Mitogen-Activated Protein Kinase Phosphatases (MKPs) in Fungal Signaling: Conservation, Function, and Regulation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
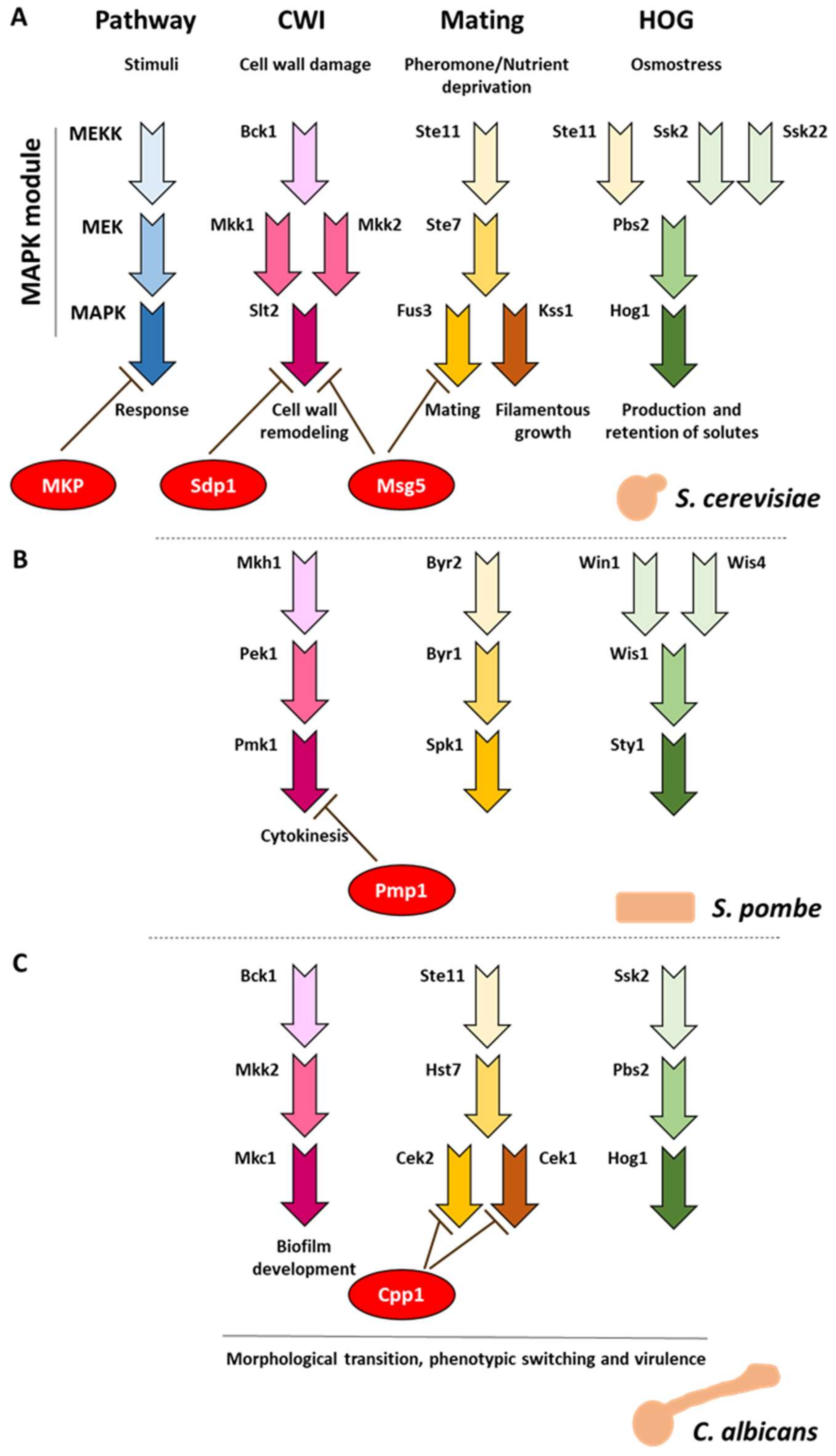
:1. Fungi Respond to Distinct Stimuli Through MAPK Pathways
2. General Structure and Essential Motifs of S. cerevisiae MKPs
3. Structural Conservation of Fungal MKPs
4. Function of Fungal MKPs
4.1. Msg5 Regulates the S. cerevisiae Mating Pathway by Targeting Fus3
4.2. CWI MAPK Slt2 is Regulated by the MKPs Msg5 and Sdp1 in S. cerevisiae
4.3. C. albicans MKP Cpp1 Participates in Phenotypic Transition, Mating, and Virulence
4.4. S. pombe Pmp1: The Only Characterized MKP Within the Subphylum Taphrinomycotina
4.5. Filamentous Fungi MKPs are Involved in Mycelial Growth and Virulence
4.6. Choose the Right Host: A Role for MKPs Within Basidiomycota
5. Regulatory Mechanisms of Fungal MKPs
6. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MAPK | Mitogen Activated Protein Kinase |
| MKP | MAP Kinase phosphatase |
| DUSP | Dual Specificity Phosphatase |
References
- Papa, S.; Choy, P.M.; Bubici, C. The ERK and JNK pathways in the regulation of metabolic reprogramming. Oncogene 2018, 10, 0582. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, W.Z.; Liu, T.; Feng, X.; Yang, N.; Zhou, H.F. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis. J. Recept. Signal. Transduct. Res. 2015, 35, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef]
- Low, H.B.; Zhang, Y. Regulatory Roles of MAPK Phosphatases in Cancer. Immune. Netw. 2016, 16, 85–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohush, A.; Niewiadomska, G.; Filipek, A. Role of Mitogen Activated Protein Kinase Signaling in Parkinson’s Disease. Int. J. Mol. Sci. 2018, 19, 2973. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Choi, E.J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta 2010, 1802, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Perez-Nadales, E.; Nogueira, M.F.; Baldin, C.; Castanheira, S.; El Ghalid, M.; Grund, E.; Lengeler, K.; Marchegiani, E.; Mehrotra, P.V.; Moretti, M.; et al. Fungal model systems and the elucidation of pathogenicity determinants. Fungal. Genet. Biol. 2014, 70, 42–67. [Google Scholar] [CrossRef] [Green Version]
- Hamel, L.P.; Nicole, M.C.; Duplessis, S.; Ellis, B.E. Mitogen-activated protein kinase signaling in plant-interacting fungi: Distinct messages from conserved messengers. Plant Cell. 2012, 24, 1327–1351. [Google Scholar] [CrossRef]
- Chen, R.E.; Thorner, J. Function and regulation in MAPK signaling pathways: Lessons learned from the yeast Saccharomyces cerevisiae. Biochim. Biophys. Acta 2007, 1773, 1311–1340. [Google Scholar] [CrossRef]
- Engelberg, D.; Perlman, R.; Levitzki, A. Transmembrane signaling in Saccharomyces cerevisiae as a model for signaling in metazoans: State of the art after 25 years. Cell Signal. 2014, 26, 2865–2878. [Google Scholar] [CrossRef]
- Perez, P.; Cansado, J. Cell integrity signaling and response to stress in fission yeast. Curr. Protein Pept. Sci. 2010, 11, 680–692. [Google Scholar] [CrossRef] [PubMed]
- Roman, E.; Arana, D.M.; Nombela, C.; Alonso-Monge, R.; Pla, J. MAP kinase pathways as regulators of fungal virulence. Trends Microbiol. 2007, 15, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.M.; Feng, M.G. Insights into regulatory roles of MAPK-cascaded pathways in multiple stress responses and life cycles of insect and nematode mycopathogens. Appl. Microbiol. Biotechnol. 2019, 103, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Rispail, N.; Soanes, D.M.; Ant, C.; Czajkowski, R.; Grünler, A.; Huguet, R.; Perez-Nadales, E.; Poli, A.; Sartorel, E.; Valiante, V.; et al. Comparative genomics of MAP kinase and calcium-calcineurin signalling components in plant and human pathogenic fungi. Fungal. Genet. Biol. 2009, 46, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, B.; Soto, T.; del Dedo, J.E.; Franco, A.; Vicente, J.; Hidalgo, E.; Gacto, M.; Cansado, J.; Madrid, M. Distinct biological activity of threonine monophosphorylated MAPK isoforms during the stress response in fission yeast. Cell Signal. 2015, 27, 2534–2542. [Google Scholar] [CrossRef] [PubMed]
- Nagiec, M.J.; McCarter, P.C.; Kelley, J.B.; Dixit, G.; Elston, T.C.; Dohlman, H.G. Signal inhibition by a dynamically regulated pool of monophosphorylated MAPK. Mol. Biol. Cell. 2015, 26, 3359–3371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askari, N.; Beenstock, J.; Livnah, O.; Engelberg, D. p38alpha is active in vitro and in vivo when monophosphorylated at threonine 180. Biochemistry 2009, 48, 2497–2504. [Google Scholar] [CrossRef]
- Caunt, C.J.; Keyse, S.M. Dual-specificity MAP kinase phosphatases (MKPs): Shaping the outcome of MAP kinase signalling. FEBS J. 2013, 280, 489–504. [Google Scholar] [CrossRef]
- Pulido, R.; Zúñiga, A.; Ullrich, A. PTP-SL and STEP protein tyrosine phosphatases regulate the activation of the extracellular signal-regulated kinases ERK1 and ERK2 by association through a kinase interaction motif. EMBO J. 1998, 17, 7337–7350. [Google Scholar] [CrossRef] [PubMed]
- Tanoue, T.; Nishida, E. Molecular recognitions in the MAP kinase cascades. Cell Signal. 2003, 15, 455–462. [Google Scholar] [CrossRef]
- Peti, W.; Page, R. Molecular basis of MAP kinase regulation. Protein Sci. 2013, 22, 1698–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, H.; Flandez, M.; Nombela, C.; Molina, M. Protein phosphatases in MAPK signalling: We keep learning from yeast. Mol. Microbiol. 2005, 58, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Kellis, M.; Birren, B.W.; Lander, E.S. Proof and evolutionary analysis of ancient genome duplication in the yeast Saccharomyces cerevisiae. Nature 2004, 428, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Gartner, A.; Ammerer, G.; Errede, B.; Shinkawa, H.; Sugimoto, K.; Matsumoto, K. MSG5, a novel protein phosphatase promotes adaptation to pheromone response in S. cerevisiae. EMBO J. 1994, 13, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Flandez, M.; Cosano, I.C.; Nombela, C.; Martin, H.; Molina, M. Reciprocal regulation between Slt2 MAPK and isoforms of Msg5 dual-specificity protein phosphatase modulates the yeast cell integrity pathway. J. Biol. Chem. 2004, 279, 11027–11034. [Google Scholar] [CrossRef]
- Palacios, L.; Dickinson, R.J.; Sacristan-Reviriego, A.; Didmon, M.P.; Marín, M.J.; Martín, H.; Keyse, S.M.; Molina, M. Distinct docking mechanisms mediate interactions between the Msg5 phosphatase and mating or cell integrity mitogen-activated protein kinases (MAPKs) in Saccharomyces cerevisiae. J. Biol. Chem. 2011, 286, 42037–42050. [Google Scholar] [CrossRef] [PubMed]
- Sacristan-Reviriego, A.; Madrid, M.; Cansado, J.; Martin, H.; Molina, M. A conserved non-canonical docking mechanism regulates the binding of dual specificity phosphatases to cell integrity mitogen-activated protein kinases (MAPKs) in budding and fission yeasts. PLoS ONE 2014, 9, e85390. [Google Scholar] [CrossRef] [PubMed]
- Fox, G.C.; Shafiq, M.; Briggs, D.C.; Knowles, P.P.; Collister, M.; Didmon, M.J.; Makrantoni, V.; Dickinson, R.J.; Hanrahan, S.; Totty, N.; et al. Redox-mediated substrate recognition by Sdp1 defines a new group of tyrosine phosphatases. Nature 2007, 447, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.J.; Peng, J.; Li, Q.X.; Peng, Y.L. Phosphorylation-mediated Regulatory Networks in Mycelia of Pyricularia oryzae Revealed by Phosphoproteomic Analyses. Mol. Cell Proteomics 2017, 16, 1669–1682. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.L.; Deschenes, R.J.; Guan, K.L. Differential regulation of FUS3 MAP kinase by tyrosine-specific phosphatases PTP2/PTP3 and dual-specificity phosphatase MSG5 in Saccharomyces cerevisiae. Genes Dev. 1997, 11, 1690–1702. [Google Scholar] [CrossRef]
- Dyjack, N.; Azeredo-Tseng, C.; Yildirim, N. Mathematical modeling reveals differential regulation of MAPK activity by phosphatase proteins in the yeast pheromone response pathway. Mol. Biosyst. 2017, 13, 1323–1335. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, E.; Halatek, I.M.; Kim, H.J.; Ellicott, A.T.; Obukhov, A.A.; Stone, D.E. Effect of the pheromone-responsive G(alpha) and phosphatase proteins of Saccharomyces cerevisiae on the subcellular localization of the Fus3 mitogen-activated protein kinase. Mol. Cell Biol. 2003, 23, 1135–1150. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, E.; Kim, H.J.; Stone, D.E. The pheromone-induced nuclear accumulation of the Fus3 MAPK in yeast depends on its phosphorylation state and on Dig1 and Dig2. BMC Cell Biol. 2007, 8, 44–48. [Google Scholar] [CrossRef]
- Marin, M.J.; Flandez, M.; Bermejo, C.; Arroyo, J.; Martin, H.; Molina, M. Different modulation of the outputs of yeast MAPK-mediated pathways by distinct stimuli and isoforms of the dual-specificity phosphatase Msg5. Mol. Genet. Genom. 2009, 281, 345–359. [Google Scholar] [CrossRef] [PubMed]
- Andersson, J.; Simpson, D.M.; Qi, M.; Wang, Y.; Elion, E.A. Differential input by Ste5 scaffold and Msg5 phosphatase route a MAPK cascade to multiple outcomes. EMBO J. 2004, 23, 2564–2576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y.; Irie, K.; Matsumoto, K. Yeast RLM1 encodes a serum response factor-like protein that may function downstream of the Mpk1 (Slt2) mitogen-activated protein kinase pathway. Mol. Cell Biol. 1995, 15, 5740–5749. [Google Scholar] [CrossRef]
- Martin, H.; Rodriguez-Pachon, J.M.; Ruiz, C.; Nombela, C.; Molina, M. Regulatory mechanisms for modulation of signaling through the cell integrity Slt2-mediated pathway in Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 1511–1519. [Google Scholar] [CrossRef]
- Tatjer, L.; Sacristan-Reviriego, A.; Casado, C.; González, A.; Rodríguez-Porrata, B.; Palacios, L.; Canadell, D.; Serra-Cardona, A.; Martín, H.; Molina, M.; et al. Wide-Ranging Effects of the Yeast Ptc1 Protein Phosphatase Acting Through the MAPK Kinase Mkk1. Genetics 2016, 202, 141–156. [Google Scholar] [CrossRef]
- Hermansyah; Sugiyama, M.; Kaneko, Y.; Harashima, S. Yeast protein phosphatases Ptp2p and Msg5p are involved in G1-S transition, CLN2 transcription, and vacuole morphogenesis. Arch. Microbiol. 2009, 191, 721–733. [Google Scholar] [CrossRef]
- Liu, L.; Levin, D.E. Intracellular mechanism by which genotoxic stress activates yeast SAPK Mpk1. Mol. Biol. Cell 2018, 29, 2898–2909. [Google Scholar] [CrossRef]
- Hahn, J.S.; Thiele, D.J. Regulation of the Saccharomyces cerevisiae Slt2 kinase pathway by the stress-inducible Sdp1 dual specificity phosphatase. J. Biol. Chem. 2002, 277, 21278–21284. [Google Scholar] [CrossRef] [PubMed]
- Kuravi, V.K.; Kurischko, C.; Puri, M.; Luca, F.C. Cbk1 kinase and Bck2 control MAP kinase activation and inactivation during heat shock. Mol. Biol. Cell 2011, 22, 4892–4907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csank, C.; Makris, C.; Meloche, S.; Schroppel, K.; Röllinghoff, M.; Dignard, D.; Meloche, S.; Thomas, D.Y.; Whiteway, M. Derepressed hyphal growth and reduced virulence in a VH1 family-related protein phosphatase mutant of the human pathogen Candida albicans. Mol. Biol. Cell 1997, 8, 2539–2551. [Google Scholar] [CrossRef] [PubMed]
- Csank, C.; Schroppel, K.; Leberer, E.; Harcus, D.; Mohamed, O.; Meloche, S.; Thomas, D.Y.; Whiteway, M. Roles of the Candida albicans mitogen-activated protein kinase homolog, Cek1p, in hyphal development and systemic candidiasis. Infect. Immun. 1998, 66, 2713–2721. [Google Scholar] [PubMed]
- Schroppel, K.; Sprosser, K.; Whiteway, M.; Thomas, D.Y.; Rollinghoff, M.; Csank, C. Repression of hyphal proteinase expression by the mitogen-activated protein (MAP) kinase phosphatase Cpp1p of Candida albicans is independent of the MAP kinase Cek1p. Infect. Immun. 2000, 68, 7159–7161. [Google Scholar] [CrossRef]
- Deng, F.S.; Lin, C.H. Cpp1 phosphatase mediated signaling crosstalk between Hog1 and Cek1 mitogen-activated protein kinases is involved in the phenotypic transition in Candida albicans. Med. Mycol. 2018, 56, 242–252. [Google Scholar] [CrossRef]
- Rastghalam, G.; Omran, R.P.; Alizadeh, M.; Fulton, D.; Mallick, J.; Whiteway, M. MAP Kinase Regulation of the Candida albicans Pheromone Pathway. mSphere 2019, 4, e00598-18. [Google Scholar] [CrossRef]
- Cheng, S.; Clancy, C.J.; Checkley, M.A.; Handfield, M.; Hillman, J.D.; Progulske-Fox, A.; Lewin, A.S.; Fidel, P.L.; Nguyen, M.H. Identification of Candida albicans genes induced during thrush offers insight into pathogenesis. Mol. Microbiol. 2003, 48, 1275–1288. [Google Scholar] [CrossRef]
- Li, R.; Puri, S.; Tati, S.; Cullen, P.J.; Edgerton, M. Candida albicans Cek1 mitogen-activated protein kinase signaling enhances fungicidal activity of salivary histatin 5. Antimicrob. Agents Chemother. 2015, 59, 3460–3468. [Google Scholar] [CrossRef]
- Sugiura, R.; Toda, T.; Shuntoh, H.; Yanagida, M.; Kuno, T. pmp1+, a suppressor of calcineurin deficiency, encodes a novel MAP kinase phosphatase in fission yeast. EMBO J. 1998, 17, 140–148. [Google Scholar] [CrossRef]
- Madrid, M.; Soto, T.; Khong, H.K.; Franco, A.; Vicente, J.; Pérez, P.; Gacto, M.; Cansado, J. Stress-induced response, localization, and regulation of the Pmk1 cell integrity pathway in Schizosaccharomyces pombe. J. Biol. Chem. 2006, 281, 2033–2043. [Google Scholar] [CrossRef] [PubMed]
- Didmon, M.; Davis, K.; Watson, P.; Ladds, G.; Broad, P.; Davey, J. Identifying regulators of pheromone signalling in the fission yeast Schizosaccharomyces pombe. Curr. Genet. 2002, 41, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Madrid, M.; Nunez, A.; Soto, T.; Vicente-Soler, J.; Gacto, M.; Cansado, J. Stress-activated protein kinase-mediated down-regulation of the cell integrity pathway mitogen-activated protein kinase Pmk1p by protein phosphatases. Mol. Biol. Cell 2007, 18, 4405–4419. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, S.; Hou, R.; Zhao, Z.; Zheng, Q.; Xu, Q.; Zheng, D.; Wang, G.; Liu, H.; Gao, X.; et al. Functional analysis of the kinome of the wheat scab fungus Fusarium graminearum. PLoS Pathog. 2011, 7, e1002460. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Gu, Q.; Yun, Y.; Yin, Y.; Xu, J.R.; Shim, W.B.; Ma, Z. The TOR signaling pathway regulates vegetative development and virulence in Fusarium graminearum. New Phytol. 2014, 203, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.; Liu, Z.; Yin, Y.; Jiang, J.; Chen, Y.; Xu, J.R.; Ma, Z. Functional analysis of the Fusarium graminearum phosphatome. New Phytol. 2015, 207, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.R.; Hamer, J.E. MAP kinase and cAMP signaling regulate infection structure formation and pathogenic growth in the rice blast fungus Magnaporthe grisea. Genes Dev. 1996, 10, 2696–2706. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.R.; Staiger, C.J.; Hamer, J.E. Inactivation of the mitogen-activated protein kinase Mps1 from the rice blast fungus prevents penetration of host cells but allows activation of plant defense responses. Proc. Natl. Acad. Sci. USA 1998, 95, 12713–12718. [Google Scholar] [CrossRef] [Green Version]
- Di Stasio, M.; Brefort, T.; Mendoza-Mendoza, A.; Munch, K.; Kahmann, R. The dual specificity phosphatase Rok1 negatively regulates mating and pathogenicity in Ustilago maydis. Mol. Microbiol. 2009, 73, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Molina, M.; Cid, V.J.; Martin, H. Fine regulation of Saccharomyces cerevisiae MAPK pathways by post-translational modifications. Yeast 2010, 27, 503–511. [Google Scholar] [CrossRef]
- Garcia, R.; Bermejo, C.; Grau, C.; Pérez, R.; Rodríguez-Peña, J.M.; Francois, J.; Nombela, C.; Arroyo, J. The global transcriptional response to transient cell wall damage in Saccharomyces cerevisiae and its regulation by the cell integrity signaling pathway. J. Biol. Chem. 2004, 279, 15183–15195. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, R.; Satoh, R.; Ishiwata, S.; Umeda, N.; Kita, A. Role of RNA-Binding Proteins in MAPK Signal Transduction Pathway. J. Signal. Transduct. 2011, 2011, 109746. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, R.; Kita, A.; Shimizu, Y.; Shuntoh, H.; Sio, S.O.; Kuno, T. Feedback regulation of MAPK signalling by an RNA-binding protein. Nature 2003, 424, 961–965. [Google Scholar] [CrossRef] [PubMed]
- Kuwano, Y.; Gorospe, M. Protecting the stress response, guarding the MKP-1 mRNA. Cell Cycle 2008, 7, 2640–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Rodriguez, E.; Fernandez-Pinar, P.; Sacristan-Reviriego, A.; Molina, M.; Martin, H. An Analog-sensitive Version of the Protein Kinase Slt2 Allows Identification of Novel Targets of the Yeast Cell Wall Integrity Pathway. J. Biol. Chem. 2016, 291, 5461–5472. [Google Scholar] [CrossRef]
- Lin, Y.W.; Chuang, S.M.; Yang, J.L. ERK1/2 achieves sustained activation by stimulating MAPK phosphatase-1 degradation via the ubiquitin-proteasome pathway. J. Biol. Chem. 2003, 278, 21534–21541. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Rubio, G.; Fernández-Acero, T.; Martín, H.; Molina, M. Mitogen-Activated Protein Kinase Phosphatases (MKPs) in Fungal Signaling: Conservation, Function, and Regulation. Int. J. Mol. Sci. 2019, 20, 1709. https://doi.org/10.3390/ijms20071709
González-Rubio G, Fernández-Acero T, Martín H, Molina M. Mitogen-Activated Protein Kinase Phosphatases (MKPs) in Fungal Signaling: Conservation, Function, and Regulation. International Journal of Molecular Sciences. 2019; 20(7):1709. https://doi.org/10.3390/ijms20071709
Chicago/Turabian StyleGonzález-Rubio, Gema, Teresa Fernández-Acero, Humberto Martín, and María Molina. 2019. "Mitogen-Activated Protein Kinase Phosphatases (MKPs) in Fungal Signaling: Conservation, Function, and Regulation" International Journal of Molecular Sciences 20, no. 7: 1709. https://doi.org/10.3390/ijms20071709
APA StyleGonzález-Rubio, G., Fernández-Acero, T., Martín, H., & Molina, M. (2019). Mitogen-Activated Protein Kinase Phosphatases (MKPs) in Fungal Signaling: Conservation, Function, and Regulation. International Journal of Molecular Sciences, 20(7), 1709. https://doi.org/10.3390/ijms20071709