Metalloproteinases TACE and MMP-9 Differentially Regulate Death Factors on Adult and Neonatal Monocytes After Infection with Escherichia coli

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
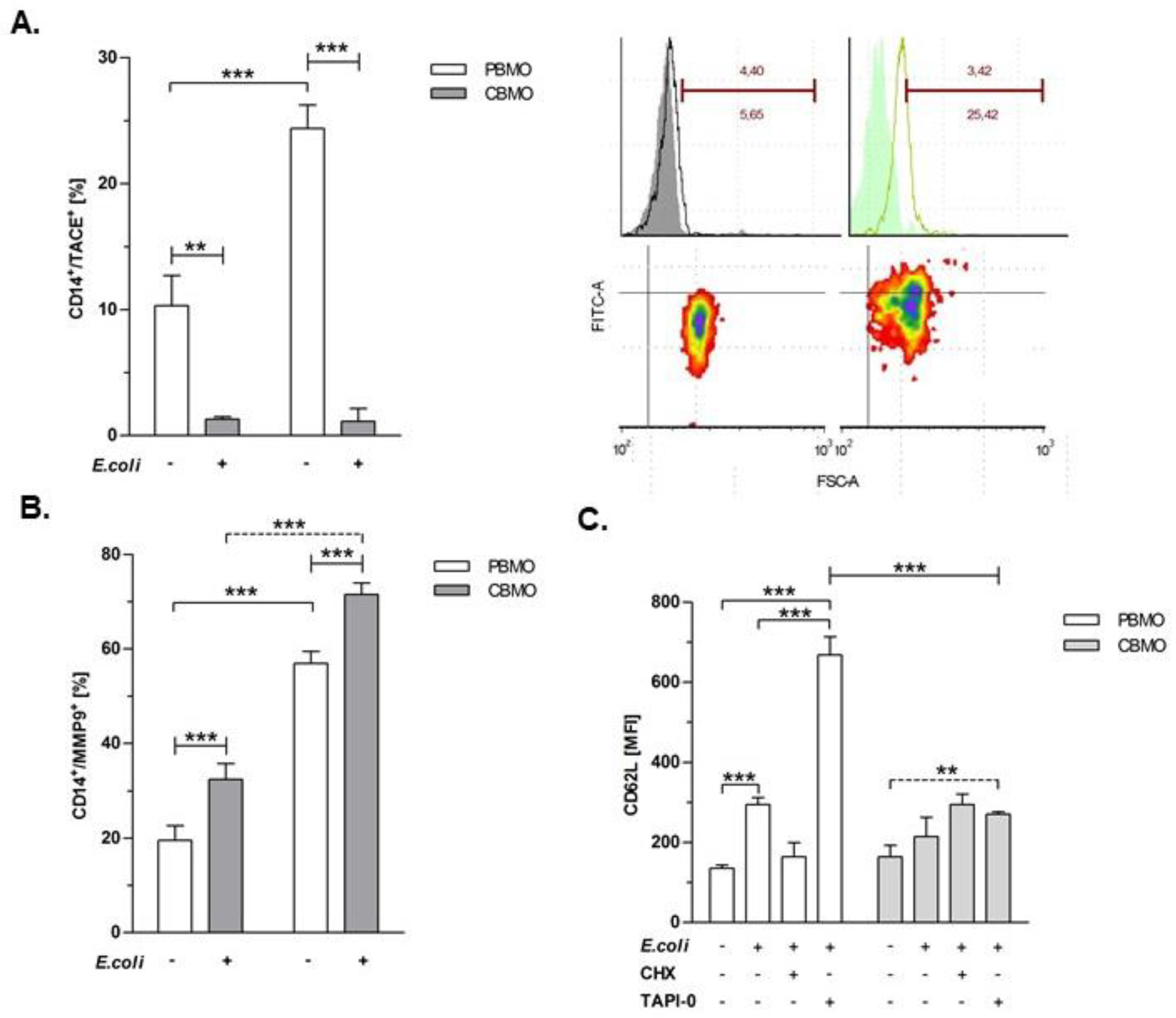
2.1. Plasma-Membrane Expression of Metalloproteinases is Induced by E. coli Infection
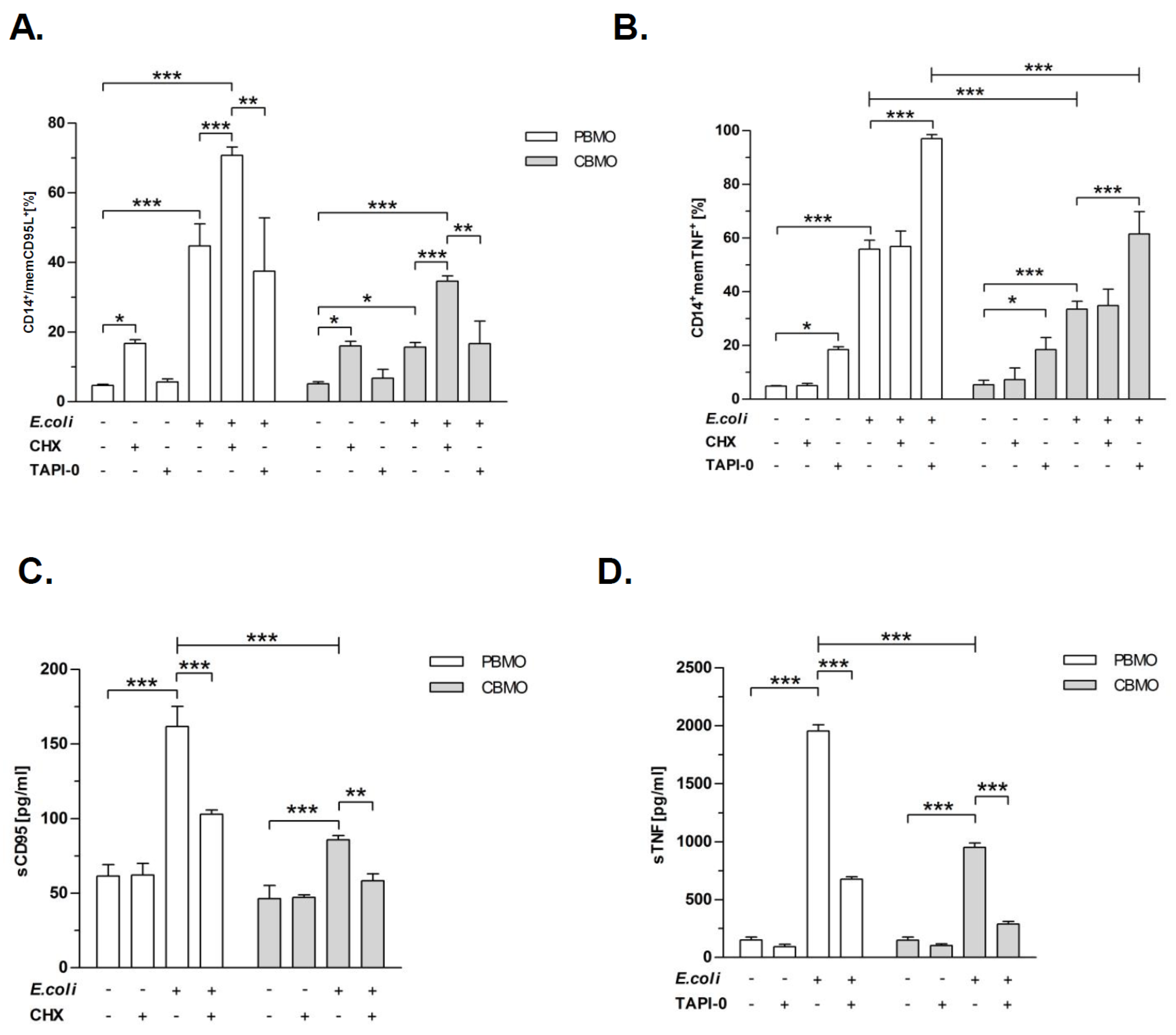
2.2. Metalloproteinase-Inhibition Down-Regulates the Expression of CD95L and TNF
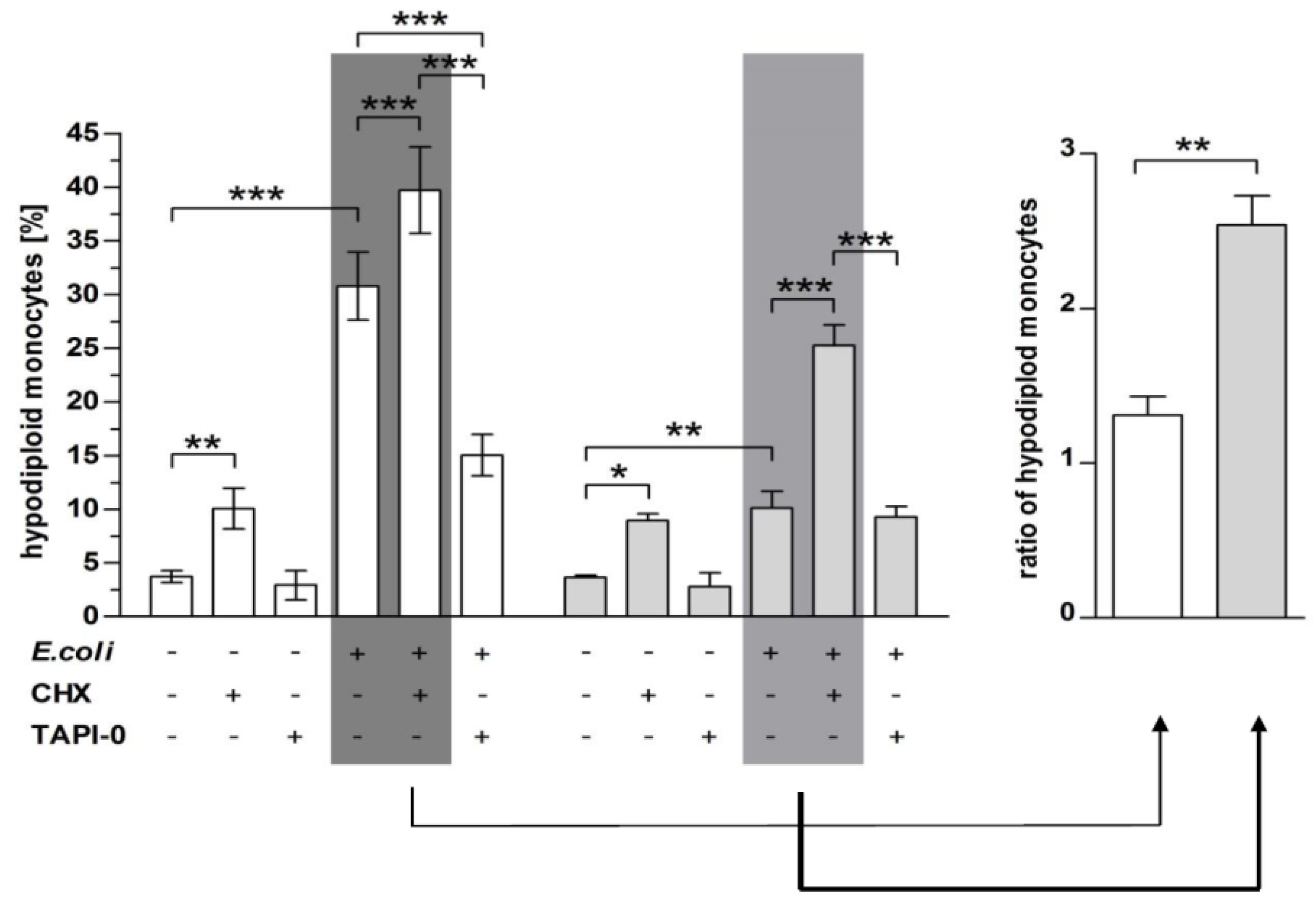
2.3. Inhibition of TACE Reduced PICD, but Inhibition of MMP-9 Restored PICD in CBMO
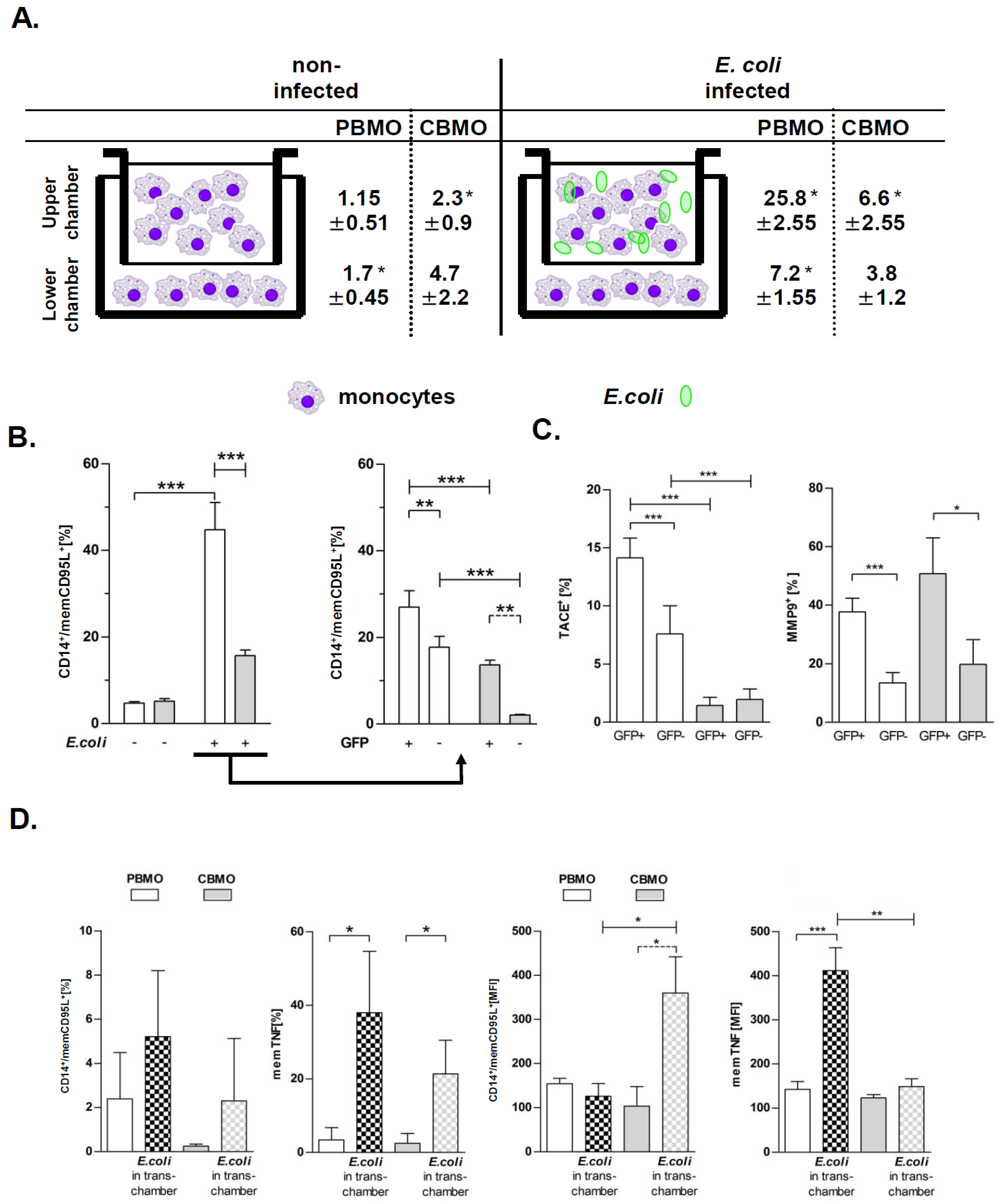
2.4. TNF and E. coli Infection Triggers Expression of memCD95L
2.5. Apoptosis of Bystander Monocytes is Reduced Without Cellular Contact
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Bacteria
E. coli-GFP
4.3. Reagents
4.4. Mononuclear Cell Cultures
4.5. Flow Cytometry
4.6. Detection of Hypodiploid Nuclei
4.7. Transwell Experiments
4.8. ELISA
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
References
- Cockle, J.V.; Gopichandran, N.; Walker, J.J.; Levene, M.I.; Orsi, N.M. Matrix metalloproteinases and their tissue inhibitors in preterm perinatal complications. Reprod. Sci. 2007, 14, 629–645. [Google Scholar] [CrossRef] [PubMed]
- Gille, C.; Steffen, F.; Lauber, K.; Keppeler, H.; Leiber, A.; Spring, B.; Poets, C.F.; Orlikowsky, T.W. Clearance of apoptotic neutrophils is diminished in cord blood monocytes and does not lead to reduced IL-8 production. Pediatr. Res. 2009, 66, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Dammann, O.; Leviton, A. Intermittent or sustained systemic inflammation and the preterm brain. Pediatr. Res. 2014, 75, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Gille, C.; Leiber, A.; Spring, B.; Kempf, V.A.; Loeffler, J.; Poets, C.F.; Orlikowsky, T.W. Diminished phagocytosis-induced cell death (PICD) in neonatal monocytes upon infection with Escherichia coli. Pediatr. Res. 2008, 63, 33–38. [Google Scholar] [CrossRef]
- Gille, C.; Dreschers, S.; Leiber, A.; Lepiorz, F.; Krusch, M.; Grosse-Opphoff, J.; Spring, B.; Haas, M.; Urschitz, M.; Poets, C.F. The CD95/CD95L pathway is involved in phagocytosis-induced cell death of monocytes and may account for sustained inflammation in neonates. Pediatr. Res. 2013, 73, 402–408. [Google Scholar] [CrossRef]
- Dreschers, S.; Gille, C.; Haas, M.; Grosse-Ophoff, J.; Schneider, M.; Leiber, A.; Bühring, H.-J.; Orlikowsky, T.W. Infection-induced bystander-apoptosis of monocytes is TNF-alpha-mediated. PLoS ONE 2013, 8, e53589. [Google Scholar] [CrossRef]
- Gille, C.; Leiber, A.; Mundle, I.; Spring, B.; Abele, H.; Spellerberg, B.; Hartmann, H.; Poets, C.F.; Orlikowsky, T.W. Phagocytosis and postphagocytic reaction of cord blood and adult blood monocyte after infection with green fluorescent protein-labeled Escherichia coli and group B Streptococci. Cytom. B Clin. Cytom. 2009, 76, 271–284. [Google Scholar] [CrossRef]
- Dreschers, S.; Gille, C.; Haas, M.; Seubert, F.; Platen, C.; Orlikowsky, T.W. Reduced internalization of TNF-a/TNFR1 down-regulates caspase dependent phagocytosis induced cell death (PICD) in neonatal monocytes. PLoS ONE 2017, 12, e0182415. [Google Scholar] [CrossRef]
- Vandooren, J.; Van den Steen, P.E.; Opdenakker, G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9): The next decade. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 222–272. [Google Scholar] [CrossRef]
- Levine, S.J. Molecular mechanisms of soluble cytokine receptor generation. J. Biol. Chem. 2008, 283, 14177–14181. [Google Scholar] [CrossRef]
- Scheller, J.; Chalaris, A.; Garbers, C.; Rose-John, S. ADAM17: A molecular switch to control inflammation and tissue regeneration. Trends Immunol. 2011, 32, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Brazzell, J.; Herrera, A.; Walcheck, B. ADAM17 deficiency by mature neutrophils has differential effects on L-selectin shedding. Blood 2006, 108, 2275–2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murthy, A.; Defamie, V.; Smookler, D.S.; Di Grappa, M.A.; Horiuchi, K.; Federici, M.; Sibilia, M.; Blobel, C.P.; Khokha, R. Ectodomain shedding of EGFR ligands and TNFR1 dictates hepatocyte apoptosis during fulminant hepatitis in mice. J. Clin. Investig. 2010, 120, 2731–2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scannevin, R.H.; Alexander, R.; Haarlander, T.M.; Burke, S.L.; Singer, M.; Huo, C.; Zhang, Y.-M.; Maguire, D.; Spurlino, J.; Deckman, I.; et al. Discovery of a highly selective chemical inhibitor of matrix metalloproteinase-9 (MMP-9) that allosterically inhibits zymogen activation. J. Biol. Chem. 2017, 292, 17963–17974. [Google Scholar] [CrossRef]
- Kiaei, M.; Kipiani, K.; Calingasan, N.Y.; Wille, E.; Chen, J.; Heissig, B.; Rafii, S.; Lorenzl, S.; Beal, M.F. Matrix metalloproteinase-9 regulates TNF-alpha and FasL expression in neuronal, glial cells and its absence extends life in a transgenic mouse model of amyotrophic lateral sclerosis. Exp. Neurol. 2007, 205, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Vaday, G.G.; Hershkoviz, R.; Rahat, M.A.; Lahat, N.; Cahalon, L.; Lider, O. Fibronectin-bound TNF-alpha stimulates monocyte matrix metalloproteinase-9 expression and regulates chemotaxis. J. Leukoc. Biol. 2000, 68, 737–747. [Google Scholar] [PubMed]
- Martínez, E.; Fernández-Ruíz, I.; Fresno, C.; Soares-Schanoski, A.; Jurado, T.; Siliceo, M.; Toledano, V.; Fernández-Palomares, R.; García-Rio, F.; et al. Role of MMPs in orchestrating inflammatory response in human monocytes via a TREM-1-PI3K-NF-kappaB pathway. J. Leukoc. Biol. 2012, 91, 933–945. [Google Scholar]
- Gendron, R.; Grenier, D.; Sorsa, T.; Mayrand, D. Inhibition of the activities of matrix metalloproteinases 2, 8, and 9 by chlorhexidine. Clin. Diagn. Lab. Immunol. 1999, 6, 437–439. [Google Scholar] [PubMed]
- Eissner, G.; Kolch, W.; Scheurich, P. Ligands working as receptors: Reverse signaling by members of the TNF superfamily enhance the plasticity of the immune system. Cytokine Growth Factor Rev. 2004, 15, 353–366. [Google Scholar] [CrossRef]
- Schutze, S.; Schneider-Brachert, W. Impact of TNF-R1 and CD95 internalization on apoptotic and antiapoptotic signaling. Results Probl. Cell Differ. 2009, 49, 63–85. [Google Scholar]
- Suda, T.; Hashimoto, H.; Tanaka, M.; Ochi, T.; Nagata, S. Membrane Fas ligand kills human peripheral blood T lymphocytes, and soluble Fas ligand blocks the killing. J. Exp. Med. 1997, 186, 2045–2050. [Google Scholar] [CrossRef] [PubMed]
- Dockrell, D.H.; Lee, M.; Lynch, D.H.; Read, R.C. Immune-mediated phagocytosis and killing of Streptococcus pneumoniae are associated with direct and bystander macrophage apoptosis. J. Infect. Dis. 2001, 184, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.B.; Savill, J. Phagocytosis triggers macrophage release of Fas ligand and induces apoptosis of bystander leukocytes. J. Immunol. 1999, 162, 480–485. [Google Scholar] [PubMed]
- Holm, G.H.; Zhang, C.; Gorry, P.R.; Peden, K.; Schols, D.; De Clercq, E.; Gabuzda, D. Apoptosis of bystander T cells induced by human immunodeficiency virus type 1 with increased envelope/receptor affinity and coreceptor binding site exposure. J. Virol. 2004, 78, 4541–4551. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R.; Janeway, C., Jr. Innate immunity. N. Engl. J. Med. 2000, 343, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Yang, F.; Yu, L.; Yu, Z.; Jiang, L.; Wang, Q.; Yang, Y.; Wang, L.; Cao, X.; Wang, J. Activated T cell exosomes promote tumor invasion via Fas signaling pathway. J. Immunol. 2012, 188, 5954–5961. [Google Scholar] [CrossRef]
- Scott, A.J.; O’Dea, K.P.; O’Callaghan, D.; Williams, L.; Dokpesi, J.O.; Tatton, L.; Handy, J.M.; Hogg, P.J.; Takata, M. Reactive oxygen species and p38 mitogen-activated protein kinase mediate tumor necrosis factor alpha-converting enzyme (TACE/ADAM-17) activation in primary human monocytes. J. Biol. Chem. 2011, 286, 35466–35476. [Google Scholar] [CrossRef]
- Black, R.A.; Rauch, C.T.; Kozlosky, C.J.; Peschon, J.J.; Slack, J.L.; Wolfson, M.F.; Castner, B.J.; Stocking, K.L.; Reddy, P.; Srinivasan, S.; et al. A metalloproteinase disintegrin that releases tumour-necrosis factor-alpha from cells. Nature 1997, 385, 729–733. [Google Scholar] [CrossRef]
- Walcheck, B.; Herrera, A.H.; St Hill, C.; Mattila, P.E.; Whitney, A.R.; Deleo, F.R. ADAM17 activity during human neutrophil activation and apoptosis. Eur. J. Immunol. 2006, 36, 968–976. [Google Scholar] [CrossRef] [Green Version]
- Kirchner, S.; Boldt, S.; Kolch, W.; Haffner, S.; Kazak, S.; Janosch, P.; Holler, E.; Andreesen, R.; Eissner, G. LPS resistance in monocytic cells caused by reverse signaling through transmembrane TNF (mTNF) is mediated by the MAPK/ERK pathway. J. Leukoc. Biol. 2004, 75, 324–331. [Google Scholar] [CrossRef]
- Muthukuru, M.; Cutler, C.W. Resistance of MMP9 and TIMP1 to endotoxin tolerance. Pathog. Dis. 2015, 73, ftu003. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Karakiulakis, G.; Batzios, S.; Savic, S.; Roth, M.; Tamm, M.; Stolz, D. Acute exacerbations of COPD are associated with significant activation of matrix metalloproteinase 9 irrespectively of airway obstruction, emphysema and infection. Respir. Res. 2015, 16, 78. [Google Scholar] [CrossRef] [PubMed]
- Sakthivel, P.; Breithaupt, A.; Gereke, M.; Copland, D.A.; Schulz, C.; Gruber, A.D.; Dick, A.D.; Schreiber, J.; Bruder, D. Soluble CD200 Correlates with Interleukin-6 Levels in Sera of COPD Patients: Potential Implication of the CD200/CD200R Axis in the Disease Course. Lung 2017, 195, 59–68. [Google Scholar] [CrossRef]
- Sukhikh, G.T.; Kan, N.E.; Tyutyunnik, V.L.; Sannikova, M.V.; Dubova, E.A.; Pavlov, K.A.; Amiraslanov, E.Y.; Dolgushina, N.V. The role of extracellular inducer of matrix metalloproteinases in premature rupture of membranes. J. Matern.-Fetal Neonatal Med. 2016, 29, 656–659. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, M.F.; Smith, C.M.; Weiss, S.L.; Dawson, S.; Ralay Ranaivo, H.; Wainwright, M.S. Evaluation of New Diagnostic Biomarkers in Pediatric Sepsis: Matrix Metalloproteinase-9, Tissue Inhibitor of Metalloproteinase-1, Mid-Regional Pro-Atrial Natriuretic Peptide, and Adipocyte Fatty-Acid Binding Protein. PLoS ONE 2016, 11, e0153645. [Google Scholar] [CrossRef]
- Milla, M.E.; Gonzales, P.E.; Leonard, J.D. The TACE zymogen: Re-examining the role of the cysteine switch. Cell Biochem. Biophys. 2006, 44, 342–348. [Google Scholar] [CrossRef]
- Lee, J.H.; Choi, Y.J.; Heo, S.H.; Lee, J.M.; Cho, J.Y. Tumor necrosis factor-alpha converting enzyme (TACE) increases RANKL expression in osteoblasts and serves as a potential biomarker of periodontitis. BMB Rep. 2011, 44, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Mohan, M.J.; Seaton, T.; Mitchell, J.; Howe, A.; Blackburn, K.; Burkhart, W.; Moyer, M.; Patel, I.; Waitt, G.M.; Becherer, J.D.; et al. The tumor necrosis factor-alpha converting enzyme (TACE): A unique metalloproteinase with highly defined substrate selectivity. Biochemistry 2002, 41, 9462–9469. [Google Scholar] [CrossRef] [PubMed]
- Tapader, R.; Bose, D.; Pal, A. YghJ, the secreted metalloprotease of pathogenic E. coli induces hemorrhagic fluid accumulation in mouse ileal loop. Microb. Pathog. 2017, 105, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Stinemetz, E.K.; Gao, P.; Pinkston, K.L.; Montealegre, M.C.; Murray, B.E.; Harvey, B.R. Processing of the major autolysin of E. faecalis, AtlA, by the zinc-metalloprotease, GelE, impacts AtlA septal localization and cell separation. PLoS ONE 2017, 12, e0186706. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dreschers, S.; Platen, C.; Ludwig, A.; Gille, C.; Köstlin, N.; Orlikowsky, T.W. Metalloproteinases TACE and MMP-9 Differentially Regulate Death Factors on Adult and Neonatal Monocytes After Infection with Escherichia coli. Int. J. Mol. Sci. 2019, 20, 1399. https://doi.org/10.3390/ijms20061399
Dreschers S, Platen C, Ludwig A, Gille C, Köstlin N, Orlikowsky TW. Metalloproteinases TACE and MMP-9 Differentially Regulate Death Factors on Adult and Neonatal Monocytes After Infection with Escherichia coli. International Journal of Molecular Sciences. 2019; 20(6):1399. https://doi.org/10.3390/ijms20061399
Chicago/Turabian StyleDreschers, Stephan, Christopher Platen, Andreas Ludwig, Christian Gille, Natascha Köstlin, and Thorsten W. Orlikowsky. 2019. "Metalloproteinases TACE and MMP-9 Differentially Regulate Death Factors on Adult and Neonatal Monocytes After Infection with Escherichia coli" International Journal of Molecular Sciences 20, no. 6: 1399. https://doi.org/10.3390/ijms20061399
APA StyleDreschers, S., Platen, C., Ludwig, A., Gille, C., Köstlin, N., & Orlikowsky, T. W. (2019). Metalloproteinases TACE and MMP-9 Differentially Regulate Death Factors on Adult and Neonatal Monocytes After Infection with Escherichia coli. International Journal of Molecular Sciences, 20(6), 1399. https://doi.org/10.3390/ijms20061399