Citrus Auraptene Induces Glial Cell Line-Derived Neurotrophic Factor in C6 Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
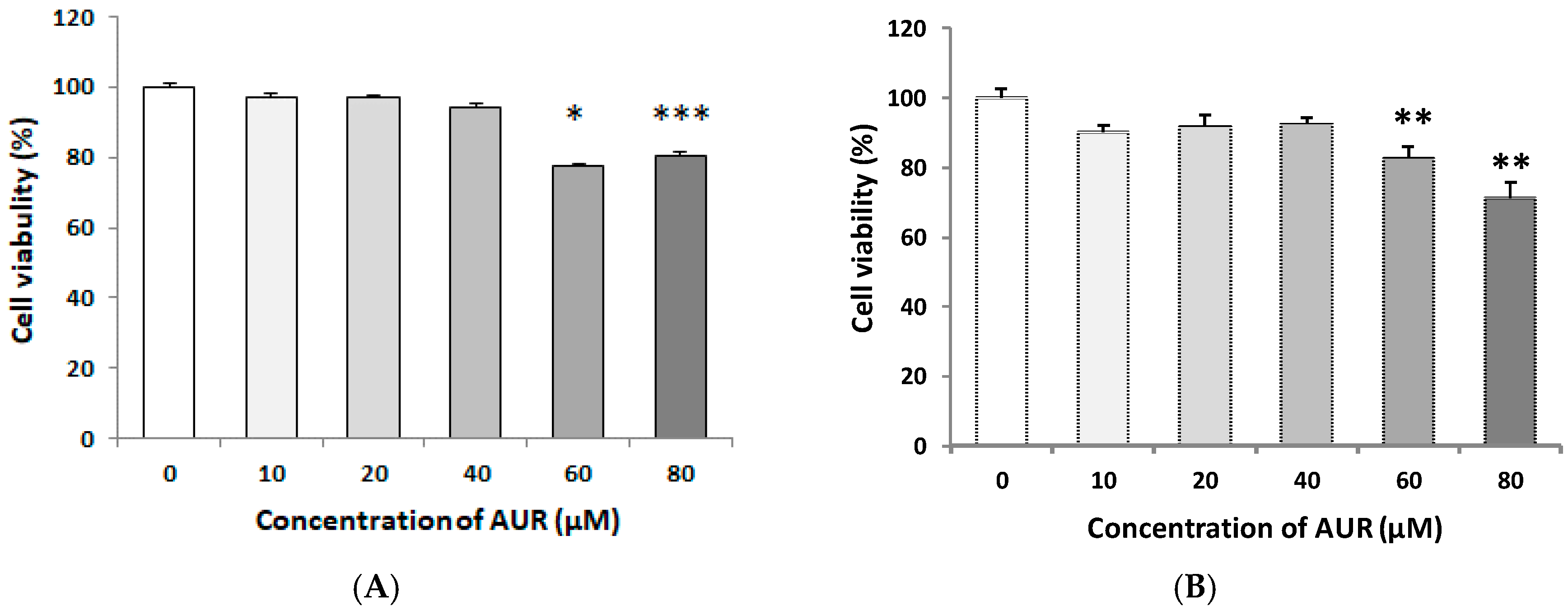
2.1. Effects of AUR on the Viability of C6 Cells
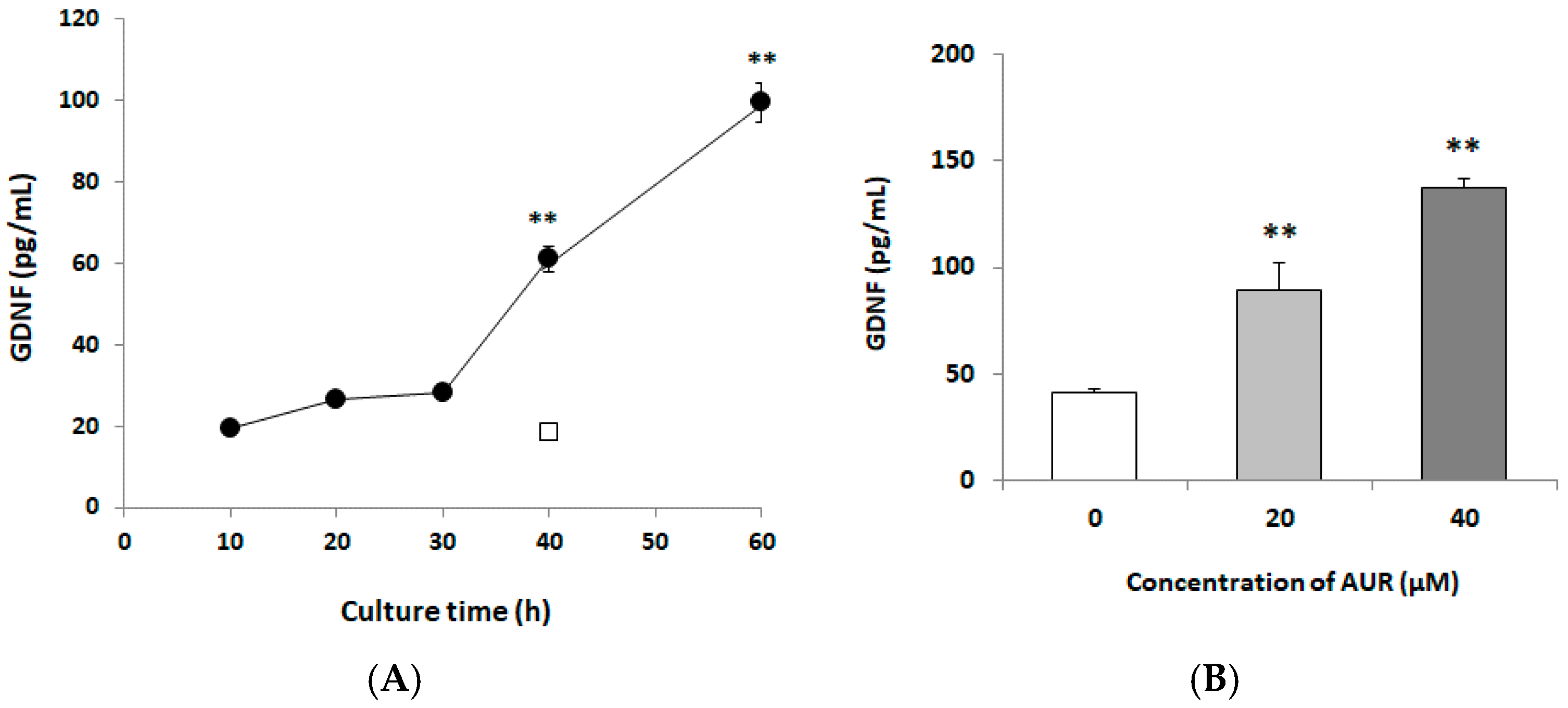
2.2. Effects of AUR on GDNF Content of Conditioned Media
2.3. Effects of AUR on GDNF Levels in Cell Lysates
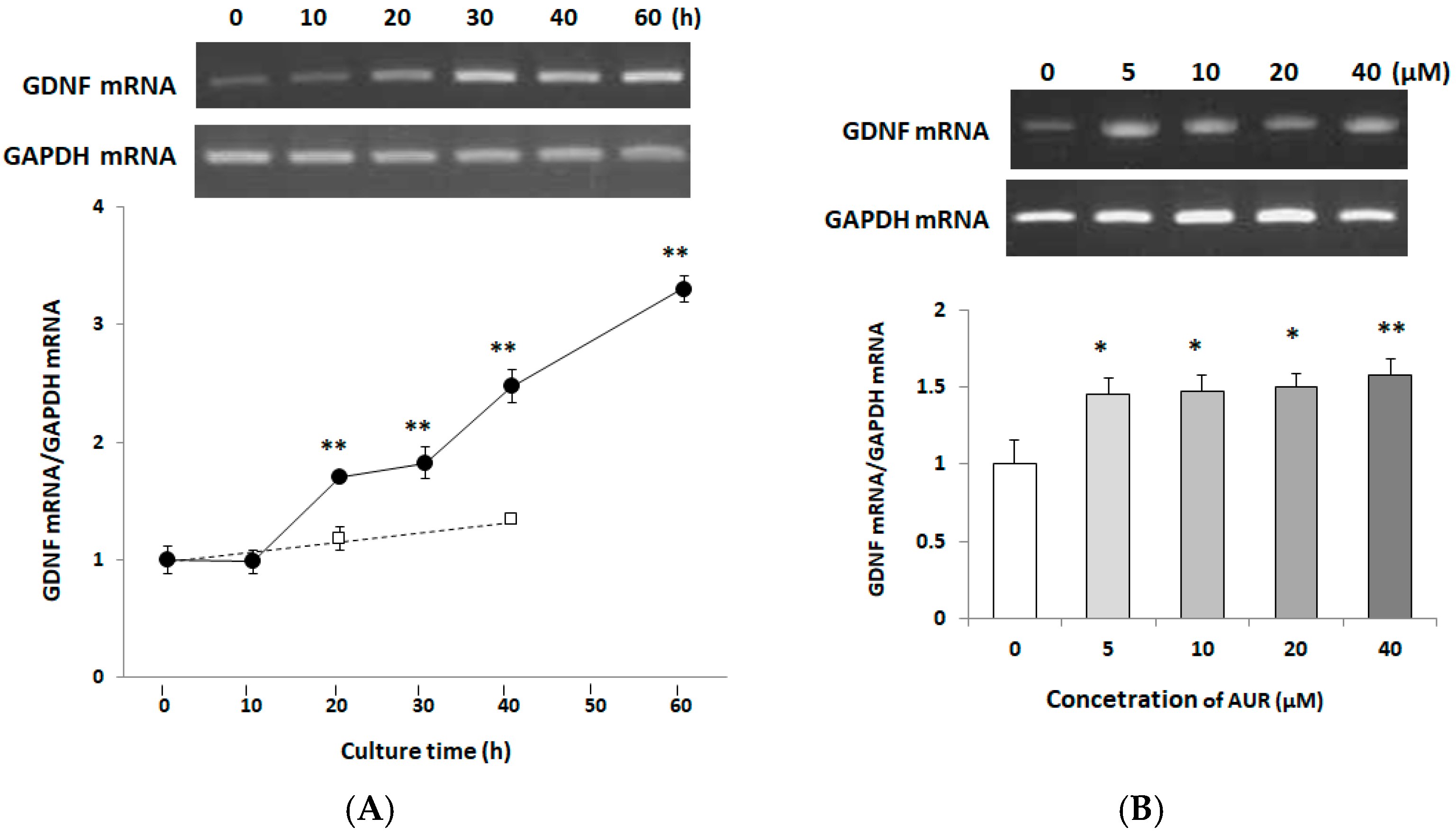
2.4. Effects of AUR on GDNF mRNA Expression in C6 Cells
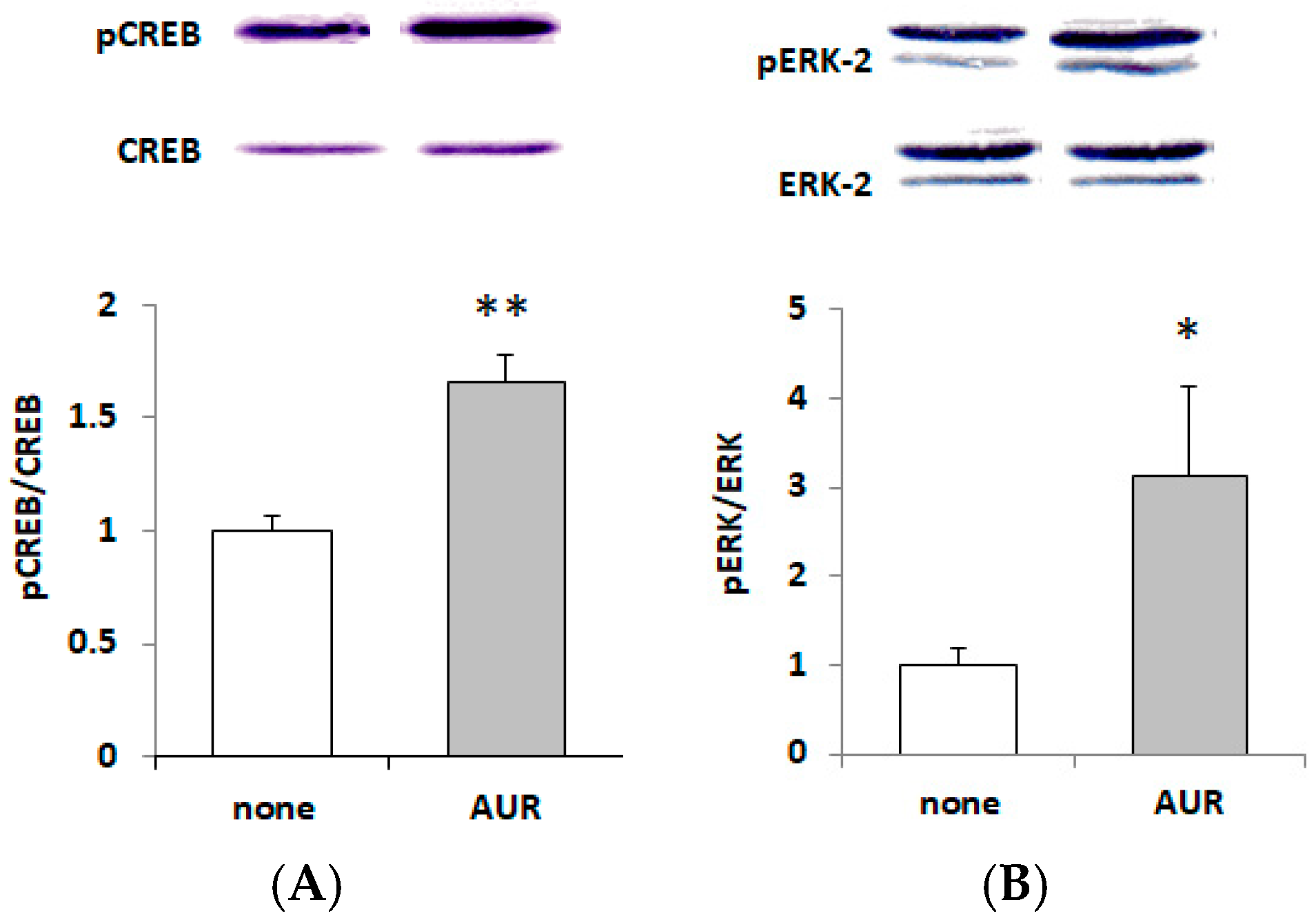
2.5. Effects of AUR on the Phosphorylation of CREB and ERK
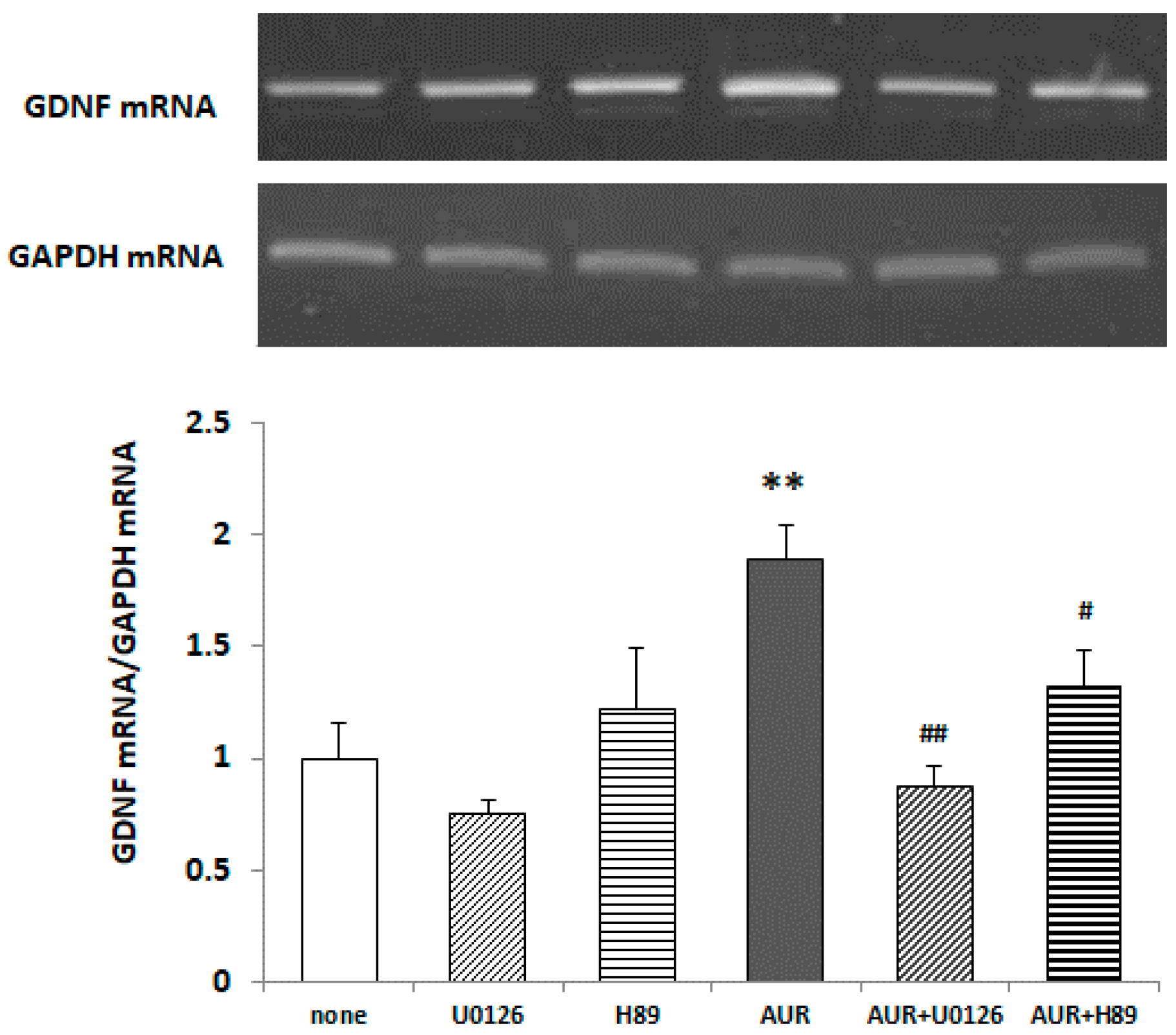
2.6. Effects of Inhibitors of ERK/PKA on AUR-Induced GDNF mRNA Expression in C6 Cells
3. Discussion
4. Materials and Methods
4.1. Chemical and Reagents
4.2. Cell Culture
4.3. Determination of Cell Viability
4.4. ELISA
4.5. Immunoblot Analysis
4.6. Total RNA Extraction and RT-PCR
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Genovese, S.; Epifano, F. Auraptene: A natural biologically active compound with multiple targets. Curr. Drug Targets 2011, 12, 381–386. [Google Scholar] [CrossRef]
- DeRosa, G.; Maffioli, P.; Sahebkar, A. Auraptene and Its Role in Chronic Diseases. Adv. Exp. Med. Biol. 2016, 929, 399–407. [Google Scholar]
- Okuyama, S.; Minami, S.; Shimada, N.; Makihata, N.; Nakajima, M.; Furukawa, Y. Anti-inflammatory and neuroprotective effects of auraptene, a citrus coumarin, following cerebral global ischemia in mice. Eur. J. Pharmacol. 2013, 699, 118–123. [Google Scholar] [CrossRef]
- Okuyama, S.; Yamamoto, K.; Mori, H.; Toyoda, N.; Yoshimura, M.; Amakura, Y.; Yoshida, T.; Sugawara, K.; Sudo, M.; Nakajima, M.; et al. Auraptene in the Peels of Citrus kawachiensis (Kawachi Bankan) Ameliorates Lipopolysaccharide-Induced Inflammation in the Mouse Brain. Evid. Based Complement. Altern. Med. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Okuyama, S.; Morita, M.; Kaji, M.; Amakura, Y.; Yoshimura, M.; Shimamoto, K.; Ookido, Y.; Nakajima, M.; Furukawa, Y. Auraptene Acts as an Anti-Inflammatory Agent in the Mouse Brain. Molecules 2015, 20, 20230–20239. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Semba, T.; Toyoda, N.; Epifano, F.; Genovese, S.; Fiorito, S.; Taddeo, V.A.; Sawamoto, A.; Nakajima, M.; Furukawa, Y. Auraptene and Other Prenyloxyphenylpropanoids Suppress Microglial Activation and Dopaminergic Neuronal Cell Death in a Lipopolysaccharide-Induced Model of Parkinson’s Disease. Int. J. Mol. Sci. 2016, 17, 1716. [Google Scholar] [CrossRef] [PubMed]
- Toyomoto, M.; Inoue, S.; Ohta, K.; Kuno, S.; Ohta, M.; Hayashi, K.; Ikeda, K. Production of NGF, BDNF and GDNF in mouse astrocyte cultures is strongly enhanced by a cerebral vasodilator, ifenprodil. Neurosci. Lett. 2005, 379, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Rocha, S.M.; Cristovão, A.C.; Campos, F.L.; Fonseca, C.P.; Baltazar, G. Astrocyte-derived GDNF is a potent inhibitor of microglial activation. Neurobiol. Dis. 2012, 47, 407–415. [Google Scholar] [CrossRef]
- Suter-Crazzolara, C.; Unsicker, K. GDNF mRNA levels are induced by FGF-2 in rat C6 glioblastoma cells. Mol. Brain Res. 1996, 41, 175–182. [Google Scholar] [CrossRef]
- Golan, M.; Schreiber, G.; Avissar, S. Antidepressants elevate GDNF expression and release from C₆ glioma cells in a β-arrestin1-dependent, CREB interactive pathway. Int. J. Neuropsychopharmacol. 2011, 14, 1289–1300. [Google Scholar] [CrossRef]
- Di Benedetto, B.; Kühn, R.; Nothdurfter, C.; Rein, T.; Wurst, W.; Rupprecht, R. N-desalkylquetiapine activates ERK1/2 to induce GDNF release in C6 glioma cells: A putative cellular mechanism for quetiapine as antidepressant. Neuropharmacology 2012, 62, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Eun, C.-S.; Lim, J.-S.; Lee, J.; Lee, S.-P.; Yang, S.-A. The protective effect of fermented Curcuma longa L. on memory dysfunction in oxidative stress-induced C6 gliomal cells, proinflammatory-activated BV2 microglial cells, and scopolamine-induced amnesia model in mice. BMC Complement. Altern. Med. 2017, 17, 367. [Google Scholar] [CrossRef] [PubMed]
- Sawamoto, A.; Okuyama, S.; Nakajima, M.; Furukawa, Y. Citrus flavonoid 3,5,6,7,8,3’,4’-heptamethoxy-flavone induces BDNF via cAMP/ERK/CREB signaling and reduces phosphodiesterase activity in C6 glioma cells. Pharmacol. Rep. 2019, 71, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Weingarten, D.P.; Callahan, J.W.; Sachar, K.; deVellis, J. Regulation of mRNAs for three enzymes in the glial cell model C6 cell line. J. Neurochem. 1984, 43, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- Mangoura, D.; Sakellaridis, N.; Jones, J.; Vernadakis, A. Early and late passage C-6 glial cell growth: Similarities with primary glial cells in culture. Neurochem. Res. 1989, 14, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Doherty, D.; Lile, J.; Bektesh, S.; Collins, F. GDNF: A glial cell line-derived neurotrophic factor for midbrain dopaminergic neurons. Science 1993, 260, 1130–1132. [Google Scholar] [CrossRef]
- Tomac, A.; Lindqvist, E.; Lin, L.-F.H.; Ögren, S.O.; Young, D.; Hoffer, B.J.; Olson, L. Protection and repair of the nigrostriatal dopaminergic system by GDNF in vivo. Nature 1995, 373, 335–339. [Google Scholar] [CrossRef]
- Airaksinen, M.S.; Saarma, M. The GDNF family: Signalling, biological functions and therapeutic value. Nat. Rev. Neurosci. 2002, 3, 383–394. [Google Scholar] [CrossRef]
- Tenenbaum, L.; Humbert-Claude, M. Glial Cell Line-Derived Neurotrophic Factor Gene Delivery in Parkinson’s Disease: A Delicate Balance between Neuroprotection, Trophic Effects, and Unwanted Compensatory Mechanisms. Front. Neuroanat. 2017, 11, 2336. [Google Scholar] [CrossRef]
- Pöyhönen, S.; Er, S.; Domanskyi, A.; Airavaara, M. Effects of Neurotrophic Factors in Glial Cells in the Central Nervous System: Expression and Properties in Neurodegeneration and Injury. Front. Physiol. 2019, 10, 486. [Google Scholar] [CrossRef]
- Arenas, E.; Trupp, M.; Åkerud, P.; Ibañez, C.F. GDNF prevents degeneration and promotes the phenotype of brain noradrenergic neurons in vivo. Neuron 1995, 15, 1465–1473. [Google Scholar] [CrossRef]
- Pascual, A.; Hidalgo-Figueroa, M.; I Piruat, J.; Pintado, C.O.; Gómez-Díaz, R.; López-Barneo, J. Absolute requirement of GDNF for adult catecholaminergic neuron survival. Nat. Neurosci. 2008, 11, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Egawa, H.; Osakada, M.; Baba, A.; Matsuda, T. Increase by FK960, a novel cognitive enhancer, in glial cell line-derived neurotrophic factor production in cultured rat astrocytes. Biochem. Pharmacol. 2004, 68, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Buscà, R.; Pouysségur, J.; Lenormand, P. ERK1 and ERK2 map kinases: Specific roles or functional redundancy? Front. Cell. Dev. Biol. 2016, 4, 53. [Google Scholar] [CrossRef] [PubMed]
- Impey, S.; Obrietan, K.; Wong, S.T.; Poser, S.; Yano, S.; Wayman, G.; Deloulme, J.C.; Chan, G.; Storm, D.R. Cross talk between ERK and PKA is required for Ca2+ stimulation of CREB-dependent transcription and ERK nuclear translocation. Neuron 1998, 21, 869–883. [Google Scholar] [CrossRef]
- Afshari, A.R.; Roshan, M.K.; Soukhtanloo, M.; Ghorbani, A.; Rahmani, F.; Jalili-Nik, M.; Vahedi, M.M.; Hoseini, A.; Sadeghnia, H.R.; Mollazadeh, H.; et al. Cytotoxic effects of auraptene against a human malignant glioblastoma cell line. Avicenna J. Phytomed. 2019, 9, 334–346. [Google Scholar]
- Bao, X.-Q.; Wang, L.; Yang, H.-Y.; Hou, L.-Y.; Wang, Q.-S.; Zhang, D. Induction of glial cell line-derived neurotrophic factor by the squamosamide derivative FLZ in astroglia has neuroprotective effects on dopaminergic neurons. Brain Res. Bull. 2019. [Google Scholar] [CrossRef]
- Yu, H.; Oh-Hashi, K.; Tanaka, T.; Sai, A.; Inoue, M.; Hirata, Y.; Kiuchi, K. Rehmannia glutinosa induces glial cell line-derived neurotrophic factor gene expression in astroglial cells via cPKC and ERK1/2 pathways independently. Pharmacol. Res. 2006, 54, 39–45. [Google Scholar] [CrossRef]
- Okuyama, S.; Kotani, Y.; Yamamoto, K.; Sawamoto, A.; Sugawara, K.; Sudo, M.; Ohkubo, Y.; Tamanaha, A.; Nakajima, M.; Furukawa, Y. The peel ofCitrus kawachiensis(kawachi bankan) ameliorates microglial activation, tau hyper-phosphorylation, and suppression of neurogenesis in the hippocampus of senescence-accelerated mice. Biosci. Biotechnol. Biochem. 2018, 82, 869–878. [Google Scholar] [CrossRef]
- Okuyama, S.; Yamamoto, K.; Mori, H.; Sawamoto, A.; Amakura, Y.; Yoshimura, M.; Tamanaha, A.; Ohkubo, Y.; Sugawara, K.; Sudo, M.; et al. Neuroprotective effect of Citrus kawachiensis (Kawachi Bankan) peels, a rich source of naringin, against global cerebral ischemia/reperfusion injury in mice. Biosci. Biotechnol. Biochem. 2018, 82, 1216–1224. [Google Scholar] [CrossRef]
- Okuyama, S.; Shinoka, W.; Nakamura, K.; Kotani, M.; Sawamoto, A.; Sugawara, K.; Sudo, M.; Nakajima, M.; Furukawa, Y. Suppressive effects of the peel of Citrus kawachiensis (Kawachi Bankan) on astroglial activation, tau phosphorylation, and inhibition of neurogenesis in the hippocampus of type 2 diabetic db/db mice. Biosci. Biotechnol. Biochem. 2018, 82, 1384–1395. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Kanzaki, T.; Kotani, Y.; Katoh, M.; Sawamoto, A.; Nakajima, M.; Furukawa, Y. Continual treatment with the peels of Citrus kawachiensis (Kawachi Bankan) protects the dopaminergic neuronal cell death in a lipopolysaccharide-induced model of Parkinson’s disease. J. Nutr. Sci. Vitaminol. 2019, 65, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Katoh, M.; Kanzaki, T.; Kotani, Y.; Amakura, Y.; Yoshimura, M.; Fukuda, N.; Tamai, T.; Sawamoto, A.; Nakajima, M.; et al. Auraptene/Naringin-Rich Fruit Juice of Citrus kawachiensis (Kawachi Bankan) Prevents Ischemia-Induced Neuronal Cell Death in Mouse Brain through Anti-Inflammatory Responses. J. Nutr. Sci. Vitaminol. 2019, 65, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Igase, M.; Okada, Y.; Ochi, M.; Igase, K.; Ochi, H.; Okuyama, S.; Furukawa, Y.; Ohyagi, Y. Auraptene in the peels of Citrus Kawachiensis (Kawachibankan) contributes to the preservation of cognitive function: A randomized, placebo-controlled, double-blind study in healthy volunteers. J. Prev. Alz. Dis. 2017, 3, 1–5. [Google Scholar]
- Budni, J.; Bellettini-Santos, T.; Mina, F.; Garcez, M.L.; Zugno, A.I. The involvement of BDNF, NGF and GDNF in aging and Alzheimer’s disease. Aging Dis. 2015, 6, 331–341. [Google Scholar] [CrossRef]
- Ghribi, O.; Herman, M.M.; Forbes, M.S.; DeWitt, D.A.; Savory, J. GDNF Protects against Aluminum-Induced Apoptosis in Rabbits by Upregulating Bcl-2 and Bcl-XL and Inhibiting Mitochondrial Bax Translocation. Neurobiol. Dis. 2001, 8, 764–773. [Google Scholar] [CrossRef]
- Revilla, S.; Ursulet, S.; Álvarez-López, M.J.; Castro-Freire, M.; Perpiña, U.; García-Mesa, Y.; Bortolozzi, A.; Giménez-Llort, L.; Kaliman, P.; Cristòfol, R.; et al. Lenti-GDNF Gene Therapy Protects Against Alzheimer’s Disease-Like Neuropathology in 3xTg-AD Mice and MC65 Cells. CNS Neurosci. Ther. 2014, 20, 961–972. [Google Scholar] [CrossRef]
- Airavaara, M.; Pletnikova, O.; Doyle, M.E.; Zhang, Y.E.; Troncoso, J.C.; Liu, Q.-R. Identification of Novel GDNF Isoforms and cis-Antisense GDNFOS Gene and Their Regulation in Human Middle Temporal Gyrus of Alzheimer Disease*. J. Boil. Chem. 2011, 286, 45093–45102. [Google Scholar] [CrossRef]
- Naoi, M.; Inaba-Hasegawa, K.; Shamoto-Nagai, M.; Maruyama, W. Neurotrophic function of phytochemicals for neuroprotection in aging and neurodegenerative disorders: Modulation of intracellular signaling and gene expression. J. Neural Transm. 2017, 124, 1515–1527. [Google Scholar] [CrossRef]
- Furukawa, Y.; Sawamoto, A.; Yamaoka, M.; Nakaya, M.; Hieda, Y.; Choshi, T.; Hatae, N.; Okuyama, S.; Nakajima, M.; Hibino, S. Effects of Carbazole Derivatives on Neurite Outgrowth and Hydrogen Peroxide-Induced Cytotoxicity in Neuro2a Cells. Molecules 2019, 24, 1366. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furukawa, Y.; Hara, R.-i.; Nakaya, M.; Okuyama, S.; Sawamoto, A.; Nakajima, M. Citrus Auraptene Induces Glial Cell Line-Derived Neurotrophic Factor in C6 Cells. Int. J. Mol. Sci. 2020, 21, 253. https://doi.org/10.3390/ijms21010253
Furukawa Y, Hara R-i, Nakaya M, Okuyama S, Sawamoto A, Nakajima M. Citrus Auraptene Induces Glial Cell Line-Derived Neurotrophic Factor in C6 Cells. International Journal of Molecular Sciences. 2020; 21(1):253. https://doi.org/10.3390/ijms21010253
Chicago/Turabian StyleFurukawa, Yoshiko, Ryu-ichi Hara, Makiko Nakaya, Satoshi Okuyama, Atsushi Sawamoto, and Mitsunari Nakajima. 2020. "Citrus Auraptene Induces Glial Cell Line-Derived Neurotrophic Factor in C6 Cells" International Journal of Molecular Sciences 21, no. 1: 253. https://doi.org/10.3390/ijms21010253
APA StyleFurukawa, Y., Hara, R.-i., Nakaya, M., Okuyama, S., Sawamoto, A., & Nakajima, M. (2020). Citrus Auraptene Induces Glial Cell Line-Derived Neurotrophic Factor in C6 Cells. International Journal of Molecular Sciences, 21(1), 253. https://doi.org/10.3390/ijms21010253