Intermittent Hypoxia Up-Regulates Gene Expressions of Peptide YY (PYY), Glucagon-like Peptide-1 (GLP-1), and Neurotensin (NTS) in Enteroendocrine Cells

 , and
, and 
Abstract
:
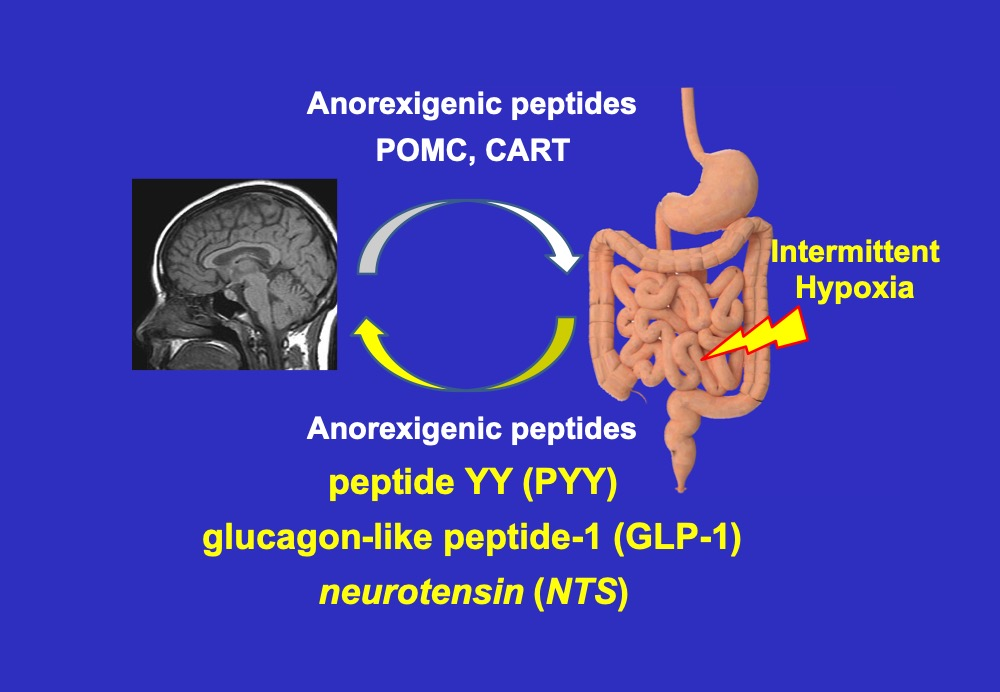
1. Introduction
2. Results
2.1. IH Significantly Increased the mRNA Levels of PYY, GLP-1, and NTS in Enteroendocrine Cells
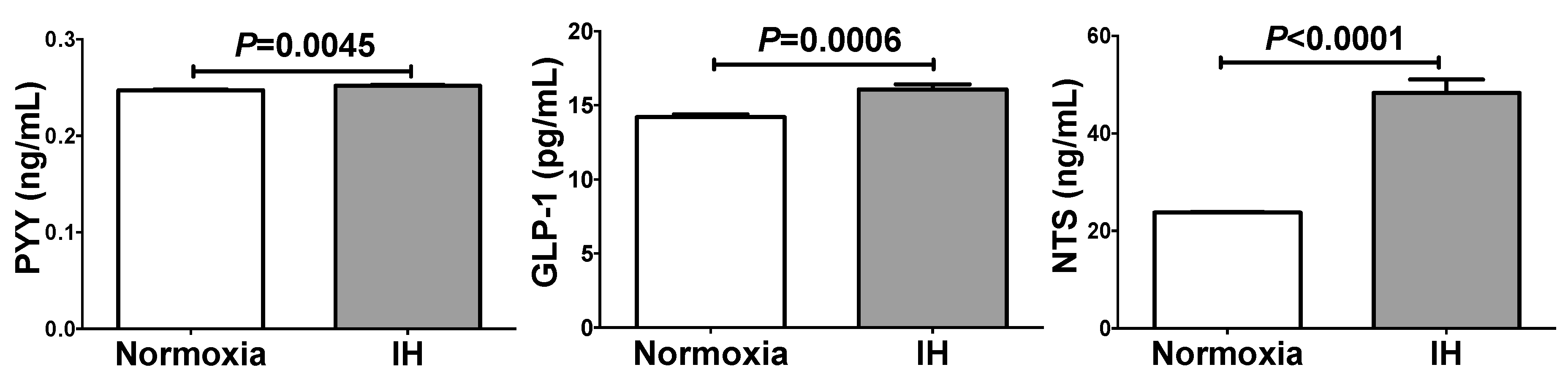
2.2. IH Significantly Increased Secretion of PYY, GLP-1, and NTS from Caco-2 Cells
2.3. The Promoter Activities of PYY, Glucagon (GCG), and NTS Were Not Up-Regulated by IH in Caco-2 Cells
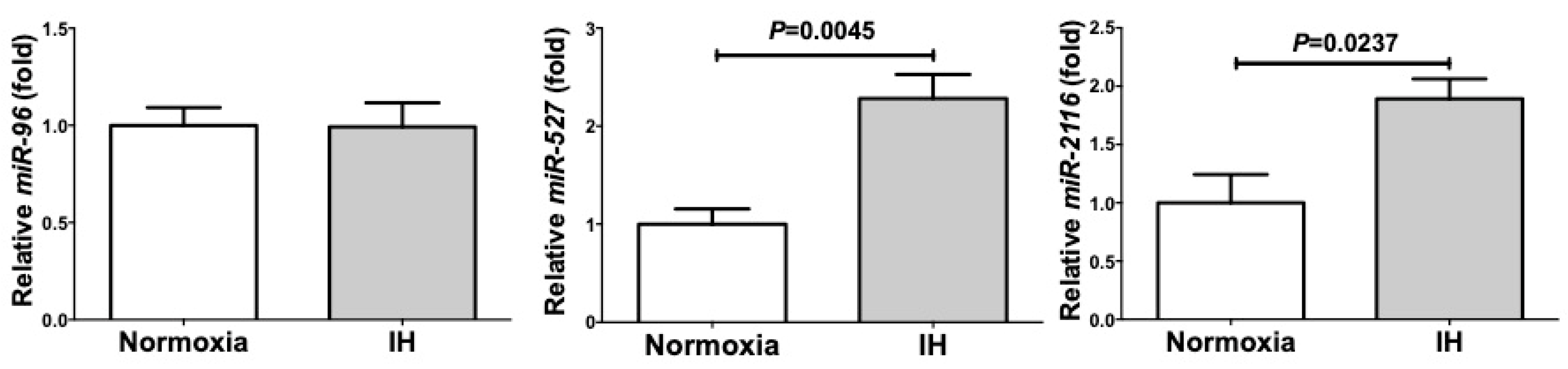
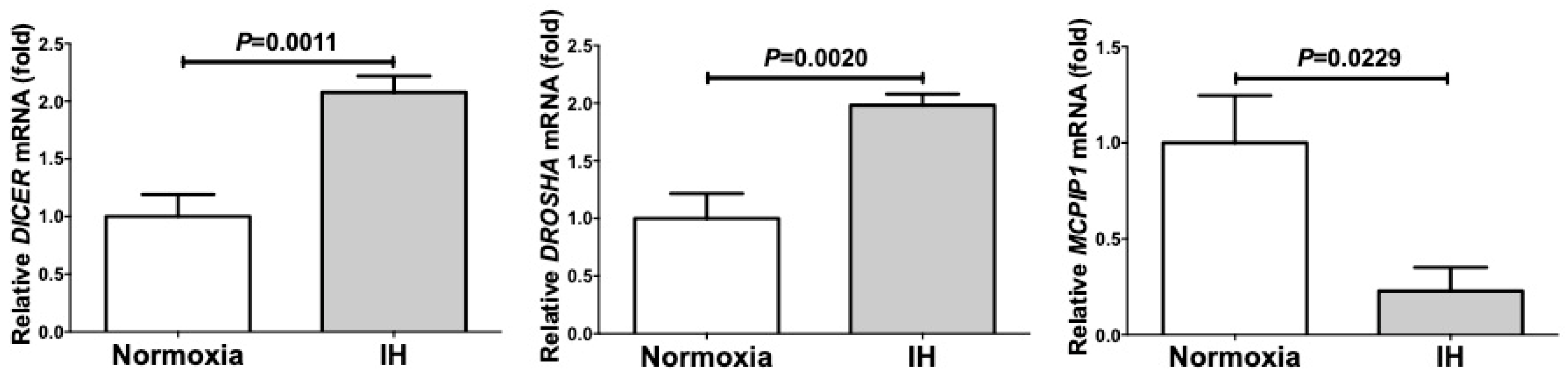
2.4. MicroRNA-Mediated Mechanism Was Not Involved in the IH-Induced Up-Regulation of PYY, GLP-1, and NTS
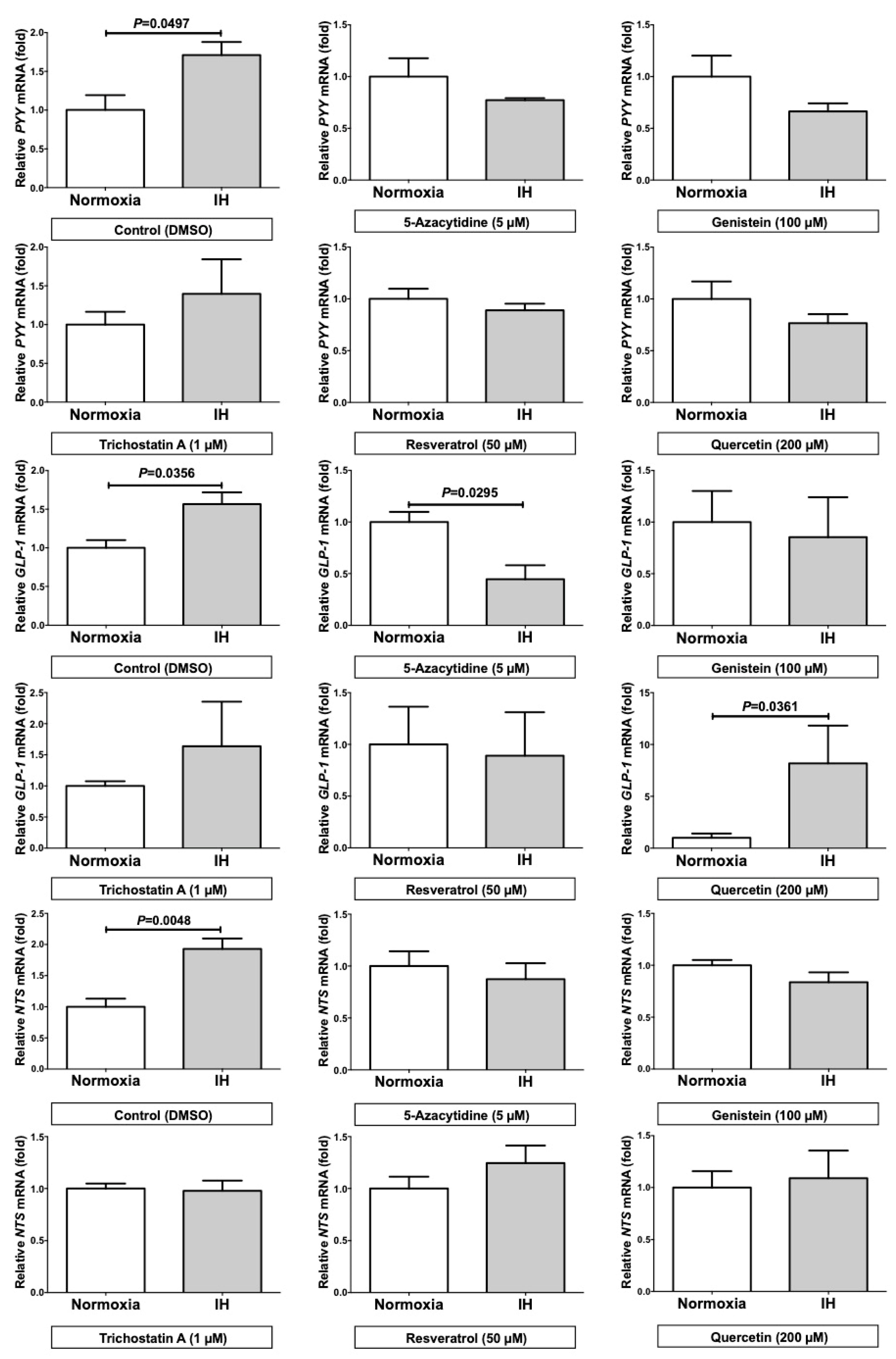
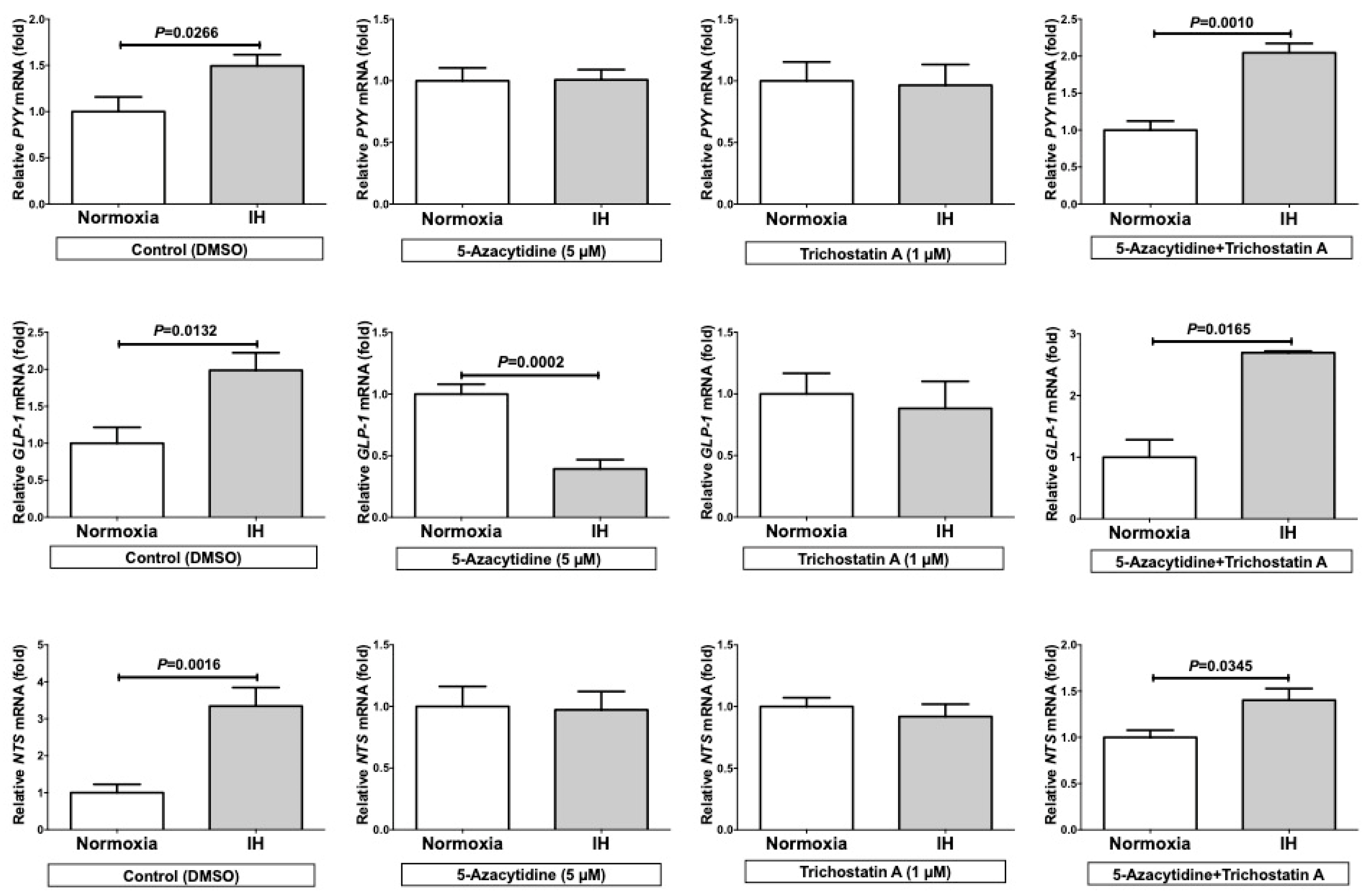
2.5. Trichostatin A (TSA) Significantly Up-regulated the mRNA Levels of PYY, GLP-1, and NTS in the Normoxia Condition and 5-Azacytidine (5AZC) Significantly Decreased the mRNA Levels of PYY, GLP-1, and NTS in the IH Condition
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.3. Measurement of PYY, GLP-1, and NTS in Culture Medium by Enzyme-Linked Immunosorbent Assay (ELISA)
4.4. Construction of Reporter Plasmids and Luciferase Reporter Assay
4.5. Treatment of Caco-2 cells with 5AZC, Genistein, TSA, Resveratrol and Quercetin
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 5AZC | 5-Azacytidine |
| AGRP | Agouti-related peptide |
| CART | Cocaine- and amphetamine-regulated transcript |
| CNS | Central nervous system |
| DICER | Endoribonuclease Dicer |
| DROSHA | Ribonuclease type III |
| ENS | Enteric nervous system |
| GAL | Galanin |
| GALP | Galanin-like peptide |
| GHRL | Ghrelin |
| GCG | Glucagon |
| GI | Gastrointestinal |
| GLP-1 | Glucagon-like peptide-1 |
| IH | Intermittent hypoxia |
| MC4R | Melanocortin 4 receptor |
| MCPIP1 | Monocyte chemotactic protein-induced protein 1 |
| NPY | Neuropeptide Y |
| NTS | Neurotensin |
| POMC | Proopiomelanocortin |
| PYY | Peptide YY |
| QRFP | Pyroglutamylated RFamide peptide |
| RT-PCR | Reverse transcription-polymerase chain reaction |
| SAS | Sleep apnea syndrome |
| TSA | Trichostatin A |
References
- Dempsey, J.A.; Veasey, S.C.; Morgan, B.J.; O’Donnell, C.P. Pathophysiology of sleep apnea. Physiol. Rev. 2010, 90, 47–112. [Google Scholar] [CrossRef]
- Ota, H.; Takasawa, S.; Yamauchi, M.; Yoshikawa, M.; Tomoda, K.; Kimura, H. Intermittent hypoxia in pancreatic beta cells. Pancreat. Disord. Ther. 2015, 5, S5-004. [Google Scholar]
- Romero-Corral, A.; Caples, S.M.; Lopez-Jimenez, F.; Somers, V.K. Interactions between obesity and obstructive sleep apnea: Implications for treatment. Chest 2010, 137, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Shobatake, R.; Takasawa, K.; Ota, H.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Uchiyama, T.; Makino, M.; Sugie, K.; Takasawa, S.; et al. Up-regulation of POMC and CART mRNAs by intermittent hypoxia via GATA transcription factors in human neuronal cells. Int. J. Biochem. Cell Biol. 2018, 95, 100–107. [Google Scholar] [CrossRef]
- Bauer, P.V.; Hamr, S.C.; Duca, F.A. Regulation of energy balance by a gut-brain axis and involvement of the gut microbiota. Cell Mol. Life Sci. 2016, 73, 737–755. [Google Scholar] [CrossRef] [PubMed]
- Gu, N.; Adachi, T.; Matsunaga, T.; Takeda, J.; Tsujimoto, G.; Ishihara, A.; Yasuda, K.; Tsuda, K. Mutant HNF-1α and mutant HNF-1β identified in MODY3 and MODY5 downregulate DPP-IV gene expression in Caco-2 cells. Biochem. Biophys. Res. Commun. 2006, 346, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Beau, I.; Berger, A.; Servin, A.L. Rotavirus impairs the biosynthesis of brush-border-associated dipeptidyl peptidase IV in human enterocyte-like Caco-2/TC7 cells. Cell. Microbiol. 2007, 9, 779–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drucker, D.J. The biology of incretin hormones. Cell Metab. 2006, 3, 153–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocher, B.; Reichetzeder, C.; Alter, M.L. Renal and cardiac effects of DPP-4 inhibitors—From preclinical development to clinical research. Kidney Blood Press. Res. 2012, 36, 65–84. [Google Scholar] [CrossRef] [PubMed]
- Young, T.; Peppard, P.E.; Taheri, S. Excess weight and sleep-disordered breathing. J. Appl. Physiol. 2005, 99, 1592–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.K.; Kushida, C.; Powell, N.B.; Riley, R.W.; Guilleminault, C. Obstructive sleep apnea syndrome: A comparison between Far-East Asian and white men. Laryngoscope 2000, 110, 1689–1693. [Google Scholar] [CrossRef]
- Batterham, R.L.; Heffron, H.; Kapoor, S.; Chivers, J.E.; Chandarana, K.; Herzog, H.; Le Roux, C.W.; Thomas, E.L.; Bell, J.D.; Withers, D.J. Critical role of peptide YY in protein-mediated satiation and body-weight regulation. Cell Metab. 2006, 4, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Grunddal, K.V.; Ratner, C.F.; Svendsen, B.; Sommer, F.; Engelstoft, M.S.; Madsen, A.N.; Pedersen, J.; Nøhr, M.K.; Egerod, K.L.; Nawrocki, A.R.; et al. Neurotensin is coexpressed, coreleased, and acts together with GLP-1 and PYY in enteroendocrine control of metabolism. Endocrinology 2016, 157, 176–194. [Google Scholar] [CrossRef]
- Christman, J.K. 5-Azacytidine and 5-aza-2′-deoxycytidine as inhibitors of DNA methylation: Mechanistic studies and their implications for cancer therapy. Oncogene 2002, 21, 5483–5495. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Chen, D.; Yang, C.S. Dietary polyphenols may affect DNA methylation. J. Nutr. 2007, 137, 223S–228S. [Google Scholar] [CrossRef] [PubMed]
- Vanhaecke, T.; Papeleu, P.; Elaut, G.; Rogiers, V. Trichostatin A-like hydroxamate histone deacetylase inhibitors as therapeutic agents: Toxicological point of view. Curr. Med. Chem. 2004, 11, 1629–1643. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, G.F.S.; Silva, G.D.B.; Pavan, A.R.; Chiba, D.E.; Chin, C.M.; Dos Santos, J.L. Epigenetic regulatory mechanisms induced by resveratrol. Nutrients 2017, 9, 1201. [Google Scholar] [CrossRef] [PubMed]
- Mikami, S.; Ota, I.; Masui, T.; Itaya-Hironaka, A.; Shobatake, R.; Okamoto, H.; Takasawa, S.; Kitahara, T. Effect of resveratrol on cancer progression through the REG III expression pathway in head and neck cancer cells. Int. J. Oncol. 2016, 49, 1553–1560. [Google Scholar] [CrossRef]
- Busch, C.; Burkard, M.; Leischner, C.; Lauer, U.M.; Frank, J.; Venturelli, S. Epigenetic activities of flavonoids in the prevention and treatment of cancer. Clin. Epigenetics 2015, 7, 64. [Google Scholar] [CrossRef]
- Ota, H.; Tamaki, S.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Morioka, T.; Takasawa, S.; Kimura, H. Attenuation of glucose-induced insulin secretion by intermittent hypoxia via down-regulation of CD38. Life Sci. 2012, 90, 206–211. [Google Scholar] [CrossRef]
- Ota, H.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Miyaoka, T.; Fujimura, T.; Tsujinaka, H.; Yoshimoto, K.; Nakagawara, K.; Tamaki, S.; et al. Pancreatic β cell proliferation by intermittent hypoxia via up-regulation of Reg family genes and HGF gene. Life Sci. 2013, 93, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, K.; Fujimoto, T.; Itaya-Hironaka, A.; Miyaoka, T.; Sakuramoto-Tsuchida, S.; Yamauchi, A.; Takeda, M.; Kasai, T.; Nakagawara, K.; Nonomura, A.; et al. Involvement of autoimmunity to REG, a regeneration factor, in patients with primary Sjögren’s syndrome. Clin. Exp. Immunol. 2013, 174, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, T.; Fujimoto, T.; Itaya-Hironaka, A.; Miyaoka, T.; Yoshimoto, K.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Kondo, S.; Takeda, M.; Tsujinaka, H.; et al. Interleukin-6/STAT pathway is responsible for the induction of gene expression of REG Iα, a new auto-antigen in Sjögren’s syndrome patients, in salivary duct epithelial cells. Biochem. Biophys. Rep. 2015, 2, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Tsujinaka, H.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Ota, H.; Takeda, M.; Fujimura, T.; Takasawa, S.; Ogata, N. Human retinal pigment epithelial cell proliferation by the combined stimulation of hydroquinone and advanced glycation end-products via up-regulation of VEGF gene. Biochem. Biophys. Rep. 2015, 2, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, T.; Ota, H.; Itaya-Hironaka, A.; Shobatake, R.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Makino, M.; Kimura, H.; Takeda, M.; Ohbayashi, C.; et al. Up-regulation of selenoprotein P and HIP/PAP mRNAs in hepatocytes by intermittent hypoxia via down-regulation of miR-203. Biochem. Biophys. Rep. 2017, 11, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Tohma, Y.; Dohi, Y.; Shobatake, R.; Uchiyama, T.; Takeda, M.; Takasawa, S.; Tanaka, Y.; Ohgushi, H. Reg gene expression in periosteum after fracture and its in vitro induction triggered by IL-6. Int. J. Mol. Sci. 2017, 18, 2257. [Google Scholar] [CrossRef] [PubMed]
- Kyotani, Y.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Makino, M.; Takasawa, S.; Yoshizumi, M. Intermittent hypoxia-induced epiregulin expression by IL-6 production in human coronary artery smooth muscle cells. FEBS Open Bio 2018, 8, 868–876. [Google Scholar] [CrossRef]
- Tsuchida, C.; Sakuramoto-Tsuchida, S.; Takeda, M.; Itaya-Hironaka, A.; Yamauchi, A.; Misu, M.; Shobatake, R.; Uchiyama, T.; Makino, M.; Pujol-Autonell, I.; et al. Expression of REG family genes in human inflammatory bowel diseases and its regulation. Biochem. Biophys. Rep. 2017, 12, 198–205. [Google Scholar] [CrossRef]
- Takasawa, S.; Tsuchida, C.; Sakuramoto-Tsuchida, S.; Takeda, M.; Itaya-Hironaka, A.; Yamauchi, A.; Misu, M.; Shobatake, R.; Uchiyama, T.; Makino, M.; et al. Expression of human REG family genes in inflammatory bowel diseases and their molecular mechanism. Immunol. Res. 2018, 66, 800–805. [Google Scholar] [CrossRef]
- Nakagawa, K.; Takasawa, S.; Nata, K.; Yamauchi, A.; Itaya-Hironaka, A.; Ota, H.; Yoshimoto, K.; Sakuramoto-Tsuchida, S.; Miyaoka, T.; Takeda, M.; et al. Prevention of Reg I-induced β-cell apoptosis by IL-6/dexamethasone through activation of HGF gene regulation. Biochim. Biophys. Acta 2013, 1833, 2988–2995. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target mRNA | Primer Sequence (Accession Number: Position) |
|---|---|
| hPYY | 5′-TATGGTGTTCGTGCGCAGGC-3′ (NM_004160: 541-560) |
| 5′-TCACCACAGGTCTGGGCCCTC-3′ (NM_004160: 815-835) | |
| mPyy | 5′-GCAGCGGTATGGAAAAAGAGA-3′ (NM_145435: 219-239) |
| 5′-TCCAAACCTTCTGGCCTGAA- 3′ (NM_145435: 304-323) | |
| hGLP-1 | 5′-TGTCAGCGTAATATCTGTGAGGC-3′ (NM_002054: 86-108) |
| 5′-AGCAGGTGAAGAGAGAGCAAGC-3′ (NM_002054: 201-222) | |
| mGlp-1 | 5′-GCACATTCACCAGCGACTACA-3′ (NM_008100: 267-287) |
| 5′-GCAATGTTGTTCCGGTTCCT-3′ (NM_008100: 347-366) | |
| hNTS | 5′-CAGCAGGGCTTTTCAACACTGGG-3′ (NM_006183: 445-467) |
| 5′-TCTCTTTTGAGTATGTAGGGTCTTCTGGG-3′ (NM_006183: 578-606) | |
| mNts | 5′-CTGGTGTGCCTGACTCTCCT-3′ (NM_024435: 117-136) |
| 5′-TCACATCTTCTTCTGAATCTGAGC-3′ (NM_024435: 160-183) | |
| DROSHA | 5′-GGCCCGAGAGCCTTTTATAG-3′ (NM_013235: 167-186) |
| 5′-TGCACACGTCTAACTCTTCCAC-3′ (NM_013235: 263-284) | |
| DICER | 5′-GAGCTGTCCTATCAGATCAGGG-3′ (NM_177438: 476-497) |
| 5′-ACTTGTTGAGCAACCTGGTTT-3′ (NM_177438: 547-567) | |
| MCPIP1 | 5′-TGCCTATCACAGACCAGCAC-3′ (NM_025079: 722-741) |
| 5′-CTCACCTTCGCGAAGTAGCTC-3′ (NM_025079: 913-933) | |
| miR-96 | 5′-TGGCCGATTTTGGCACTAGCAC-3′ (NR_029512: 1-22) |
| 5′-TTCCCATATTGGCACTGCACATGA-3′ (NR_029512: 54-77) | |
| miR-527 | 5′-CTCAAGCTGTGACTGCAAA-3′ (NR_030219: 2-20) |
| 5′-TCTCAAACCGTAATTCACCAAA-3′ (NR_030219: 64-85) | |
| miR-2116 | 5′-CTAGGGGTTCTTAGCATAGG-3′ (NR_031750: 9-28) |
| 5′-CTAGTCTGGGAGTTCTTGG-3′ (NR_031750: 60-78) | |
| β-actin | 5′-GCGAGAAGATGACCCAGA-3′ (NM_001101: 420-437) |
| 5′-CAGAGGCGTACAGGGATA-3′ (NM_001101: 492-509) | |
| mRig/RpS15 | 5′-ACGGCAAGACCTTCAACCAG-3′ (NM_009091: 323-342) |
| 5′-ATGGAGAACTCGCCCAGGTAG-3′ (NM_009091: 372-392) | |
| U6 | 5′-CTCGCTTCGGCAGCACA-3′ (NR_004394) |
| 5′-AACGCTTCACGAATTTGCGT-3′ (NR_004394) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shobatake, R.; Itaya-Hironaka, A.; Yamauchi, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Uchiyama, T.; Ota, H.; Takahashi, N.; Ueno, S.; Sugie, K.; et al. Intermittent Hypoxia Up-Regulates Gene Expressions of Peptide YY (PYY), Glucagon-like Peptide-1 (GLP-1), and Neurotensin (NTS) in Enteroendocrine Cells. Int. J. Mol. Sci. 2019, 20, 1849. https://doi.org/10.3390/ijms20081849
Shobatake R, Itaya-Hironaka A, Yamauchi A, Makino M, Sakuramoto-Tsuchida S, Uchiyama T, Ota H, Takahashi N, Ueno S, Sugie K, et al. Intermittent Hypoxia Up-Regulates Gene Expressions of Peptide YY (PYY), Glucagon-like Peptide-1 (GLP-1), and Neurotensin (NTS) in Enteroendocrine Cells. International Journal of Molecular Sciences. 2019; 20(8):1849. https://doi.org/10.3390/ijms20081849
Chicago/Turabian StyleShobatake, Ryogo, Asako Itaya-Hironaka, Akiyo Yamauchi, Mai Makino, Sumiyo Sakuramoto-Tsuchida, Tomoko Uchiyama, Hiroyo Ota, Nobuyuki Takahashi, Satoshi Ueno, Kazuma Sugie, and et al. 2019. "Intermittent Hypoxia Up-Regulates Gene Expressions of Peptide YY (PYY), Glucagon-like Peptide-1 (GLP-1), and Neurotensin (NTS) in Enteroendocrine Cells" International Journal of Molecular Sciences 20, no. 8: 1849. https://doi.org/10.3390/ijms20081849
APA StyleShobatake, R., Itaya-Hironaka, A., Yamauchi, A., Makino, M., Sakuramoto-Tsuchida, S., Uchiyama, T., Ota, H., Takahashi, N., Ueno, S., Sugie, K., & Takasawa, S. (2019). Intermittent Hypoxia Up-Regulates Gene Expressions of Peptide YY (PYY), Glucagon-like Peptide-1 (GLP-1), and Neurotensin (NTS) in Enteroendocrine Cells. International Journal of Molecular Sciences, 20(8), 1849. https://doi.org/10.3390/ijms20081849