Altered Glutamate Receptor Ionotropic Delta Subunit 2 Expression in Stau2-Deficient Cerebellar Purkinje Cells in the Adult Brain


 and
and 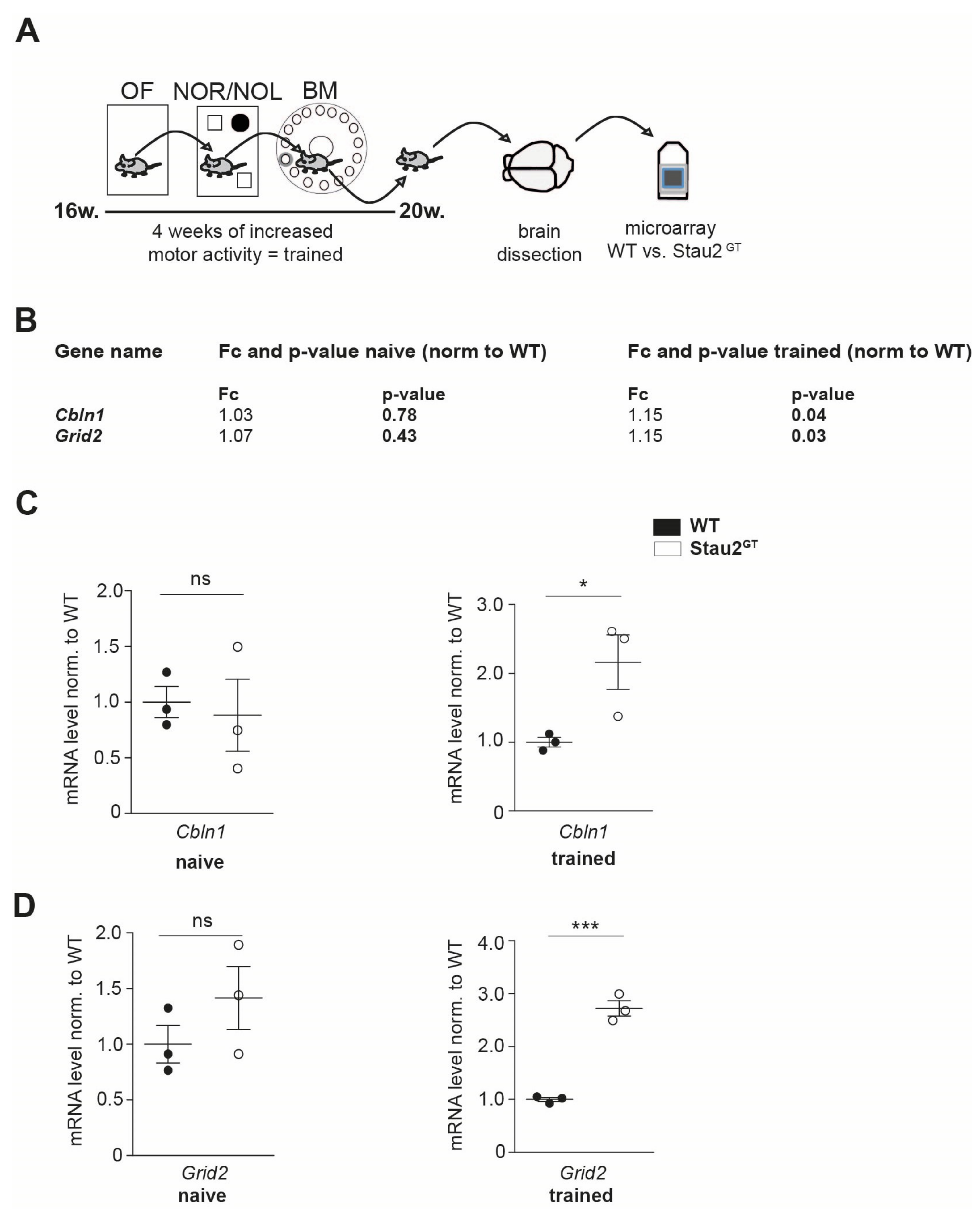{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Adult Stau2GT Mice Show an Upregulation of Cbln1 and GluD2 mRNA during Behavior Testing
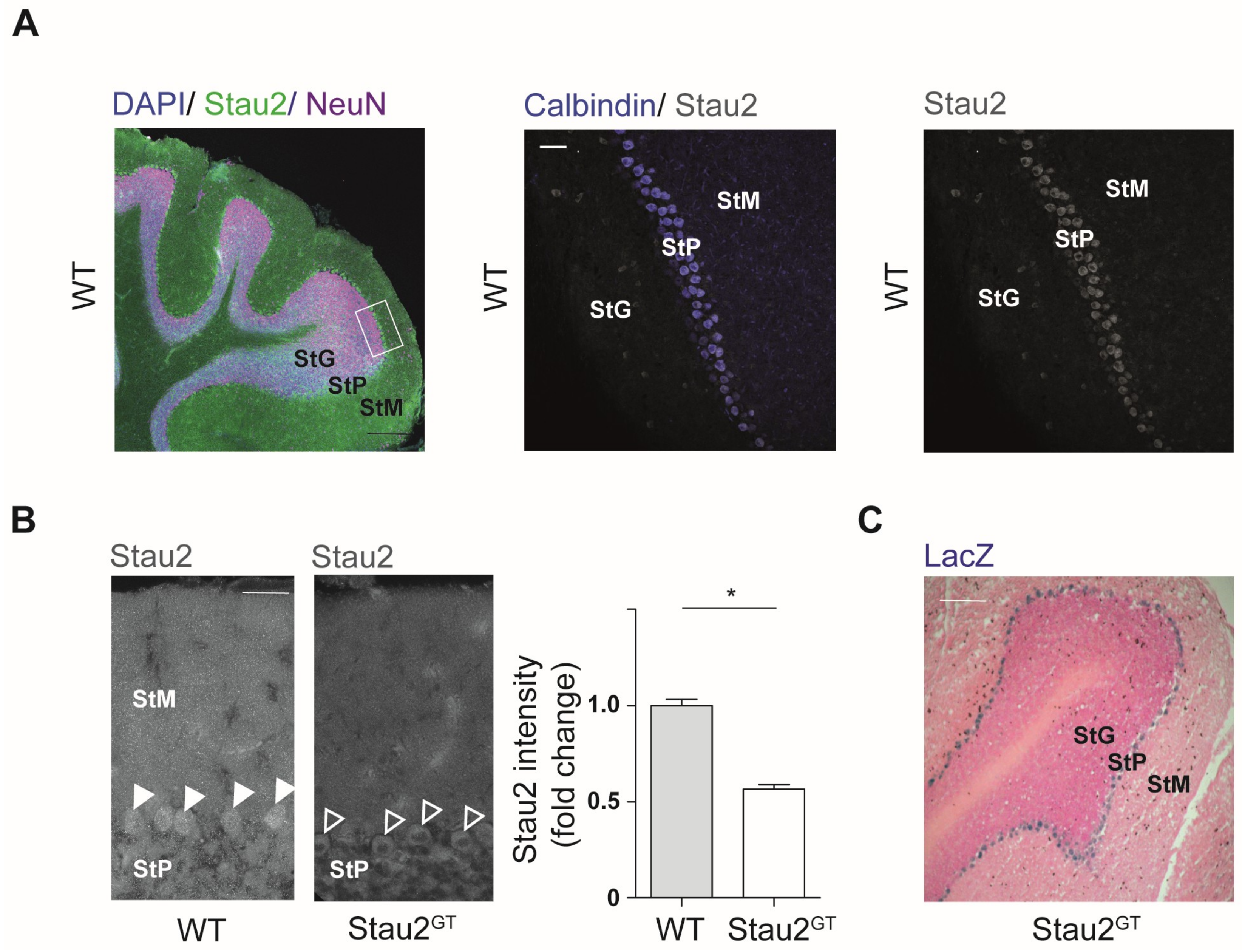
2.2. Reduced Stau2 Expression in Cerebellar Purkinje Cells of Adult Stau2GT Mice
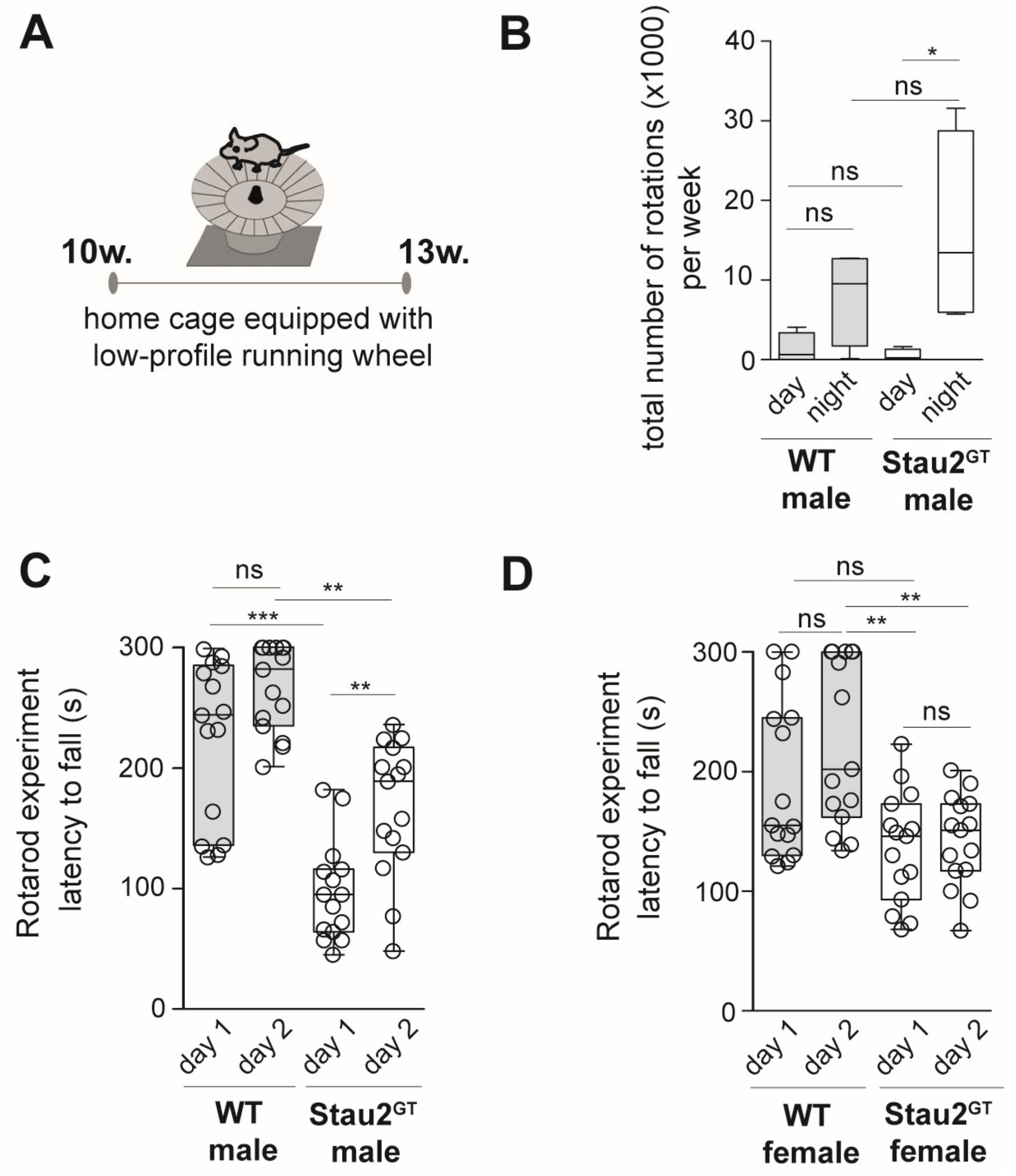
2.3. Decreased Motor Coordination Abilities but Increased Motor Learning Capacity in Adult Male Stau2GT Mice
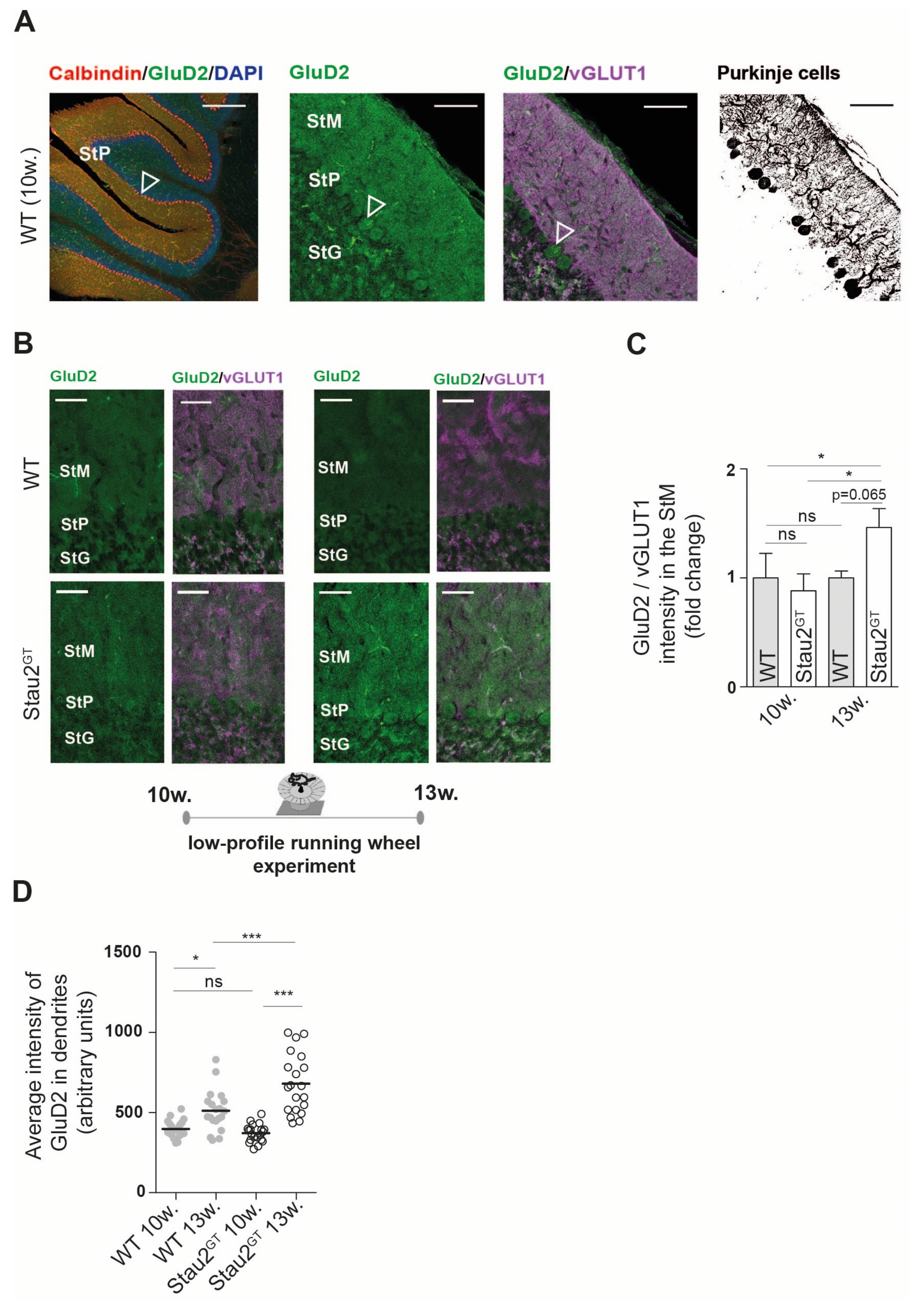
2.4. Increased Dendritic GluD2 Protein Expression in Adult Stau2GT Mice after Motor Activity.
3. Discussion
4. Materials and Methods
4.1. Generation and Housing of Stau2 Gene Trap Mice
4.2. Behavioral Analysis
4.3. Histology
4.4. RT-qPCR
4.5. Expression Analysis
4.6. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Cbln1 GluD2 KO LTD RBP Stau2 Stau2GT StG StM StP vGLUT1 WT | Cerebellin1 Glutamate receptor ionotropic delta subunit 2 Knock out Long-term depression RNA-binding protein Staufen2 Staufen2 gene trap Stratum granulosum Stratum moleculare Stratum pyramidale Vesicular glutamate transporter 1 Wild type |
References
- Jung, H.; Gkogkas, C.G.; Sonenberg, N.; Holt, C.E. Remote control of gene function by local translation. Cell 2014, 157, 26–40. [Google Scholar] [CrossRef]
- Doyle, M.; Kiebler, M.A. Mechanisms of dendritic mRNA transport and its role in synaptic tagging. EMBO J. 2011, 30, 3540–3552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiebler, M.A.; Bassell, G.J. Neuronal RNA Granules: Movers and Makers. Neuron 2006, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Jarero-Basulto, J.J.; Gasca-Martinez, Y.; Rivera-Cervantes, M.; Ureña-Guerrero, M.; Feria-Valesco, A.I.; Beas-Zarate, C. Interactions Between Epilepsy and Plasticity. Pharmaceuticals 2018, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Liu-Yesucevitz, L.; Bassell, G.J.; Gitler, A.D.; Hart, A.C.; Klann, E.; Richter, J.D.; Warren, S.T.; Wolozin, B. Local RNA Translation at the Synapse and in Disease. J. Neurosci. 2011. [Google Scholar] [CrossRef] [PubMed]
- Swanger, S.A.; Bassell, G.J. Dendritic protein synthesis in the normal and diseased brain. Neuroscience 2013, 232, 106–127. [Google Scholar] [CrossRef] [PubMed]
- Hanson, K.A.; Kim, S.H.; Tibbetts, R.S. RNA-binding proteins in neurodegenerative disease: TDP-43 and beyond. Wiley Interdiscip. Rev. RNA 2012, 265–285. [Google Scholar] [CrossRef]
- Heraud-Farlow, J.E.; Kiebler, M.A. The multifunctional Staufen proteins: Conserved roles from neurogenesis to synaptic plasticity. Trends Neurosci. 2014, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Popper, B.; Demleitner, A.; Bolivar, V.J.; Kusek, G.; Snyder-Keller, A.; Schieweck, R.; Temple, S.; Kiebler, M.A. Staufen2 deficiency leads to impaired response to novelty in mice. Neurobiol. Learn. Mem. 2018. [Google Scholar] [CrossRef]
- Berger, S.M.; Fernández-Lamo, I.; Schönig, K.; Fernández Moya, S.M.; Ehses, J.; Schieweck, R.; Clementi, S.; Enkel, T.; Grothe, S.; von Bohlen und Halbach, O.; et al. Forebrain-specific, conditional silencing of Staufen2 alters synaptic plasticity, learning, and memory in rats. Genome Biol. 2017, 18, 222. [Google Scholar] [Green Version]
- Hirano, T. Glutamate-receptor-like molecule GluRδ2 involved in synapse formation at parallel fiber-Purkinje neuron synapses. Cerebellum 2012, 11, 71–77. [Google Scholar] [CrossRef]
- Matsuda, K.; Miura, E.; Miyazaki, T.; Kakegawa, W.; Emi, K.; Narumi, S.; Fukazawa, Y.; Ito-lshida, A.; Kondo, T.; Shigemoto, R.; et al. Cbln1 is a ligand for an orphan glutamate receptor δ2, a bidirectional synapse organizer. Science. 2010. [Google Scholar] [CrossRef]
- Mishina, M.; Uemura, T.; Yasumura, M.; Yoshida, T. Molecular mechanism of parallel fiber-Purkinje cell synapse formation. Front. Neural Circuits 2012, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Seven, A.B.; Wang, J.; Skiniotis, G.; Özkan, E. Conformational Plasticity in the Transsynaptic Neurexin-Cerebellin-Glutamate Receptor Adhesion Complex. Structure 2016, 24, 2163–2173. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, M.; Miyazaki, T.; Sakimura, K.; Watanabe, M.; Kitamura, K.; Kano, M. Disruption of cerebellar microzonal organization in GluD2 (GluRδ2) knockout mouse. Front. Neural Circuits 2013, 7, 130. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, Y.; Yoshioka, Y.; Suzuki, K.; Miyazaki, T.; Koura, M.; Saigoh, K.; Kajimura, N.; Monobe, Y.; Kusunoki, S.; Matsuda, J.; et al. A new mouse allele of glutamate receptor delta 2 with cerebellar atrophy and progressive ataxia. PLoS ONE 2014, 9, e107867. [Google Scholar] [CrossRef] [PubMed]
- Coutelier, M.; Burglen, L.; Mundwiller, E.; Abada-Bendib, M.; Rodriguez, D.; Chantot-Bastaraud, S.; Rougeot, C.; Cournelle, M.A.; Milh, M.; Toutain, A.; et al. GRID2 mutations span from congenital to mild adult-onset cerebellar ataxia. Neurology 2015, 84, 1751–1759. [Google Scholar] [CrossRef]
- Miura, E.; Matsuda, K.; Morgan, J.I.; Yuzaki, M.; Watanabe, M. Cbln1 accumulates and colocalizes with Cbln3 and GluRδ2 at parallel fiber-Purkinje cell synapses in the mouse cerebellum. Eur. J. Neurosci. 2009, 29, 693–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, M.; Mason, G. A comparison of two types of running wheel in terms of mouse preference, health, and welfare. Physiol. Behav. 2018. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.P.; Trueman, R.C.; Dunnett, S.B. Assessment of Motor Coordination and Balance in Mice Using the Rotarod, Elevated Bridge, and Footprint Tests. Curr. Protoc. Mouse Biol. 2012, 2, 37–53. [Google Scholar]
- Aguiar, A.S.; Castro, A.A.; Moreira, E.L.; Glaser, V.; Santos, A.R.S.; Tasca, C.I.; Latini, A.; Prediger, R.D.S. Short bouts of mild-intensity physical exercise improve spatial learning and memory in aging rats: Involvement of hippocampal plasticity via AKT, CREB and BDNF signaling. Mech. Ageing Dev. 2011, 132, 560–567. [Google Scholar] [CrossRef]
- Fremeau, R.T.; Troyer, M.D.; Pahner, I.; Nygaard, G.O.; Tran, C.H.; Reimer, R.J.; Bellocchio, E.E.; Fortin, D.; Storm-Mathisen, J.; Edwards, R.H. The expression of vesicular glutamate transporters defines two classes of excitatory synapse. Neuron 2001, 31, 247–260. [Google Scholar] [CrossRef]
- Goetze, B.; Tuebing, F.; Xie, Y.; Dorostkar, M.M.; Thomas, S.; Pehl, U.; Boehm, S.; Macchi, P.; Kiebler, M.A. The brain-specific double-stranded RNA-binding protein Staufen2 is required for dendritic spine morphogenesis. J. Cell Biol. 2006, 172, 221–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharangdhar, T.; Sugimoto, Y.; Heraud-Farlow, J.; Fernández-Moya, S.M.; Ehses, J.; Ruiz de los Mozos, I.; Ule, J.; Kiebler, M.A. A retained intron in the 3′-UTR of Calm3 mRNA mediates its Staufen2- and activity-dependent localization to neuronal dendrites. EMBO Rep. 2017, 18, 1762–1774. [Google Scholar] [CrossRef] [PubMed]
- Uemura, T.; Lee, S.J.; Yasumura, M.; Takeuchi, T.; Yoshida, T.; Ra, M.; Taguchi, R.; Sakimura, K.; Mishina, M. Trans-synaptic interaction of GluRδ2 and neurexin through Cbln1 mediates synapse formation in the cerebellum. Cell 2010, 18, 1762–1774. [Google Scholar] [CrossRef]
- Zanjani, H.S.; Vogel, M.W.; Mariani, J. Deletion of the GluRδ2 Receptor in the Hotfoot Mouse Mutant Causes Granule Cell Loss, Delayed Purkinje Cell Death, and Reductions in Purkinje Cell Dendritic Tree Area. Cerebellum 2016, 15, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Kashiwabuchi, N.; Ikeda, K.; Araki, K.; Hirano, T.; Shibuki, K.; Takayama, C.; Inoue, Y.; Kutsuwada, T.; Yagi, T.; Kang, Y.; et al. Impairment of motor coordination, Purkinje cell synapse formation, and cerebellar long-term depression in GluR delta 2 mutant mice. Cell 1995, 81, 245–252. [Google Scholar] [CrossRef]
- Kakegawa, W.; Miyoshi, Y.; Hamase, K.; Matsuda, S.; Matsuda, K.; Kohda, K.; Emi, K.; Motohashi, J.; Konno, R.; Zaitsu, K.; et al. D-Serine regulates cerebellar LTD and motor coordination through the δ2 glutamate receptor. Nat. Neurosci. 2011, 14, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Hirai, H.; Launey, T.; Mikawa, S.; Torashima, T.; Yanagihara, D.; Kasaura, T.; Miyamoto, A.; Yuzaki, M. New role of delta2-glutamate receptors in AMPA receptor trafficking and cerebellar function. Nat. Neurosci. 2003, 6, 869–876. [Google Scholar] [CrossRef]
- Uemura, T.; Mori, H.; Mishina, M. Direct interaction of GluRdelta2 with Shank scaffold proteins in cerebellar Purkinje cells. Mol. Cell. Neurosci. 2004. [CrossRef]
- Lebeau, G.; Miller, L.C.; Tartas, M.; McAdam, R.; Laplante, I.; Badeaux, F.; DesGroseillers, L.; Sossin, W.S.; Lacaille, J.C. Staufen 2 regulates mGluR long-term depression and Map1b mRNA distribution in hippocampal neurons. Learn. Mem. 2011, 18, 314–326. [Google Scholar] [CrossRef] [Green Version]
- Hirai, H.; Pang, Z.; Bao, D.; Miyazaki, T.; Li, L.; Miura, E.; Parris, J.; Rong, Y.; Watanabe, M.; Yuzaki, M.; et al. Cbln1 is essential for synaptic integrity and plasticity in the cerebellum. Nat. Neurosci. 2005. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Yuzaki, M. Cbln1 and the delta2 glutamate receptor—An orphan ligand and an orphan receptor find their partners. Cerebellum 2012, 11, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, S.; Konno, K.; Abe, M.; Motohashi, J.; Kohda, K.; Sakimura, K.; Watanabe, M.; Yuzaki, M. Roles of Cbln1 in Non-Motor Functions of Mice. J. Neurosci. 2016, 36, 11801–11816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, E.; Ito-Ishida, A.; Yuzaki, M.; Yanagihara, D. Improvement of cerebellar ataxic gait by injecting Cbln1 into the cerebellum of cbln1-null mice. Sci. Rep. 2018, 8, 6184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamm, R.J.; Pike, B.R.; O’Dell, D.M.; Lyeth, B.G.; Jenkins, L.W. The rotarod test: An evaluation of its effectiveness in assessing motor deficits following traumatic brain injury. J. Neurotrauma 1994, 11, 187–196. [Google Scholar] [CrossRef]
- Calzolari, F.; Michel, J.; Baumgart, E.V.; Theis, F.; Götz, M.; Ninkovic, J. Fast clonal expansion and limited neural stem cell self-renewal in the adult subependymal zone. Nat. Neurosci. 2015, 18, 490–492. [Google Scholar] [CrossRef] [PubMed]
- Follwaczny, P.; Schieweck, R.; Riedemann, T.; Demleitner, A.; Straub, T.; Klemm, A.H.; Bilban, M.; Sutor, B.; Popper, B.; Kiebler, M.A. Pumilio2-deficient mice show a predisposition for epilepsy. Dis. Model. Mech. 2017, 10, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Heraud-Farlow, J.E.; Sharangdhar, T.; Li, X.; Pfeifer, P.; Tauber, S.; Orozco, D.; Hörmann, A.; Thomas, S.; Bakosova, A.; Farlow, A.R.; et al. Staufen2 regulates neuronal target RNAs. Cell Rep. 2013, 5, 1511–1518. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pernice, H.F.; Schieweck, R.; Jafari, M.; Straub, T.; Bilban, M.; Kiebler, M.A.; Popper, B. Altered Glutamate Receptor Ionotropic Delta Subunit 2 Expression in Stau2-Deficient Cerebellar Purkinje Cells in the Adult Brain. Int. J. Mol. Sci. 2019, 20, 1797. https://doi.org/10.3390/ijms20071797
Pernice HF, Schieweck R, Jafari M, Straub T, Bilban M, Kiebler MA, Popper B. Altered Glutamate Receptor Ionotropic Delta Subunit 2 Expression in Stau2-Deficient Cerebellar Purkinje Cells in the Adult Brain. International Journal of Molecular Sciences. 2019; 20(7):1797. https://doi.org/10.3390/ijms20071797
Chicago/Turabian StylePernice, Helena F., Rico Schieweck, Mehrnoosh Jafari, Tobias Straub, Martin Bilban, Michael A. Kiebler, and Bastian Popper. 2019. "Altered Glutamate Receptor Ionotropic Delta Subunit 2 Expression in Stau2-Deficient Cerebellar Purkinje Cells in the Adult Brain" International Journal of Molecular Sciences 20, no. 7: 1797. https://doi.org/10.3390/ijms20071797
APA StylePernice, H. F., Schieweck, R., Jafari, M., Straub, T., Bilban, M., Kiebler, M. A., & Popper, B. (2019). Altered Glutamate Receptor Ionotropic Delta Subunit 2 Expression in Stau2-Deficient Cerebellar Purkinje Cells in the Adult Brain. International Journal of Molecular Sciences, 20(7), 1797. https://doi.org/10.3390/ijms20071797