Autophosphorylation of Orphan Receptor ERBB2 Can Be Induced by Extracellular Treatment with Mildly Alkaline Media


 ,
, 
Abstract
:1. Introduction
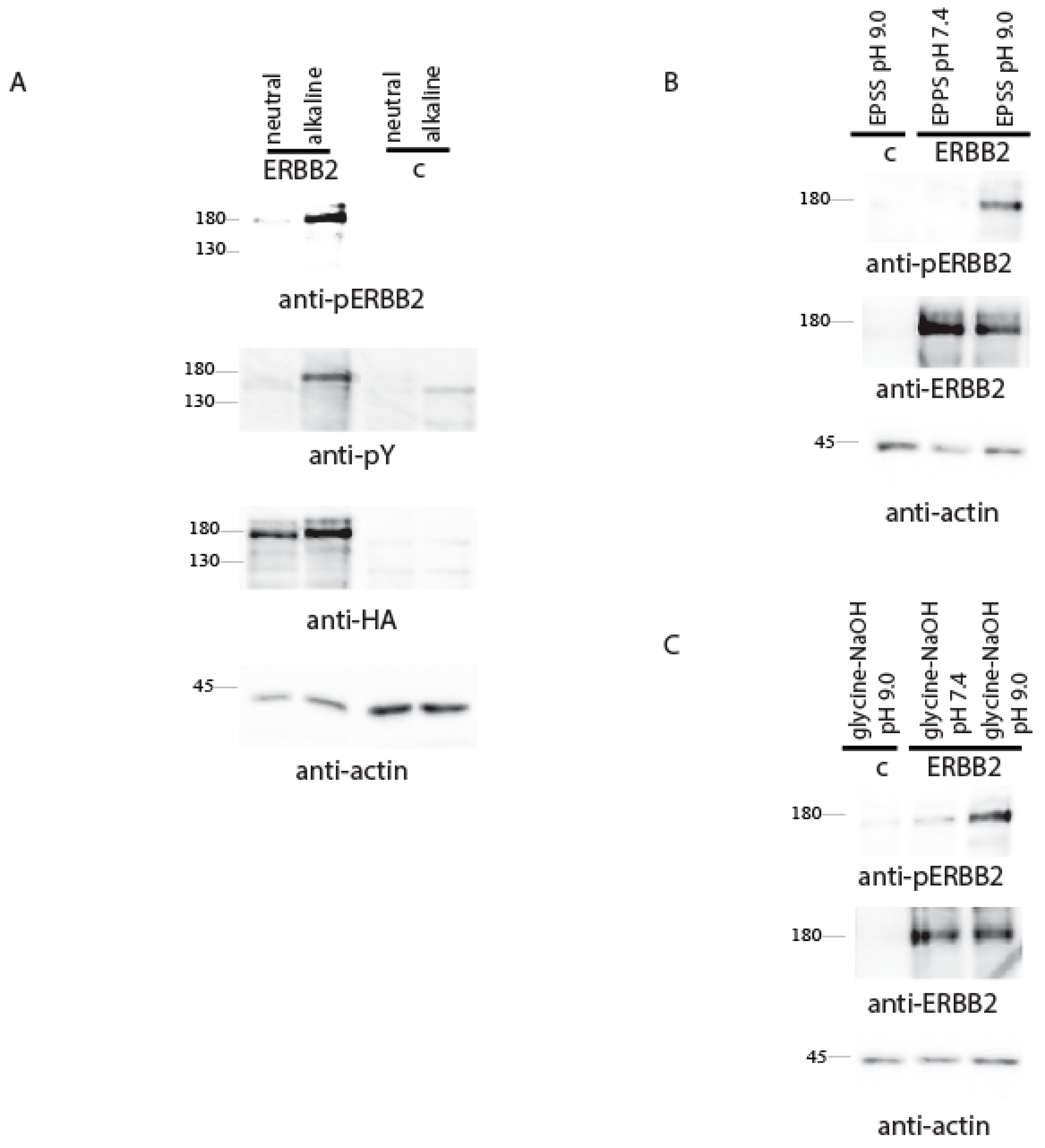
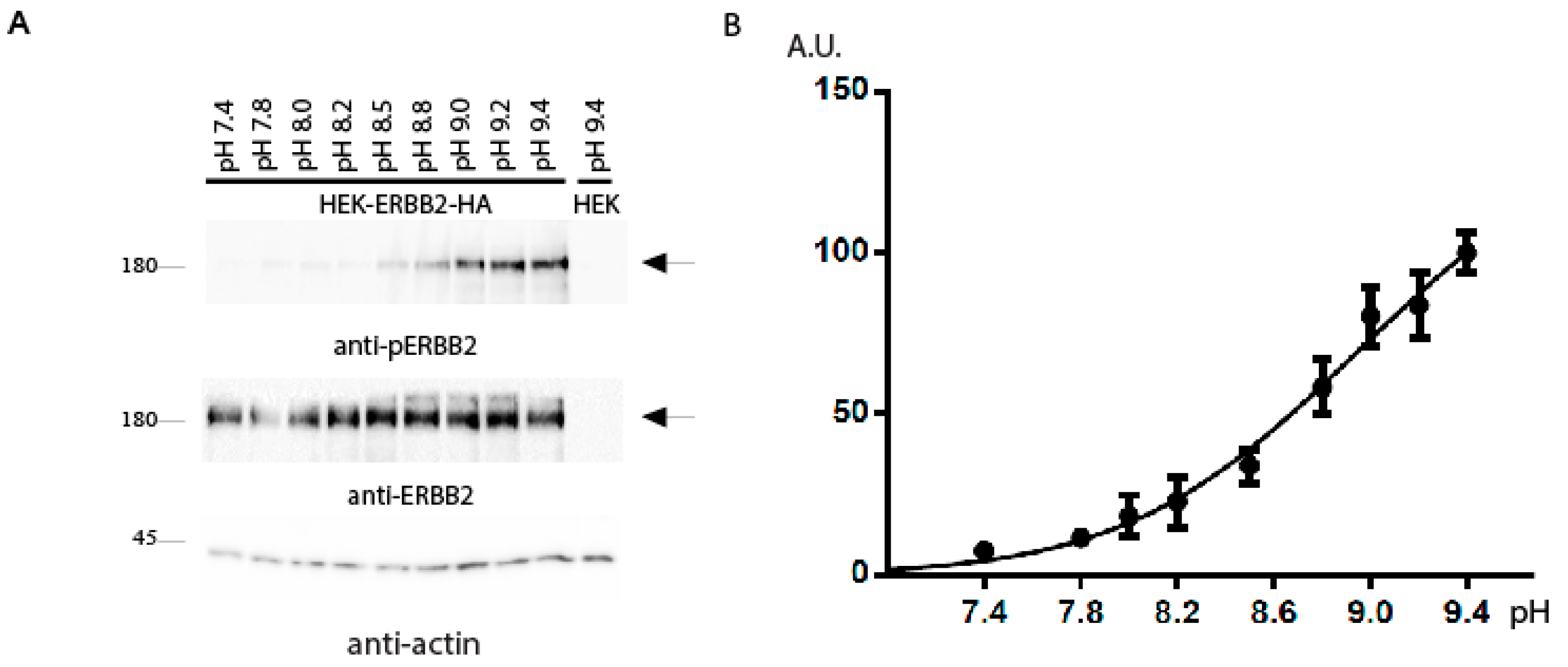
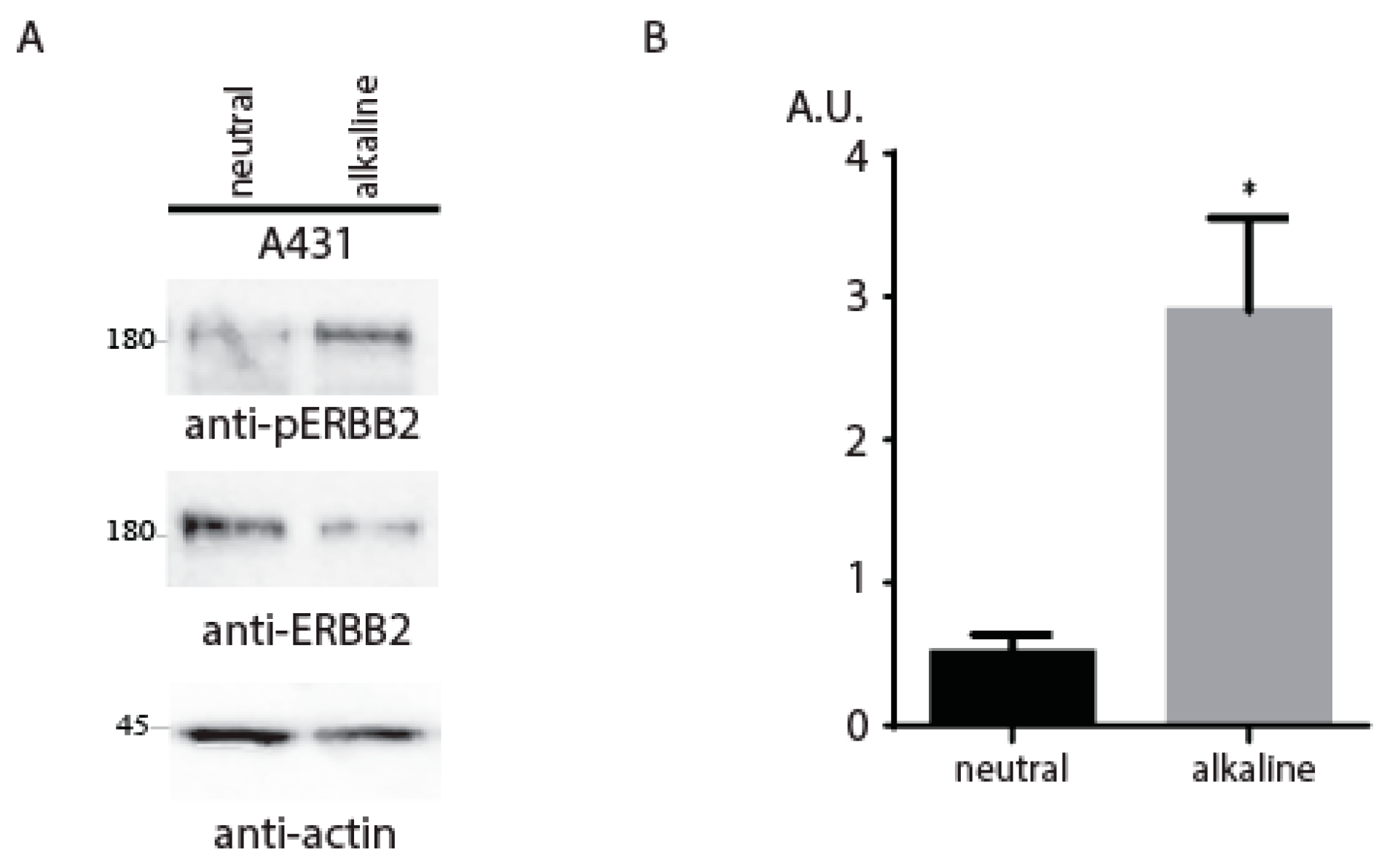
2. Results and Discussion
3. Materials and Methods
3.1. Cell Lines and Treatments
3.2. Antibodies and Immunoblotting
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bocharov, E.V.; Lesovoy, D.M.; Pavlov, K.V.; Pustovalova, Y.E.; Bocharova, O.V.; Arseniev, A.S. Alternative packing of EGFR transmembrane domain suggests that protein–lipid interactions underlie signal conduction across membrane. Biochim. Biophys. Acta 2016, 1858, 1254–1261. [Google Scholar] [CrossRef]
- Lemmon, M.A. Ligand-induced ErbB receptor dimerization. Exp. Cell Res. 2009, 315, 638–648. [Google Scholar] [CrossRef] [PubMed]
- Britsch, S. The neuregulin-I/ErbB signaling system in development and disease. Adv. Anat. Embryol. Cell Biol. 2007, 190, 1–65. [Google Scholar]
- Burgess, A.W. EGFR family: Structure physiology signalling and therapeutic targets†. Growth Factors 2008, 26, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Gebhart, G.; Flamen, P.; De Vries, E.G.E.; Jhaveri, K.; Wimana, Z. Imaging Diagnostic and Therapeutic Targets: Human Epidermal Growth Factor Receptor 2. J. Nuclear Med. 2016, 57, 81S–88S. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.T.; Chau, C.H.; Price, D.K.; Figg, W.D. Precision Oncology Medicine: The Clinical Relevance of Patient-Specific Biomarkers Used to Optimize Cancer Treatment. J. Clin. Pharmacol. 2016, 56, 1484–1499. [Google Scholar] [CrossRef] [PubMed]
- Deev, I.E.; Vasilenko, K.P.; Kurmangaliev, E.Z.; Serova, O.V.; Popova, N.V.; Galagan, Y.S.; Burova, E.B.; Zozulya, S.A.; Nikol’skii, N.N.; Petrenko, A.G. Effect of changes in ambient pH on phosphorylation of cellular proteins. Dokl. Biochem. Biophys. 2006, 408, 184–187. [Google Scholar] [CrossRef]
- Deyev, I.E.; Sohet, F.; Vassilenko, K.P.; Serova, O.V.; Popova, N.V.; Zozulya, S.A.; Burova, E.B.; Houillier, P.; Rzhevsky, D.I.; Berchatova, A.A.; et al. Insulin receptor-related receptor as an extracellular alkali sensor. Cell Metab. 2011, 13, 679–689. [Google Scholar] [CrossRef]
- Lin, H.; Chen, W.; Ding, H. AcalPred: A sequence-based tool for discriminating between acidic and alkaline enzymes. PLoS ONE 2013, 8, e75726. [Google Scholar] [CrossRef]
- Deyev, I.E.; Popova, N.V.; Petrenko, A.G. Determination of alkali-sensing parts of the insulin receptor-related receptor using the bioinformatic approach. Acta Nat. 2015, 7, 80–86. [Google Scholar]
- Natali, P.G.; Nicotra, M.R.; Bigotti, A.; Venturo, I.; Slamon, D.J.; Fendly, B.M.; Ullrich, A. Expression of the p185 encoded by HER2 oncogene in normal and transformed human tissues. Int. J. Cancer 1990, 45, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.A.; Weiner, D.B.; More, K.F.; Kokai, Y.; Williams, W.V.; Maguire, H.C.; LiVolsi, V.A.; Greene, M.I. Expression pattern of the neu (NGL) gene-encoded growth factor receptor protein (p185neu) in normal and transformed epithelial tissues of the digestive tract. Oncogene 1989, 4, 81–88. [Google Scholar] [PubMed]
- Mori, S.; Akiyama, T.; Yamada, Y.; Morishita, Y.; Sugawara, I.; Toyoshima, K.; Yamamoto, T. C-erbB-2 gene product, a membrane protein commonly expressed on human fetal epithelial cells. Lab. Investig. J. Tech. Methods Pathol. 1989, 61, 93–97. [Google Scholar]
- Petrenko, A.G.; Zozulya, S.A.; Deyev, I.E.; Eladari, D. Insulin receptor-related receptor as an extracellular pH sensor involved in the regulation of acid-base balance. Biochim. Biophys. Acta Proteins Proteom. 2013, 1834. [Google Scholar] [CrossRef]
- Feher, J.J. Quantitative Human Physiology: An Introduction; Academic Press: Cambridge, MA, USA, 2017; ISBN 9780128011546. [Google Scholar]
- Young, J.A.; Schneyer, C.A. Composition of saliva in mammalia. Aust. J. Exp. Biol. Med. Sci. 1981, 59, 1–53. [Google Scholar] [CrossRef]
- Lavy, E.; Goldberger, D.; Friedman, M.; Steinberg, D. pH values and mineral content of saliva in different breeds of dogs. Israel J. Vet. Med. 2012, 67, 244–248. [Google Scholar]
- Iacopetti, I.; Perazzi, A.; Badon, T.; Bedin, S.; Contiero, B.; Ricci, R. Salivary pH, calcium, phosphorus and selected enzymes in healthy dogs: A pilot study. BMC Vet. Res. 2017, 13, 330. [Google Scholar] [CrossRef]
- McDougall, C.J.; Wong, R.; Scudera, P.; Lesser, M.; DeCosse, J.J. Colonic mucosal pH in humans. Dig. Dis. Sci. 1993, 38, 542–545. [Google Scholar] [CrossRef]
- Ward, F.W.; Coates, M.E. Gastrointestinal pH measurement in rats: Influence of the microbial flora, diet and fasting. Lab. Anim. 1987, 21, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Deyev, I.E.; Popova, N.V.; Serova, O.V.; Zhenilo, S.V.; Regoli, M.; Bertelli, E.; Petrenko, A.G. Alkaline pH induces IRR-mediated phosphorylation of IRS-1 and actin cytoskeleton remodeling in a pancreatic beta cell line. Biochimie 2017, 138, 62–69. [Google Scholar] [CrossRef]
- Lee, K.F.; Simon, H.; Chen, H.; Bates, B.; Hung, M.C.; Hauser, C. Requirement for neuregulin receptor erbB2 in neural and cardiac development. Nature 1995, 378, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Erickson, S.L.; O’Shea, K.S.; Ghaboosi, N.; Loverro, L.; Frantz, G.; Bauer, M.; Lu, L.H.; Moore, M.W. ErbB3 is required for normal cerebellar and cardiac development: A comparison with ErbB2-and heregulin-deficient mice. Development 1997, 124, 4999–5011. [Google Scholar] [PubMed]
- Reinhardt, R.R.; Chin, E.; Zhang, B.; Roth, R.A.; Bondy, C.A. Selective coexpression of insulin receptor-related receptor (IRR) and TRK in NGF-sensitive neurons. J. Neurosci. 1994, 14, 4674–4683. [Google Scholar] [CrossRef]
- Pearson, R.J.; Carroll, S.L. ErbB transmembrane tyrosine kinase receptors are expressed by sensory and motor neurons projecting into sciatic nerve. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2004, 52, 1299–1311. [Google Scholar] [CrossRef] [PubMed]
- Krasnoperov, V.; Deyev, I.E.; Serova, O.V.; Xu, C.; Lu, Y.; Buryanovsky, L.; Gabibov, A.G.; Neubert, T.A.; Petrenko, A.G. Dissociation of the Subunits of the calcium-independent receptor of α-Latrotoxin as a result of two-step proteolysis. Biochemistry 2009, 48, 3230–3238. [Google Scholar] [CrossRef]


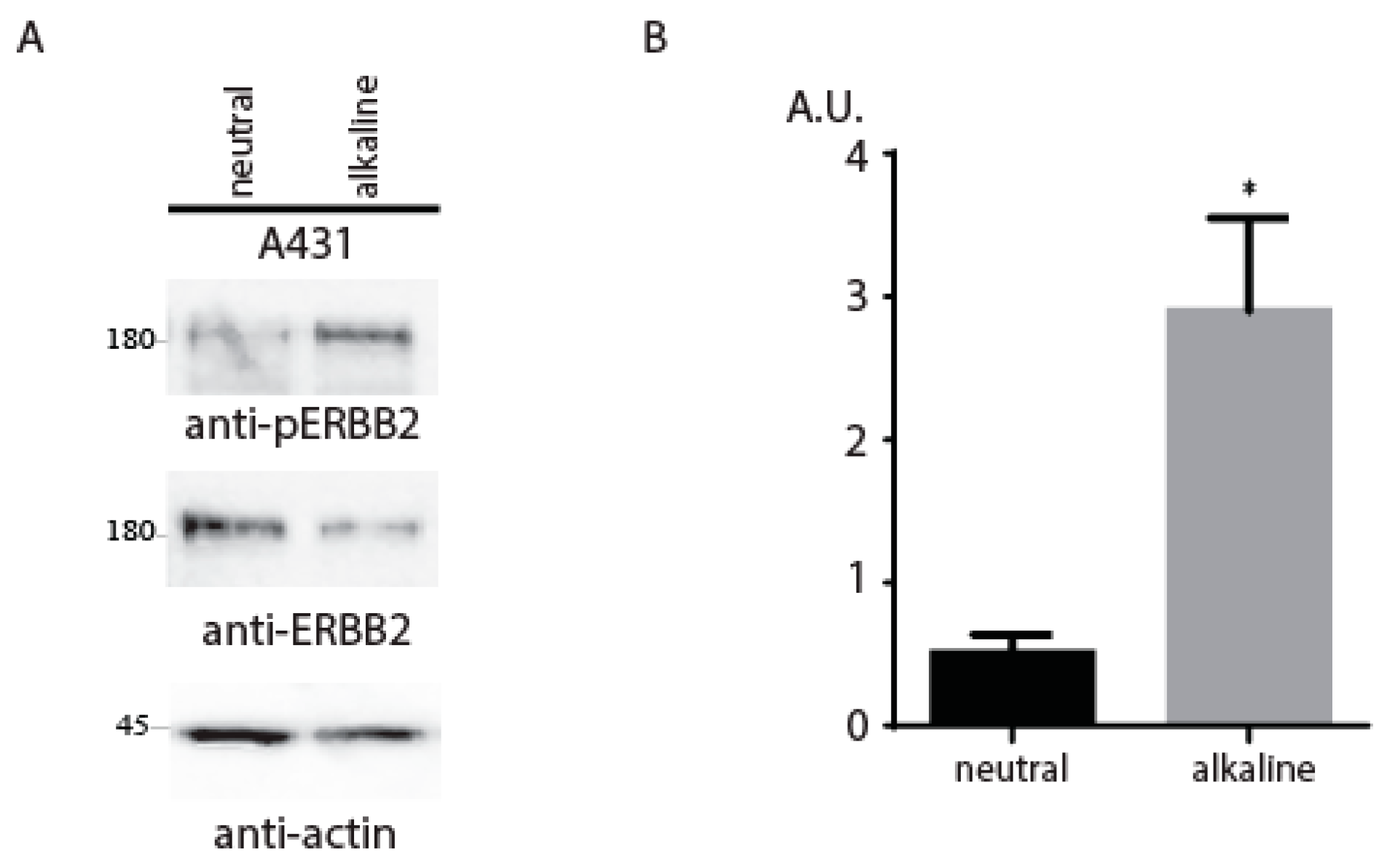

{kind=link}
{kind=link}
{kind=link}
| Receptor | Xenopus | Mouse | Rat | Dog | Chicken | Human |
|---|---|---|---|---|---|---|
| ERBB2 | 0.83 | 0.806 | 0.838 | 0.773 | 0.633 | 0.696 |
| ERBB3 | 0.622 | 0.853 | 0.746 | 0.807 | 0.595 | |
| INSRR | 0.573 | 0.798 | 0.912 | 0.839 | 0.546 | 0.726 |
| PDGFRB | 0.819 | 0.111 | 0.141 | 0.786 | 0.641 | 0.874 |
| FLT3 | 0.829 | 0.834 | 0.767 | 0.844 | 0.944 | 0.947 |
| VEGFR1 | 0.909 | 0.905 | 0.924 | 0.976 | ||
| CCK4 | 0.975 | 0.892 | 0.923 | 0.989 | 0.803 | |
| MST1R | 0.808 | 0.819 | 0.847 | 0.877 | 0.464 | 0.634 |
| EPHB4 | 0.888 | 0.905 | 0.916 | 0.594 | 0.685 | |
| TIE1 | 0.564 | 0.358 | 0.464 | 0.608 | 0.33 | 0.787 |
| TIE2 | 0.978 | 0.896 | 0.905 | 0.854 | 0.539 | 0.888 |
| DDR2 | 0.612 | 0.657 | 0.688 | 0.367 | 0.965 | 0.646 |
| RET | 0.258 | 0.618 | 0.381 | 0.739 | 0.173 | 0.667 |
| ROR1 | 0.953 | 0.856 | 0.969 | 0.227 | 0.771 | 0.939 |
| ROR2 | 0.232 | 0.679 | 0.554 | 0.699 | 0.9 | 0.793 |
| MUSK | 0.693 | 0.99 | 0.984 | 0.947 | 0.908 | 0.972 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serova, O.V.; Chachina, N.A.; Gantsova, E.A.; Popova, N.V.; Petrenko, A.G.; Deyev, I.E. Autophosphorylation of Orphan Receptor ERBB2 Can Be Induced by Extracellular Treatment with Mildly Alkaline Media. Int. J. Mol. Sci. 2019, 20, 1515. https://doi.org/10.3390/ijms20061515
Serova OV, Chachina NA, Gantsova EA, Popova NV, Petrenko AG, Deyev IE. Autophosphorylation of Orphan Receptor ERBB2 Can Be Induced by Extracellular Treatment with Mildly Alkaline Media. International Journal of Molecular Sciences. 2019; 20(6):1515. https://doi.org/10.3390/ijms20061515
Chicago/Turabian StyleSerova, Oxana V., Natalia A. Chachina, Elena A. Gantsova, Nadezhda V. Popova, Alexander G. Petrenko, and Igor E. Deyev. 2019. "Autophosphorylation of Orphan Receptor ERBB2 Can Be Induced by Extracellular Treatment with Mildly Alkaline Media" International Journal of Molecular Sciences 20, no. 6: 1515. https://doi.org/10.3390/ijms20061515
APA StyleSerova, O. V., Chachina, N. A., Gantsova, E. A., Popova, N. V., Petrenko, A. G., & Deyev, I. E. (2019). Autophosphorylation of Orphan Receptor ERBB2 Can Be Induced by Extracellular Treatment with Mildly Alkaline Media. International Journal of Molecular Sciences, 20(6), 1515. https://doi.org/10.3390/ijms20061515