; Kilian, P.; Wan Ibrahim, W.N.; Onishchenko, N.; Ulhaq, M.; Norrgren, L.; Negri, S.; Di Tuccio, M.; Ceccatelli, S. PFOS Induces Behavioral Alterations, Including Spontaneous Hyperactivity That Is Corrected by Dexamfetamine in Zebrafish Larvae. PLoS ONE 2014, 9, e94227. [Google Scholar] [CrossRef] [PubMed]
- Ornoy, A. Valproic acid in pregnancy: How much are we endangering the embryo and fetus? Reprod. Toxicol. 2009, 28, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Beunders, G.; Voorhoeve, E.; Golzio, C.; Pardo, L.M.; Rosenfeld, J.A.; Talkowski, M.E.; Simonic, I.; Lionel, A.C.; Vergult, S.; Pyatt, R.E.; et al. Exonic Deletions in AUTS2 Cause a Syndromic Form of Intellectual Disability and Suggest a Critical Role for the C Terminus. Am. J. Hum. Genet. 2013, 92, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Oksenberg, N.; Stevison, L.; Wall, J.D.; Ahituv, N. Function and Regulation of AUTS2, a Gene Implicated in Autism and Human Evolution. PLoS Genet. 2013, 9, e1003221. [Google Scholar] [CrossRef] [PubMed]
- Van Bon, B.; Hoischen, A.; Hehir-Kwa, J.; de Brouwer, A.; Ruivenkamp, C.; Gijsbers, A.; Marcelis, C.; de Leeuw, N.; Veltman, J.; Brunner, H.; et al. Intragenic deletion in DYRK1A leads to mental retardation and primary microcephaly. Clin. Genet. 2011, 79, 296–299. [Google Scholar] [CrossRef] [PubMed]
- O’Roak, B.J.; Vives, L.; Fu, W.; Egertson, J.D.; Stanaway, I.B.; Phelps, I.G.; Carvill, G.; Kumar, A.; Lee, C.; Ankenman, K.; et al. Multiplex Targeted Sequencing Identifies Recurrently Mutated Genes in Autism Spectrum Disorders. Science 2012, 338, 1619–1622. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.-H.; Cho, H.-J.; Han, E.; Hong, T.I.; Ariyasiri, K.; Choi, J.-H.; Hwang, K.-S.; Jeong, Y.-M.; Yang, S.-Y.; Yu, K.; et al. Zebrafish knockout of Down syndrome gene, DYRK1A, shows social impairments relevant to autism. Mol. Autism 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Bernier, R.; Golzio, C.; Xiong, B.; Stessman, H.A.; Coe, B.P.; Penn, O.; Witherspoon, K.; Gerdts, J.; Baker, C.; Vulto-van Silfhout, A.T.; et al. Disruptive CHD8 Mutations Define a Subtype of Autism Early in Development. Cell 2014, 158, 263–276. [Google Scholar] [CrossRef]
- Sugathan, A.; Biagioli, M.; Golzio, C.; Erdin, S.; Blumenthal, I.; Manavalan, P.; Ragavendran, A.; Brand, H.; Lucente, D.; Miles, J.; et al. CHD8 regulates neurodevelopmental pathways associated with autism spectrum disorder in neural progenitors. Proc. Natl. Acad. Sci. USA 2014, 111, E4468–E4477. [Google Scholar] [CrossRef] [PubMed]
- Marin-Valencia, I.; Novarino, G.; Johansen, A.; Rosti, B.; Issa, M.Y.; Musaev, D.; Bhat, G.; Scott, E.; Silhavy, J.L.; Stanley, V.; et al. A homozygous founder mutation in TRAPPC6B associates with a neurodevelopmental disorder characterised by microcephaly, epilepsy and autistic features. J. Med. Genet. 2018, 55, 48–54. [Google Scholar] [CrossRef]
- Wu, Y.-J.; Hsu, M.-T.; Ng, M.-C.; Amstislavskaya, T.G.; Tikhonova, M.A.; Yang, Y.-L.; Lu, K.-T.; Fragile, X. Mental Retardation-1 Knockout Zebrafish Shows Precocious Development in Social Behavior. Zebrafish 2017, 14, 438–443. [Google Scholar] [CrossRef]
- Hoffman, E.J.; Turner, K.J.; Fernandez, J.M.; Cifuentes, D.; Ghosh, M.; Ijaz, S.; Jain, R.A.; Kubo, F.; Bill, B.R.; Baier, H.; et al. Estrogens Suppress a Behavioral Phenotype in Zebrafish Mutants of the Autism Risk Gene, CNTNAP2. Neuron 2016, 89, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.C.; Voelker, L.H.; Shah, A.N.; Moens, C.B. Neurobeachin Is Required Postsynaptically for Electrical and Chemical Synapse Formation. Curr. Biol. 2015, 25, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Golzio, C.; Willer, J.; Talkowski, M.E.; Oh, E.C.; Taniguchi, Y.; Jacquemont, S.; Reymond, A.; Sun, M.; Sawa, A.; Gusella, J.F.; et al. KCTD13 is a major driver of mirrored neuroanatomical phenotypes of the 16p11.2 copy number variant. Nature 2012, 485, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Blaker-Lee, A.; Gupta, S.; McCammon, J.M.; De Rienzo, G.; Sive, H. Zebrafish homologs of genes within 16p11.2, a genomic region associated with brain disorders, are active during brain development, and include two deletion dosage sensor genes. Dis. Model. Mech. 2012, 5, 834–851. [Google Scholar] [CrossRef]
- Turner, T.N.; Sharma, K.; Oh, E.C.; Liu, Y.P.; Collins, R.L.; Sosa, M.X.; Auer, D.R.; Brand, H.; Sanders, S.J.; Moreno-De-Luca, D.; et al. Loss of δ-catenin function in severe autism. Nature 2015, 520, 51–56. [Google Scholar] [CrossRef]
- Hofmeister, W.; Nilsson, D.; Topa, A.; Anderlid, B.-M.; Darki, F.; Matsson, H.; Tapia Páez, I.; Klingberg, T.; Samuelsson, L.; Wirta, V.; et al. CTNND2—A candidate gene for reading problems and mild intellectual disability. J. Med. Genet. 2015, 52, 111–122. [Google Scholar] [CrossRef]
- Heon, E.; Kim, G.; Qin, S.; Garrison, J.E.; Tavares, E.; Vincent, A.; Nuangchamnong, N.; Scott, C.A.; Slusarski, D.C.; Sheffield, V.C. Mutations in C8ORF37 cause Bardet Biedl syndrome (BBS21). Hum. Mol. Genet. 2016, 25, 2283–2294. [Google Scholar] [CrossRef] [PubMed]
- Tucker, B.; Richards, R.I.; Lardelli, M. Contribution of mGluR and Fmr1 functional pathways to neurite morphogenesis, craniofacial development and fragile X syndrome. Hum. Mol. Genet. 2006, 15, 3446–3458. [Google Scholar] [CrossRef]
- Ng, M.-C.; Yang, Y.-L.; Lu, K.-T. Behavioral and Synaptic Circuit Features in a Zebrafish Model of Fragile X Syndrome. PLoS ONE 2013, 8, e51456. [Google Scholar] [CrossRef]
- Lange, M.; Norton, W.; Coolen, M.; Chaminade, M.; Merker, S.; Proft, F.; Schmitt, A.; Vernier, P.; Lesch, K.-P.; Bally-Cuif, L. The ADHD-susceptibility gene lphn3.1 modulates dopaminergic neuron formation and locomotor activity during zebrafish development. Mol. Psychiatry 2012, 17, 946–954. [Google Scholar] [CrossRef]
- Pietri, T.; Roman, A.-C.; Guyon, N.; Romano, S.A.; Washbourne, P.; Moens, C.B.; de Polavieja, G.G.; Sumbre, G. The first mecp2-null zebrafish model shows altered motor behaviors. Front. Neural Circuits 2013, 7, 1–10. [Google Scholar] [CrossRef]
- Nozawa, K.; Lin, Y.; Kubodera, R.; Shimizu, Y.; Tanaka, H.; Ohshima, T. Zebrafish Mecp2 is required for proper axonal elongation of motor neurons and synapse formation: Zebrafish Mecp2. Dev. Neurobiol. 2017, 77, 1101–1113. [Google Scholar] [CrossRef]
- Elsen, G.E.; Choi, L.Y.; Prince, V.E.; Ho, R.K. The autism susceptibility gene met regulates zebrafish cerebellar development and facial motor neuron migration. Dev. Biol. 2009, 335, 78–92. [Google Scholar] [CrossRef]
- Gauthier, J.; Champagne, N.; Lafrenière, R.G.; Xiong, L.; Spiegelman, D.; Brustein, E.; Lapointe, M.; Peng, H.; Côté, M.; Noreau, A.; et al. De novo mutations in the gene encoding the synaptic scaffolding protein SHANK3 in patients ascertained for schizophrenia. Proc. Natl. Acad. Sci. USA 2010, 107, 7863–7868. [Google Scholar] [CrossRef]
- Kozol, R.A.; Cukier, H.N.; Zou, B.; Mayo, V.; De Rubeis, S.; Cai, G.; Griswold, A.J.; Whitehead, P.L.; Haines, J.L.; Gilbert, J.R.; et al. Two knockdown models of the autism genes SYNGAP1 and SHANK3 in zebrafish produce similar behavioral phenotypes associated with embryonic disruptions of brain morphogenesis. Hum. Mol. Genet. 2015, 24, 4006–4023. [Google Scholar] [CrossRef]
- Kokel, D.; Bryan, J.; Laggner, C.; White, R.; Cheung, C.Y.J.; Mateus, R.; Healey, D.; Kim, S.; Werdich, A.A.; Haggarty, S.J.; et al. Rapid behavior-based identification of neuroactive small molecules in the zebrafish. Nat. Chem. Biol. 2010, 6, 231–237. [Google Scholar] [CrossRef]
- Rihel, J.; Prober, D.A.; Arvanites, A.; Lam, K.; Zimmerman, S.; Jang, S.; Haggarty, S.J.; Kokel, D.; Rubin, L.L.; Peterson, R.T.; et al. Zebrafish Behavioral Profiling Links Drugs to Biological Targets and Rest/Wake Regulation. Science 2010, 327, 348–351. [Google Scholar] [CrossRef]
- Ellis, L.D.; Soanes, K.H. A larval zebrafish model of bipolar disorder as a screening platform for neuro-therapeutics. Behav. Brain Res. 2012, 233, 450–457. [Google Scholar] [CrossRef]
- Zimmermann, F.F.; Gaspary, K.V.; Siebel, A.M.; Bonan, C.D. Oxytocin reversed MK-801-induced social interaction and aggression deficits in zebrafish. Behav. Brain Res. 2016, 311, 368–374. [Google Scholar] [CrossRef]
- Li, F.; Lin, J.; Liu, X.; Li, W.; Ding, Y.; Zhang, Y.; Zhou, S.; Guo, N.; Li, Q. Characterization of the locomotor activities of zebrafish larvae under the influence of various neuroactive drugs. Ann. Transl. Med. 2018, 6, 173. [Google Scholar] [CrossRef]
- Schaafsma, S.M.; Pfaff, D.W. Etiologies underlying sex differences in Autism Spectrum Disorders. Front. Neuroendocrinol. 2014, 35, 255–271. [Google Scholar] [CrossRef]
- Jacquemont, S.; Coe, B.P.; Hersch, M.; Duyzend, M.H.; Krumm, N.; Bergmann, S.; Beckmann, J.S.; Rosenfeld, J.A.; Eichler, E.E. A Higher Mutational Burden in Females Supports a “Female Protective Model” in Neurodevelopmental Disorders. Am. J. Hum. Genet. 2014, 94, 415–425. [Google Scholar] [CrossRef]
- McEwen, B.S.; Milner, T.A. Understanding the broad influence of sex hormones and sex differences in the brain: Sex Hormones Affect the Whole Brain. J. Neurosci. Res. 2017, 95, 24–39. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Line | Promoter | Features | Cells Tagged | References |
|---|---|---|---|---|
| Tg(alx:Kaede) | alx (human CHX10) | Kaede converts from green to red after UV radiation; cell tracking | In spinal cord: ipsilateral descending neurons, glutamatergic (V2a neurons) | [71] |
| Tg(alx:GFP) | GFP expression | |||
| Tg(atoh1a:EGFP) | atonal bHLH transcription factor 1a | GFP expression | Cerebellum (glutamatergic neurons) and tegmentum cells (cells connecting to the hypothalamus and optic tectum) | [72] |
| Tg(barhl2:GFP) | barh-like homeobox 2 promoter | GFP expression | Neurons at the dorso-lateral edge of the hindbrain and spinal cord | [73] |
| Tg(dbx1b:GFP) | developing brain homeobox 1b promoter | GFP expression | Early neuronal marker, expressed in a population of glutamatergic and glycinergic neurons in the hindbrain | [73,74] |
| Tg(dbx1b:loxP-DsRed-loxP-GFP) | Conditional system, depending on cre expression | |||
| Tg(dbx1b:Cre) | cre recombinase expression | |||
| Tg(eng1b:eng-GFP); Tg(hsp70:eng-GFP) | engrailed 1 | Expression of fusion eng-GFP protein, drived by endogenous (eng1b) or heat shock (hsp70) promoter | In spinal cord: ipsilateral gylcinergic interneurons (circumferencial ascending interneurons), V1 neurons | [75] |
| Tg(evx1:GFP) | even-skipped homeobox 1 | GFP expression | In spinal cord: commissural interneurons, ventral projections from the spinal cord | [76] |
| Tg(gad1b:RFP) | glutamic acid decarboxylase | RFP expression | GABAergic neurons | [74,77] |
| Tg(gad1b:loxP-DsRed-loxP-GFP) | Conditional system, depending on cre expression | |||
| Tg (gfap:GFP) | glial fibrillary acidic protein | GFP expression | Neural stem cells throughout the brain, retina, hindbrain, and spinal cord | [74,78] |
| Tg (gfap:dTomato) | dTomato expression | |||
| Tg(glyt2:GFP); Tg(glyt2:RFP) | glycine transporter-2 promoter | GFP or RFP expression | Glycinergic neurons | [74,77,79] |
| Tg(glyt2:loxP-DsRed-loxP-GFP) | Conditional system, depending on cre expression | |||
| Tg(vglut2a:GFP) | glutamate transporter promoter | GFP expression | Glutamatergic neurons | [37,74] |
| Tg(vglut2a:loxP-DsRed-loxP-GFP) | Conditional system, depending on cre expression | |||
| Tg(hb9:GFP) | motor neuron and pancreas homeobox 1 (mnx1), previously known as hb9 | GFP expression | In spinal cord: motor neurons and interneurons | [76] |
| Tg(HuC:CAM) | HuC (or elav3, ELAV like neuron-specific RNA binding protein 3) | cameleon expression (calcium indicator derivative from GFP) | Early neuronal marker; spinal cord: glycinergic interneurons | [74,75,80,81,82,83] |
| Tg(HuC:Ckaede) | Kaede converts from green to red after radiation with UV; cell tracking | |||
| Tg(HuC:GFP) Tg(HuC:RFP) | GFP or RFP expression | |||
| Tg(HuC:loxP-DsRed-loxP-GFP) | Conditional system, depending on cre expression | |||
| Tg(hox9a:Cre) | homeobox 9 | cre recombinase expression | Spinal cord neurons | [74] |
| Tg(isl1:GFP) | LIM/homeobox 1 | GFP expression | Ubiquitous motor neurons | [76,84] |
| Tg(lhx2a:GFP) | LIM Homeobox 2a | GFP expression | Olfactory bulb neurons | [14] |
| Tg(lhx2a:gap:YFP) | gap: membrane signal, YFP will be found in the cell membrane | |||
| Tg(NBT:GCaMP3) | neural beta-tubulin | GCaMP3: calcium reporter, allows live imaging of neuronal activity | Ubiquitous neuronal cells | [85] |
| Tg(nestin:GFP) | nestin | GFP expression | Ubiquitous neural stem cells | [86] |
| Tg(ngn1:GFP); Tg(neurog1:RFP) | neurogenin1 | GFP or RFP expression | In spinal cord: Rohon Beard and dorsal root ganglia neurons | [76,77] |
| Tg(olig2:EGFP) | oligodendrocyte lineage transcription factor 2 | GFP expression | Oligodendrocytes, spinal cord motor neurons, and cerebellum cells | [37,87] |
| Tg(ptf1a:EGFP) | pancreas associated transcription factor 1a | GFP expression | Purkinje cells and telencephalon’s ventricular zone | [72] |
| Tg(tub:CAM) | golfish neural tubulin | CAM: cameleon expression (calcium indicator derivative from GFP) | In spinal cord: ipsilateral glycinergic interneurons (circumferential ascending interneurons) | [75] |
| GFP/tRFP-ki | otx2 | knock-in of fluorescent tag | Retina, midbrain, MHB expression | [88] |
| venus/tRFP-ki | pax2a | knock-in of fluorescent tag | MHB and otic vesicle expression |
| Gene | Human Disorder | Technique | Observations | Reference |
|---|---|---|---|---|
| 16p11.2 CNV; kctd13 | ASD, ID | Splice block and translation MO, RNA overexpression | Overexpression of kctd13: microcephaly caused by decreased proliferation and increased apoptosis; kockdown of kctd13: macrocephaly caused by increased proliferation; knockdown several genes in 16p11.2 region results in neural tube and axon morphogenesis defects | [155,156] |
| auts2 (autism susceptibility candidate 2) | ASD, ADHD, DD, epilepsy | Translation and splice block MO | Smaller body size; severe decrease in the number of neurons in the brain, retina, and spinal cord. Touch evoked response assay showed decreased response from MO-injected larvae | [145] |
| chd8 (chromodomain helicase DNA binding protein 8) | ASD, DD | Splice block MO | Increased head size, with increased number of midbrain and forebrain progenitors | [149,150] |
| cntnap2 (contactin associated protein-like 2) | ASD | Zinc fingers | Decrease in GABAergic cells in the pallium and cerebellum, increased seizures when exposed to PTZ; increased nighttime activity | [153] |
| ctnnd2 (delta catenin) | ASD, ID | Splice block MO | Gastrulation defects; co-injection with wt or mutant ctnnd2 fully (wt) or partially (mutant) rescues phenotype; abnormal neuronal patterning, with ectopic Isl1-positive cells in the optic recess region | [157,158] |
| c8orf37 (chromosome 8 open reading frame 37) | Bardet Biedl syndrome, ASD | Translation and splice block MO | Abnormal response to visual motor response test; co-injection with wt mRNA rescues phenotype, co-injection with mutant mRNA not able to rescue the phenotype | [159] |
| dyrk1a (dual-specificity tyrosine-(Y)-phosphorylation regulated kinase 1A) | Down syndrome, ASD, ID | TALENs | Adult mutants present with microcephaly, possibly due to cell death, and reduced neuronal activation (c-fos expression) in the hypothalamus, and reduced expression of chr (corticotrophin-releasing hormone) in the preoptic region; novel tank test revealed shorter freezing time and increased exploration; social interaction test showed mutant specimen spending less time close to interaction fish | [148] |
| fmr (fragil x mental retardation) | Fragile X syndrome, learningand cognitive deficits, ADHD, ASD | Translation block MO, ENU mutagenesis | MO injection: patterning defects of the forebrain and MHB; hindbrain oedema; spinal cord neurites with increased branching; co-injection with fmr1 mRNA or MPEP rescues the phenotypes. Knock-out studies: embryos show no phenotypes (possible compensation mechanism?); open field test on adult zebrafish showed decreased freezing | [131,152,160,161] |
| glra2 (glycine receptor alpha 2) | ASD | Translation block MO | Hyperbranching of spinal motor neuron axons; co-injection with wt GLRA2 rescues phenotype, while co-injection with mutant RNA fails to rescue | [6] |
| lphn3.1 (latrophilin 3) | ADHD | Splice block MO | MO-injected embryos swim longer distances in an un-evoked swimming assay; neuron quantification showed a reduction in the number and a general disorganization of the dopaminergic neurons; behavioral phenotype can be rescued by exposure to ADHD treatment drugs methylphenidate and atomoxetine | [162] |
| mecp2 (methyl-CpG-binding protein 2) | ASD, Rett syndrome | Translation and splice block MOs; ENU mutagenesis | MO studies: abnormal neuronal branching and growth of motor neurons; touch-evoked response assay showed a slower response. Mutant studies: touch-evoked response assay showed that mutant larvae have a prolonged coiling response and swimming behavior at later stages shows a reduced in spontaneous activity | [163,164] |
| met (tyrosine kinase receptor) | ASD | Translation block MO | Reduction of the cerebellum size by reduced proliferation, impairs migration of hindbrain facial motor neurons | [165] |
| nbea (neurobeachin) | ASD | ENU mutagenesis | Loss of GlyR in both hindbrain and spinal cord, reduced dendritic complexity, and defects in glycinergic synaptogenesis; startle response test shows that mutant larvae respond in only 50% of the trials | [154] |
| shank3b (sh3 and multiple ankyrin repeat protein 3) | ASD and ID | Splice block MO; CRISPR/Cas9 | MO studies: affected brain patterning; reduction of GABAergic and glutamatergic neurons in the mid and hindbrain; touch-evoked response is impaired. Stable mutant studies: swimming behavior and visual motor response test showed a decrease in response in larvae and adults; embryonic brain shows a decrease in the huc-positive cells, this difference becomes smaller throughout development | [83,166,167] |
| syngap1 (synaptic ras GTPase activating protein) | ASD and ID | Splice block MO | Brain patterning affected; reduction of GABAergic and glutamatergic neurons in the mid and hindbrain; touch-evoked response is impaired in morphant larvae, with reduced swim speed | [167] |
| trappc6b (trafficking protein particle complex 6b) | Microcephaly, epilepsy, ASD | Translation and splice block MO | Decreased head size due to increased apoptosis, increased spontaneous neuron firing and activity | [151] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaz, R.; Hofmeister, W.; Lindstrand, A. Zebrafish Models of Neurodevelopmental Disorders: Limitations and Benefits of Current Tools and Techniques. Int. J. Mol. Sci. 2019, 20, 1296. https://doi.org/10.3390/ijms20061296
Vaz R, Hofmeister W, Lindstrand A. Zebrafish Models of Neurodevelopmental Disorders: Limitations and Benefits of Current Tools and Techniques. International Journal of Molecular Sciences. 2019; 20(6):1296. https://doi.org/10.3390/ijms20061296
Chicago/Turabian StyleVaz, Raquel, Wolfgang Hofmeister, and Anna Lindstrand. 2019. "Zebrafish Models of Neurodevelopmental Disorders: Limitations and Benefits of Current Tools and Techniques" International Journal of Molecular Sciences 20, no. 6: 1296. https://doi.org/10.3390/ijms20061296
APA StyleVaz, R., Hofmeister, W., & Lindstrand, A. (2019). Zebrafish Models of Neurodevelopmental Disorders: Limitations and Benefits of Current Tools and Techniques. International Journal of Molecular Sciences, 20(6), 1296. https://doi.org/10.3390/ijms20061296