Hypoxia in the Initiation and Progression of Neuroblastoma Tumours

 ,
,  , and
, and 
Abstract
:
1. Introduction
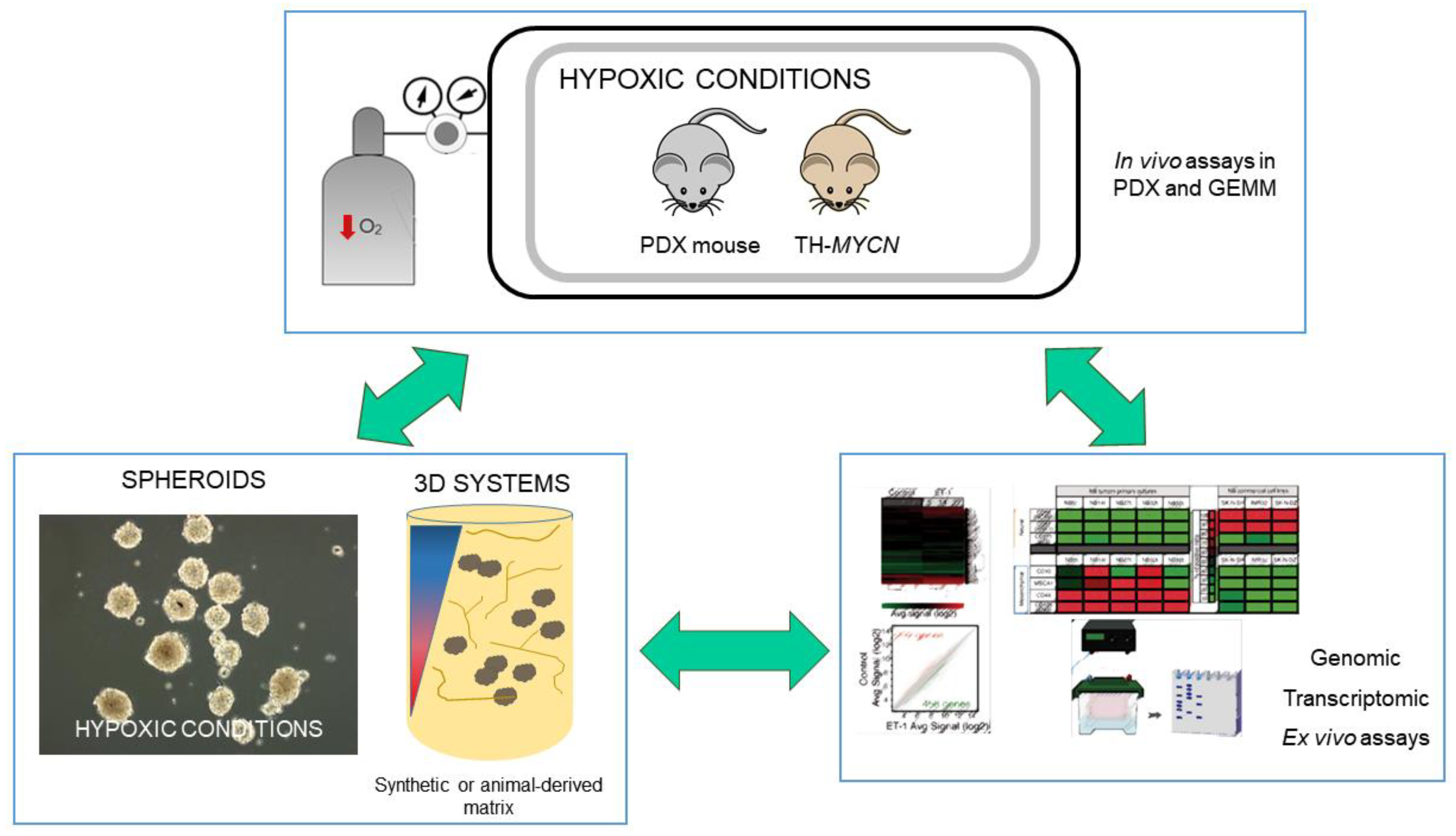
2. Approaches to the Study of Hypoxia in Neuroblastoma
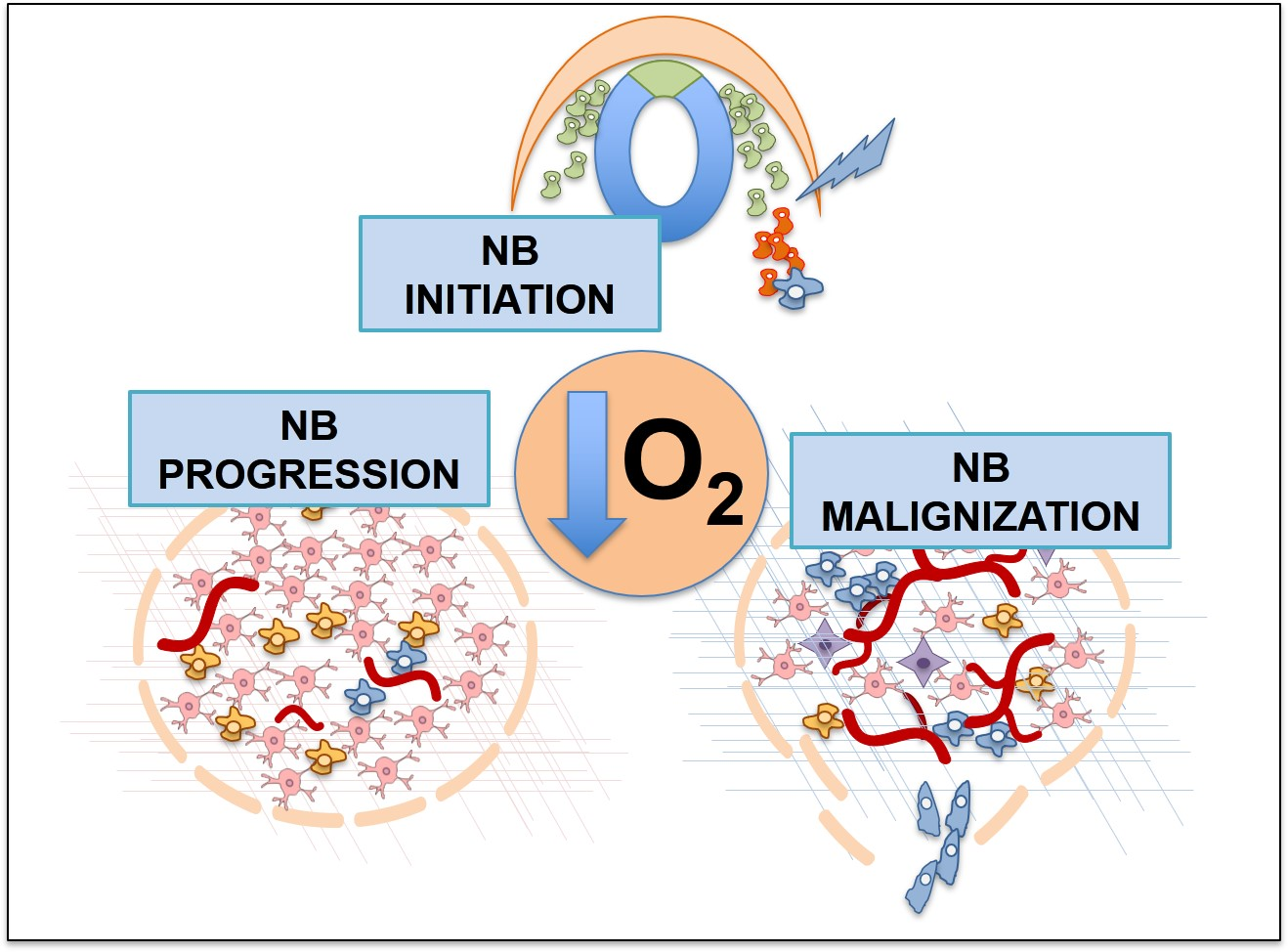
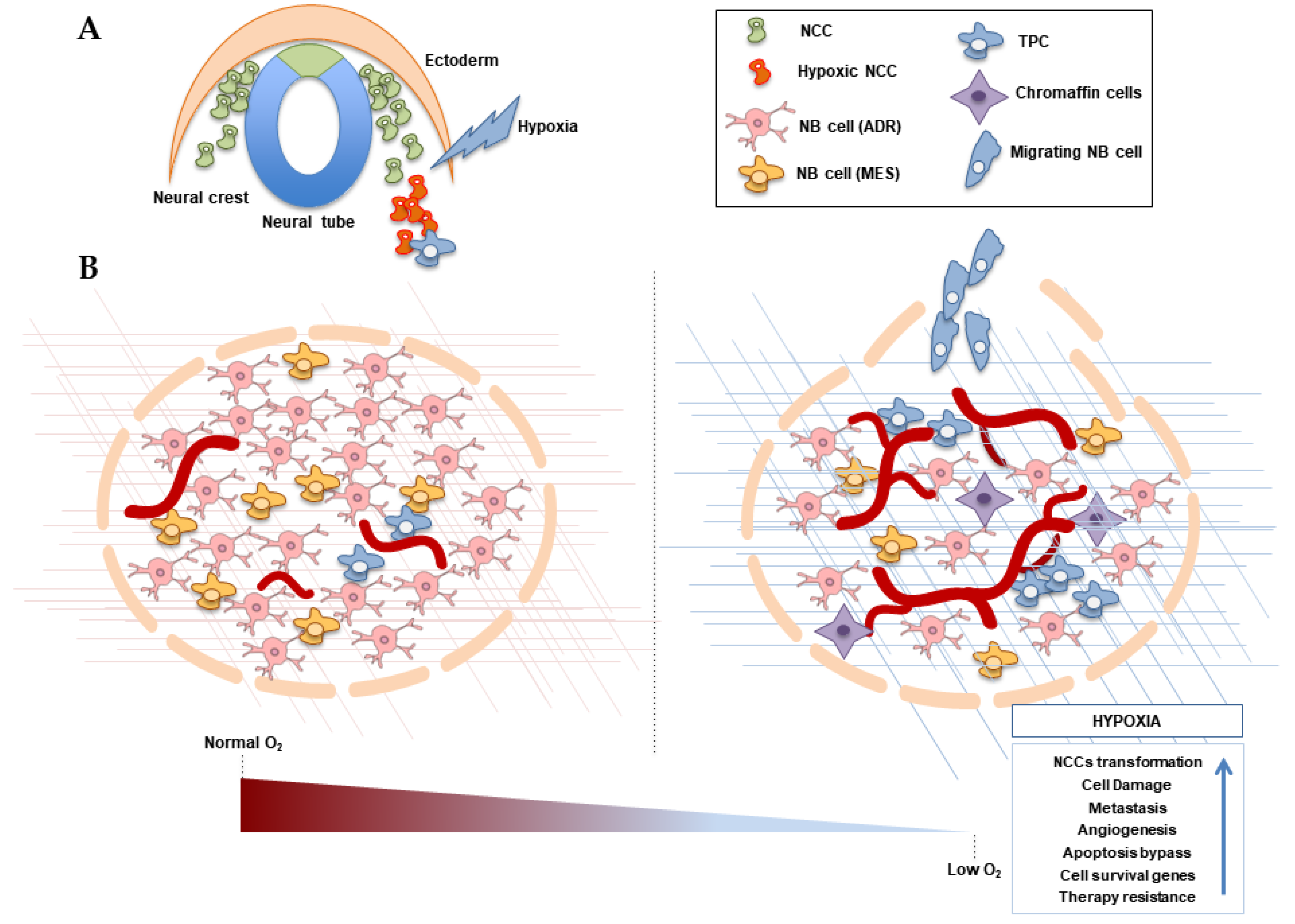
3. Hypoxia and the Origin of Neuroblastoma
4. Neuroblastoma Differentiation and Stemness in Hypoxic Contexts
5. Hypoxia and NB Cell Survival
6. Hypoxia and Metastasis in Neuroblastoma
7. Hypoxia-Related Therapies
8. Future Perspectives
Funding
Conflicts of Interest
References
- Marshall, G.M.; Carter, D.R.; Cheung, B.B.; Liu, T.; Mateos, M.K.; Meyerowitz, J.G.; Weiss, W.A. The prenatal origins of cancer. Nat. Rev. Cancer 2014, 14, 277–289. [Google Scholar] [CrossRef] [Green Version]
- Cheung, N.K.V.; Dyer, M.A. Neuroblastoma: Developmental biology, cancer genomics and immunotherapy. Nat. Rev. Cancer 2013, 13, 397–411. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.A.; Seibel, N.L.; Altekruse, S.F.; Ries, L.A.G.; Melbert, D.L.; O’Leary, M.; Smith, F.O.; Reaman, G.H. Outcomes for Children and Adolescents With Cancer: Challenges for the Twenty-First Century. J. Clin. Oncol. 2010, 28, 2625–2634. [Google Scholar] [CrossRef] [PubMed]
- Matthay, K.K.; Maris, J.M.; Schleiermacher, G.; Nakagawara, A.; Mackall, C.L.; Diller, L.; Weiss, W.A. Neuroblastoma. Nat. Rev. Dis. Prim. 2016, 2, 16078. [Google Scholar] [CrossRef] [PubMed]
- Von Stedingk, K.; Gisselsson, D.; Bexell, D. Multidimensional intratumour heterogeneity in neuroblastoma. Oncotarget 2019, 10, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M.B.; Leslie, M.T.; Gaskins, H.R.; Kenis, P.J.A. Methods to study the tumor microenvironment under controlled oxygen conditions. Trends Biotechnol. 2014, 32, 556–563. [Google Scholar] [CrossRef] [Green Version]
- Weiss, W.A. Targeted expression of MYCN causes neuroblastoma in transgenic mice. EMBO J. 1997, 16, 2985–2995. [Google Scholar] [CrossRef]
- Kiyonari, S.; Kadomatsu, K. Neuroblastoma models for insights into tumorigenesis and new therapies. Expert Opin. Drug Discov. 2015, 10, 53–62. [Google Scholar] [CrossRef]
- Chesler, L.; Goldenberg, D.D.; Seales, I.T.; Satchi-Fainaro, R.; Grimmer, M.; Collins, R.; Struett, C.; Nguyen, K.N.; Kim, G.; Tihan, T.; et al. Malignant Progression and Blockade of Angiogenesis in a Murine Transgenic Model of Neuroblastoma. Cancer Res. 2007, 67, 9435–9442. [Google Scholar] [CrossRef] [Green Version]
- Teitz, T.; Inoue, M.; Valentine, M.B.; Zhu, K.; Rehg, J.E.; Zhao, W.; Finkelstein, D.; Wang, Y.D.; Johnson, M.D.; Calabrese, C.; et al. Th-MYCN Mice with Caspase-8 Deficiency Develop Advanced Neuroblastoma with Bone Marrow Metastasis. Cancer Res. 2013, 73, 4086–4097. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, E.; Hong, S.H.; Galli, S.; Trinh, E.; Wietlisbach, L.; Misiukiewicz, S.F.; Tilan, J.U.; Chen, Y.S.; Schlegel, R.; Kitlinska, J. Murine neuroblastoma cell lines developed by conditional reprogramming preserve heterogeneous phenotypes observed in vivo. Lab. Investig. 2019. [Google Scholar] [CrossRef] [PubMed]
- Braekeveldt, N.; Bexell, D. Patient-derived xenografts as preclinical neuroblastoma models. Cell Tissue Res. 2018, 372, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Braekeveldt, N.; Wigerup, C.; Gisselsson, D.; Mohlin, S.; Merselius, M.; Beckman, S.; Jonson, T.; Börjesson, A.; Backman, T.; Tadeo, I.; et al. Neuroblastoma patient-derived orthotopic xenografts retain metastatic patterns and geno- and phenotypes of patient tumours. Int. J. Cancer 2015, 136, 252–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulon, A.; Flahaut, M.; Mühlethaler-Mottet, A.; Meier, R.; Liberman, J.; Balmas-Bourloud, K.; Nardou, K.; Yan, P.; Tercier, S.; Joseph, J.M.; et al. Functional Sphere Profiling Reveals the Complexity of Neuroblastoma Tumor-Initiating Cell Model. Neoplasia 2011, 13, 991–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.Y.C.; Lin, D.; Gout, P.W.; Collins, C.C.; Xu, Y.; Wang, Y. Lessons from patient-derived xenografts for better in vitro modeling of human cancer. Adv. Drug Deliv. Rev. 2014, 79–80, 222–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, Y.; Ta, H.P.; Yen, D.P.; Lee, S.S.; Raola, S.; Shen, K. A Microdevice Platform Recapitulating Hypoxic Tumor Microenvironments. Sci. Rep. 2017, 7, 15233. [Google Scholar] [CrossRef] [Green Version]
- Tsubota, S.; Kadomatsu, K. Origin and initiation mechanisms of neuroblastoma. Cell Tissue Res. 2018, 372, 211–221. [Google Scholar] [CrossRef]
- Bluhm, E.; McNeil, D.E.; Cnattingius, S.; Gridley, G.; El Ghormli, L.; Fraumeni, J.F. Prenatal and perinatal risk factors for neuroblastoma. Int. J. Cancer 2008, 123, 2885–2890. [Google Scholar] [CrossRef]
- Fajersztajn, L.; Veras, M.M. Hypoxia: From Placental Development to Fetal Programming. Birth Defects Res. 2017, 109, 1377–1385. [Google Scholar] [CrossRef]
- Scully, D.; Keane, E.; Batt, E.; Karunakaran, P.; Higgins, D.F.; Itasaki, N. Hypoxia promotes production of neural crest cells in the embryonic head. Development 2016, 143, 1742–1752. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.C.; Hsia, C.W.; Ho, C.W.; Liang, C.M.; Chen, C.M.; Huang, K.L.; Kang, B.H.; Chen, Y.H. Hypoxia and hyperoxia differentially control proliferation of rat neural crest stem cells via distinct regulatory pathways of the HIF1α-CXCR4 and TP53-TPM1 proteins. Dev. Dyn. 2017, 246, 162–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barriga, E.H.; Maxwell, P.H.; Reyes, A.E.; Mayor, R. The hypoxia factor Hif-1α controls neural crest chemotaxis and epithelial to mesenchymal transition. J. Cell Biol. 2013, 201, 759–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espina, J.A.; Marchant, C.L.; De Ferrari, G.V.; Reyes, A.E. Pdgf1aa regulates zebrafish neural crest cells migration through Hif-1 in an oxygen-independent manner. Mech. Dev. 2018, 154, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Holmquist-Mengelbier, L.; Fredlund, E.; Löfstedt, T.; Noguera, R.; Navarro, S.; Nilsson, H.; Pietras, A.; Vallon-Christersson, J.; Borg, Å.; Gradin, K.; et al. Recruitment of HIF-1α and HIF-2α to common target genes is differentially regulated in neuroblastoma: HIF-2α promotes an aggressive phenotype. Cancer Cell 2006, 10, 413–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, T.; Gallagher, D.; Pascual, A.; Lygate, C.A.; de Bono, J.P.; Nicholls, L.G.; Ortega-Saenz, P.; Oster, H.; Wijeyekoon, B.; Sutherland, A.I.; et al. Abnormal Sympathoadrenal Development and Systemic Hypotension in PHD3-/-Mice. Mol. Cell. Biol. 2008, 28, 3386–3400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantilena, S.; Pastorino, F.; Pezzolo, A.; Chayka, O.; Pistoia, V.; Ponzoni, M.; Sala, A. Frizzled receptor 6 marks rare, highly tumourigenic stem-like cells in mouse and human neuroblastomas. Oncotarget 2011, 2, 976. [Google Scholar] [CrossRef] [Green Version]
- Bristow, R.G. Hypoxia, DNA Repair, and Genetic Instability. AACR Educ. Book 2008, 2008, 287–291. [Google Scholar] [CrossRef]
- Luoto, K.R.; Kumareswaran, R.; Bristow, R.G. Tumor hypoxia as a driving force in genetic instability. Genome Integr. 2013, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, V.; Hoey, C.; Liu, L.Y.; Lalonde, E.; Ray, J.; Livingstone, J.; Lesurf, R.; Shiah, Y.J.; Vujcic, T.; Huang, X.; et al. Molecular landmarks of tumor hypoxia across cancer types. Nat. Genet. 2019, 51, 308–318. [Google Scholar] [CrossRef]
- Molenaar, J.J.; Koster, J.; Zwijnenburg, D.A.; van Sluis, P.; Valentijn, L.J.; van der Ploeg, I.; Hamdi, M.; van Nes, J.; Westerman, B.A.; van Arkel, J.; et al. Sequencing of neuroblastoma identifies chromothripsis and defects in neuritogenesis genes. Nature 2012, 483, 589–593. [Google Scholar] [CrossRef]
- Farina, A.R.; Tacconelli, A.; Cappabianca, L.; Cea, G.; Panella, S.; Chioda, A.; Romanelli, A.; Pedone, C.; Gulino, A.; Mackay, A.R. The Alternative TrkAIII Splice Variant Targets the Centrosome and Promotes Genetic Instability. Mol. Cell. Biol. 2009, 29, 4812–4830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohn, S.L.; Pearson, A.D.J.; London, W.B.; Monclair, T.; Ambros, P.F.; Brodeur, G.M.; Faldum, A.; Hero, B.; Iehara, T.; Machin, D.; et al. The International Neuroblastoma Risk Group (INRG) Classification System: An INRG Task Force Report. J. Clin. Oncol. 2009, 27, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Boeva, V.; Louis-Brennetot, C.; Peltier, A.; Durand, S.; Pierre-Eugène, C.; Raynal, V.; Etchevers, H.C.; Thomas, S.; Lermine, A.; Daudigeos-Dubus, E.; et al. Heterogeneity of neuroblastoma cell identity defined by transcriptional circuitries. Nat. Genet. 2017, 49, 1408–1413. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.M.; Colmenero-Repiso, A.; Gómez-Muñoz, M.A.; Rodríguez-Prieto, I.; Aguilar-Morante, D.; Ramírez, G.; Márquez, C.; Cabello, R.; Pardal, R. CD44-high neural crest stem-like cells are associated with tumour aggressiveness and poor survival in neuroblastoma tumours. EBioMedicine 2019, 49, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Bahmad, H.F.; Chamaa, F.; Assi, S.; Chalhoub, R.M.; Abou-Antoun, T.; Abou-Kheir, W. Cancer Stem Cells in Neuroblastoma: Expanding the Therapeutic Frontier. Front. Mol. Neurosci. 2019, 12, 131. [Google Scholar] [CrossRef] [Green Version]
- Hedborg, F.; Ullerås, E.; Grimelius, L.; Wassberg, E.; Maxwell, P.H.; Hero, B.; Berthold, F.; Schilling, F.; Harms, D.; Sandstedt, B.; et al. Evidence for hypoxia-induced neuronal-to-chromaffin metaplasia in neuroblastoma. FASEB J. 2003, 17, 598–609. [Google Scholar] [CrossRef]
- Hedborg, F.; Fischer-Colbrie, R.; Östlin, N.; Sandstedt, B.; Tran, M.G.B.; Maxwell, P.H. Differentiation in Neuroblastoma: Diffusion-Limited Hypoxia Induces Neuro-Endocrine Secretory Protein 55 and Other Markers of a Chromaffin Phenotype. PLoS ONE 2010, 5, 1–15. [Google Scholar] [CrossRef]
- Jogi, A.; Ora, I.; Nilsson, H.; Lindeheim, A.; Makino, Y.; Poellinger, L.; Axelson, H.; Pahlman, S. Hypoxia alters gene expression in human neuroblastoma cells toward an immature and neural crest-like phenotype. Proc. Natl. Acad. Sci. USA 2002, 99, 7021–7026. [Google Scholar] [CrossRef] [Green Version]
- Fredlund, E.; Ringner, M.; Maris, J.M.; Pahlman, S. High Myc pathway activity and low stage of neuronal differentiation associate with poor outcome in neuroblastoma. Proc. Natl. Acad. Sci. USA 2008, 105, 14094–14099. [Google Scholar] [CrossRef] [Green Version]
- Jögi, A.; Vallon-Christersson, J.; Holmquist, L.; Axelson, H.; Borg, Å.; Påhlman, S. Human neuroblastoma cells exposed to hypoxia: Induction of genes associated with growth, survival, and aggressive behavior. Exp. Cell Res. 2004, 295, 469–487. [Google Scholar] [CrossRef]
- Axelson, H.; Fredlund, E.; Ovenberger, M.; Landberg, G.; Påhlman, S. Hypoxia-induced dedifferentiation of tumor cells—A mechanism behind heterogeneity and aggressiveness of solid tumors. Semin. Cell Dev. Biol. 2005, 16, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Covello, K.L. HIF-2 regulates Oct-4: Effects of hypoxia on stem cell function, embryonic development, and tumor growth. Genes Dev. 2006, 20, 557–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabrese, C.; Poppleton, H.; Kocak, M.; Hogg, T.L.; Fuller, C.; Hamner, B.; Oh, E.Y.; Gaber, M.W.; Finklestein, D.; Allen, M.; et al. A Perivascular Niche for Brain Tumor Stem Cells. Cancer Cell 2007, 11, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Pietras, A.; Gisselsson, D.; Øra, I.; Noguera, R.; Beckman, S.; Navarro, S.; Påhlman, S. High levels of HIF-2α highlight an immature neural crest-like neuroblastoma cell cohort located in a perivascular niche. J. Pathol. 2007, 214, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Das, B.; Tsuchida, R.; Malkin, D.; Koren, G.; Baruchel, S.; Yeger, H. Hypoxia Enhances Tumor Stemness by Increasing the Invasive and Tumorigenic Side Population Fraction. Stem Cells 2008, 26, 1818–1830. [Google Scholar] [CrossRef] [PubMed]
- Pietras, A.; Hansford, L.M.; Johnsson, A.S.; Bridges, E.; Sjolund, J.; Gisselsson, D.; Rehn, M.; Beckman, S.; Noguera, R.; Navarro, S.; et al. HIF-2 maintains an undifferentiated state in neural crest-like human neuroblastoma tumor-initiating cells. Proc. Natl. Acad. Sci. USA 2009, 106, 16805–16810. [Google Scholar] [CrossRef] [Green Version]
- Cimmino, F.; Pezone, L.; Avitabile, M.; Persano, L.; Vitale, M.; Sassi, M.; Bresolin, S.; Serafin, V.; Zambrano, N.; Scaloni, A.; et al. Proteomic Alterations in Response to Hypoxia Inducible Factor 2α in Normoxic Neuroblastoma Cells. J. Proteome Res. 2016, 15, 3643–3655. [Google Scholar] [CrossRef]
- Guo, X.; Chen, Q.R.; Song, Y.K.; Wei, J.S.; Khan, J. Exon array analysis reveals neuroblastoma tumors have distinct alternative splicing patterns according to stage and MYCN amplification status. BMC Med. Genom. 2011, 4, 35. [Google Scholar] [CrossRef] [Green Version]
- Applebaum, M.A.; Jha, A.R.; Kao, C.; Hernandez, K.M.; DeWane, G.; Salwen, H.R.; Chlenski, A.; Dobratic, M.; Mariani, C.J.; Godley, L.A.; et al. Integrative genomics reveals hypoxia inducible genes that are associated with a poor prognosis in neuroblastoma patients. Oncotarget 2016, 7, 76816–76826. [Google Scholar] [CrossRef] [Green Version]
- Cangelosi, D.; Pelassa, S.; Morini, M.; Conte, M.; Bosco, M.C.; Eva, A.; Sementa, A.R.; Varesio, L. Artificial neural network classifier predicts neuroblastoma patients’ outcome. BMC Bioinform. 2016, 17, 347. [Google Scholar] [CrossRef] [Green Version]
- Ognibene, M.; Cangelosi, D.; Morini, M.; Segalerba, D.; Bosco, M.C.; Sementa, A.R.; Eva, A.; Varesio, L. Immunohistochemical analysis of PDK1, PHD3 and HIF-1α expression defines the hypoxic status of neuroblastoma tumors. PLoS ONE 2017, 12, e0187206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerlund, I.; Shi, Y.; Holmberg, J. EPAS1/HIF2α correlates with features of low-risk neuroblastoma and with adrenal chromaffin cell differentiation during sympathoadrenal development. Biochem. Biophys. Res. Commun. 2019, 508, 1233–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlan, A.; Dyachuk, V.; Kastriti, M.E.; Calvo-Enrique, L.; Abdo, H.; Hadjab, S.; Chontorotzea, T.; Akkuratova, N.; Usoskin, D.; Kamenev, D.; et al. Multipotent peripheral glial cells generate neuroendocrine cells of the adrenal medulla. Science 2017, 357, eaal3753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Groningen, T.; Koster, J.; Valentijn, L.J.; Zwijnenburg, D.A.; Akogul, N.; Hasselt, N.E.; Broekmans, M.; Haneveld, F.; Nowakowska, N.E.; Bras, J.; et al. Neuroblastoma is composed of two super-enhancer-associated differentiation states. Nat. Genet. 2017, 49, 1261–1266. [Google Scholar] [CrossRef]
- Hockel, M.; Vaupel, P. Tumor Hypoxia: Definitions and Current Clinical, Biologic, and Molecular Aspects. JNCI J. Natl. Cancer Inst. 2001, 93, 266–276. [Google Scholar] [CrossRef] [Green Version]
- Gordan, J.D.; Bertout, J.A.; Hu, C.J.; Diehl, J.A.; Simon, M.C. HIF-2α Promotes Hypoxic Cell Proliferation by Enhancing c-Myc Transcriptional Activity. Cancer Cell 2007, 11, 335–347. [Google Scholar] [CrossRef] [Green Version]
- Zaatiti, H.; Abdallah, J.; Nasr, Z.; Khazen, G.; Sandler, A.; Abou-Antoun, T. Tumorigenic proteins upregulated in the MYCN-amplified IMR-32 human neuroblastoma cells promote proliferation and migration. Int. J. Oncol. 2018, 52, 787–803. [Google Scholar] [CrossRef]
- Yin, C.P.; Guan, S.H.; Zhang, B.; Wang, X.X.; Yue, S.W. Upregulation of HIF-1α protects neuroblastoma cells from hypoxia-induced apoptosis in a RhoA-dependent manner. Mol. Med. Rep. 2015, 12, 7123–7131. [Google Scholar] [CrossRef]
- Skuli, N.; Monferran, S.; Delmas, C.; Lajoie-Mazenc, I.; Favre, G.; Toulas, C.; Cohen-Jonathan-Moyal, E. Activation of RhoB by Hypoxia Controls Hypoxia-Inducible Factor-1α Stabilization through Glycogen Synthase Kinase-3 in U87 Glioblastoma Cells. Cancer Res. 2006, 66, 482–489. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Yin, C.P.; Zhao, Q.; Yue, S.W. Upregulation of HIF-1α by Hypoxia Protect Neuroblastoma Cells from Apoptosis by Promoting Survivin Expression. Asian Pac. J. Cancer Prev. 2014, 15, 8251–8257. [Google Scholar] [CrossRef]
- Das, B.; Yeger, H.; Tsuchida, R.; Torkin, R.; Gee, M.F.W.; Thorner, P.S.; Shibuya, M.; Malkin, D.; Baruchel, S. A Hypoxia-Driven Vascular Endothelial Growth Factor/Flt1 Autocrine Loop Interacts with Hypoxia-Inducible Factor-1α through Mitogen-Activated Protein Kinase/Extracellular Signal-Regulated Kinase 1/2 Pathway in Neuroblastoma. Cancer Res. 2005, 65, 7267–7275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, J.H.; Jang, J.E.; Kang, C.M.; Chung, H.Y.; Kim, N.D.; Kim, K.W. Hypoxia-induced VEGF enhances tumor survivability via suppression of serum deprivation-induced apoptosis. Oncogene 2000, 19, 4621–4631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Weng, Q.; Zhang, L.; He, Q.; Yang, B. VEGF and Bcl-2 Interact Via MAPKs Signaling Pathway in the Response to Hypoxia in Neuroblastoma. Cell. Mol. Neurobiol. 2009, 29, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Crowder, S.W.; Balikov, D.A.; Hwang, Y.S.; Sung, H.J. Cancer Stem Cells Under Hypoxia as a Chemoresistance Factor in the Breast and Brain. Curr. Pathobiol. Rep. 2014, 2, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Hussein, D. Chronic hypoxia promotes hypoxia-inducible factor-1 -dependent resistance to etoposide and vincristine in neuroblastoma cells. Mol. Cancer Ther. 2006, 5, 2241–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, J.Y.; Chun, Y.S.; Kim, T.Y.; Kim, H.L.; Kim, M.S.; Park, J.W. Hypoxia-Inducible Factor 1alpha- Mediated Resistance to Phenolic Anticancer. Chemotherapy 2004, 50, 119–126. [Google Scholar] [CrossRef]
- Wentzel, J.F.; Lewies, A.; Bronkhorst, A.J.; van Dyk, E.; du Plessis, L.H.; Pretorius, P.J. Exposure to high levels of fumarate and succinate leads to apoptotic cytotoxicity and altered global DNA methylation profiles in vitro. Biochimie 2017, 135, 28–34. [Google Scholar] [CrossRef]
- Thienpont, B.; Steinbacher, J.; Zhao, H.; D’Anna, F.; Kuchnio, A.; Ploumakis, A.; Ghesquière, B.; Van Dyck, L.; Boeckx, B.; Schoonjans, L.; et al. Tumour hypoxia causes DNA hypermethylation by reducing TET activity. Nature 2016, 537, 63–68. [Google Scholar] [CrossRef]
- Cimmino, F.; Avitabile, M.; Lasorsa, V.A.; Montella, A.; Pezone, L.; Cantalupo, S.; Visconte, F.; Corrias, M.V.; Iolascon, A.; Capasso, M. HIF-1 transcription activity: HIF1A driven response in normoxia and in hypoxia. BMC Med. Genet. 2019, 20, 37. [Google Scholar] [CrossRef] [Green Version]
- Tweddle, D.A.; Pearson, A.D.J.; Haber, M.; Norris, M.D.; Xue, C.; Flemming, C.; Lunec, J. The p53 pathway and its inactivation in neuroblastoma. Cancer Lett. 2003, 197, 93–98. [Google Scholar] [CrossRef]
- Robertson, E.D.; Semenchenko, K.; Wasylyk, B. Crosstalk Between Mdm2, p53 and HIF1-α: Distinct Responses to Oxygen Stress and Implications for Tumour Hypoxia. In Sub-Cellular Biochemistry; Springer: Dordrecht, The Netherlands, 2014; pp. 199–214. [Google Scholar] [CrossRef]
- DuBois, S.G.; Kalika, Y.; Lukens, J.N.; Brodeur, G.M.; Seeger, R.C.; Atkinson, J.B.; Haase, G.M.; Black, C.T.; Perez, C.; Shimada, H.; et al. Metastatic Sites in Stage IV and IVS Neuroblastoma Correlate With Age, Tumor Biology, and Survival. J. Pediatr. Hematol. Oncol. 1999, 21, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Muz, B.; de la Puente, P.; Azab, F.; Azab, A.K. The role of hypoxia in cancer progression, angiogenesis, metastasis, and resistance to therapy. Hypoxia 2015, 3, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, H.; Li, J.; Entenberg, D.; Xue, A.; Wang, W.; Condeelis, J. Direct visualization of the phenotype of hypoxic tumor cells at single cell resolution in vivo using a new hypoxia probe. IntraVital 2016, 5, e1187803. [Google Scholar] [CrossRef] [PubMed]
- Fluegen, G.; Avivar-Valderas, A.; Wang, Y.; Padgen, M.R.; Williams, J.K.; Nobre, A.R.; Calvo, V.; Cheung, J.F.; Bravo-Cordero, J.J.; Entenberg, D.; et al. Phenotypic heterogeneity of disseminated tumour cells is preset by primary tumour hypoxic microenvironments. Nat. Cell Biol. 2017, 19, 120–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshizuka, N.; Moriuchi, R.; Mori, T.; Yamada, K.; Hasegawa, S.; Maeda, T.; Shimada, T.; Yamada, Y.; Kamihira, S.; Tomonaga, M.; et al. An Alternative Transcript Derived from the Trio Locus Encodes a Guanosine Nucleotide Exchange Factor with Mouse Cell-transforming Potential. J. Biol. Chem. 2004, 279, 43998–44004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oeggerli, M.; Tomovska, S.; Schraml, P.; Calvano-Forte, D.; Schafroth, S.; Simon, R.; Gasser, T.; Mihatsch, M.J.; Sauter, G. E2F3 amplification and overexpression is associated with invasive tumor growth and rapid tumor cell proliferation in urinary bladder cancer. Oncogene 2004, 23, 5616–5623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ten Berge, R.L.; Meijer, C.J.L.M.; Dukers, D.F.; Kummer, J.A.; Bladergroen, B.A.; Vos, W.; Hack, C.E.; Ossenkoppele, G.J.; Oudejans, J.J. Expression levels of apoptosis-related proteins predict clinical outcome in anaplastic large cell lymphoma. Blood 2002, 99, 4540–4546. [Google Scholar] [CrossRef] [Green Version]
- Van Houdt, I.S. Expression of the Apoptosis Inhibitor Protease Inhibitor 9 Predicts Clinical Outcome in Vaccinated Patients with Stage III and IV Melanoma. Clin. Cancer Res. 2005, 11, 6400–6407. [Google Scholar] [CrossRef] [Green Version]
- Poomthavorn, P.; Wong, S.H.X.; Higgins, S.; Werther, G.A.; Russo, V.C. Activation of a prometastatic gene expression program in hypoxic neuroblastoma cells. Endocr.-Relat. Cancer 2009, 16, 991–1004. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhang, M.; Xing, L.; Wang, Y.; Xiao, Y.; Wu, Y. HIF-1α Contributes to Proliferation and Invasiveness of Neuroblastoma Cells via SHH Signaling. PLoS ONE 2015, 10, e0121115. [Google Scholar] [CrossRef]
- Wickström, M.; Dyberg, C.; Shimokawa, T.; Milosevic, J.; Baryawno, N.; Fuskevåg, O.M.; Larsson, R.; Kogner, P.; Zaphiropoulos, P.G.; Johnsen, J.I. Targeting the hedgehog signal transduction pathway at the level of GLI inhibits neuroblastoma cell growth in vitro and in vivo. Int. J. Cancer 2013, 132, 1516–1524. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, A.; Rice, M.; Lévy, R.; Pizer, B.L.; Losty, P.D.; Moss, D.; Sée, V. Cellular memory of hypoxia elicits neuroblastoma metastasis and enables invasion by non-aggressive neighbouring cells. Oncogenesis 2015, 4, e138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieminen, A.L.; Qanungo, S.; Schneider, E.A.; Jiang, B.H.; Agani, F.H. Mdm2 and HIF-1α interaction in tumor cells during hypoxia. J. Cell. Physiol. 2005, 204, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Patterson, D.M.; Gao, D.; Trahan, D.N.; Johnson, B.A.; Ludwig, A.; Barbieri, E.; Chen, Z.; Diaz-Miron, J.; Vassilev, L.; Shohet, J.M.; et al. Effect of MDM2 and vascular endothelial growth factor inhibition on tumor angiogenesis and metastasis in neuroblastoma. Angiogenesis 2011, 14, 255–266. [Google Scholar] [CrossRef]
- Sun, W.; Zhang, L.; Hou, L.; Ju, C.; Zhao, S.; Wei, Y. Isatin inhibits SH-SY5Y neuroblastoma cell invasion and metastasis through MAO/HIF-1α/CXCR4 signaling. Anti-Cancer Drugs 2017, 28, 645–653. [Google Scholar] [CrossRef]
- Zhang, H.; Pu, J.; Qi, T.; Qi, M.; Yang, C.; Li, S.; Huang, K.; Zheng, L.; Tong, Q. MicroRNA-145 inhibits the growth, invasion, metastasis and angiogenesis of neuroblastoma cells through targeting hypoxia-inducible factor 2 alpha. Oncogene 2014, 33, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Rey, S.; Schito, L.; Wouters, B.G.; Eliasof, S.; Kerbel, R.S. Targeting Hypoxia-Inducible Factors for Antiangiogenic Cancer Therapy. Trends Cancer 2017, 3, 529–541. [Google Scholar] [CrossRef]
- Garner, E.; Beierle, E. Cancer Stem Cells and Their Interaction with the Tumor Microenvironment in Neuroblastoma. Cancers 2015, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D. Anti-angiogenesis in neuroblastoma. Crit. Rev. Oncol. Hematol. 2013, 86, 212–221. [Google Scholar] [CrossRef]
- Kaneko, S.; Ishibashi, M.; Kaneko, M. Vascular endothelial growth factor expression is closely related to irinotecan-mediated inhibition of tumor growth and angiogenesis in neuroblastoma xenografts. Cancer Sci. 2008, 99, 1209–1217. [Google Scholar] [CrossRef]
- Zhang, L.; Smith, K.M.; Chong, A.L.; Stempak, D.; Yeger, H.; Marrano, P.; Thorner, P.S.; Irwin, M.S.; Kaplan, D.R.; Baruchel, S. In Vivo Antitumor and Antimetastatic Activity of Sunitinib in Preclinical Neuroblastoma Mouse Model. Neoplasia 2009, 11, 426–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Sun, J.D.; Zhang, L.; Mokhtari, R.B.; Wu, B.; Meng, F.; Liu, Q.; Bhupathi, D.; Wang, Y.; Yeger, H.; et al. Hypoxia-Targeting Drug Evofosfamide (TH-302) Enhances Sunitinib Activity in Neuroblastoma Xenograft Models. Transl. Oncol. 2018, 11, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Hartwich, J.; Orr, W.S.; Ng, C.Y.; Spence, Y.; Morton, C.; Davidoff, A.M. HIF-1α activation mediates resistance to anti-angiogenic therapy in neuroblastoma xenografts. J. Pediatr. Surg. 2013, 48, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguera, R.; Fredlund, E.; Piqueras, M.; Pietras, A.; Beckman, S.; Navarro, S.; Pahlman, S. HIF-1 and HIF-2 Are Differentially Regulated In vivo in Neuroblastoma: High HIF-1 Correlates Negatively to Advanced Clinical Stage and Tumor Vascularization. Clin. Cancer Res. 2009, 15, 7130–7136. [Google Scholar] [CrossRef] [Green Version]
- Cimmino, F.; Pezone, L.; Avitabile, M.; Acierno, G.; Andolfo, I.; Capasso, M.; Iolascon, A. Inhibition of hypoxia inducible factors combined with all-trans retinoic acid treatment enhances glial transdifferentiation of neuroblastoma cells. Sci. Rep. 2015, 5, 11158. [Google Scholar] [CrossRef] [Green Version]
- Westerlund, I.; Shi, Y.; Toskas, K.; Fell, S.M.; Li, S.; Surova, O.; Södersten, E.; Kogner, P.; Nyman, U.; Schlisio, S.; et al. Combined epigenetic and differentiation-based treatment inhibits neuroblastoma tumor growth and links HIF2α to tumor suppression. Proc. Natl. Acad. Sci. USA 2017, 114, 6137–6146. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Cellular Processes | Genes | References | |
|---|---|---|---|
| Initiation | ↑ HIF-1α, HIF-2α, SNAIL-2, SOX10, CXCR4, PDGF1-α, FZD-6, TRKAIII | [21,22,23,25,26,31] | |
| Differentiation | ↑ IGF-2, NESP55, ID2, HES-1, TH, DDC, HIF-2α, OCT4, NOTCH1, c-Kit, Vimentin, dHAND | ↓ GAP-43, CHGA | [37,38,39,42,44] |
| Cell survival | ↑ HIF-1α, HIF-2α, VEGF, NEUROPILIN 1, ADRENOMEDULLIN, IGF-1, IGF-2, RHOA, BCL-2, Survivin, Methallothioneins, p53, MDM2 | [39,54,56,58,59,60,61,68] | |
| Metastasis | ↑ VEGF, TRIO, SERPINB9, ITGA7, HGFR, TGFβ1, IL1B, MMP2, TWIST1, SNAIL1, CDH1, HIF-1α | ↓ TIMP4, CDH1, CTNNB1, FN1 | [24,39,74,75,76,77,78,79,81] |
| Malignant phenotype | ↑ LCO4A1, ENO1, HK2, PGK1, MTFP1, HILPDA, VKORC1, TPI1, HIST1HIC | [49] | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huertas-Castaño, C.; Gómez-Muñoz, M.A.; Pardal, R.; Vega, F.M. Hypoxia in the Initiation and Progression of Neuroblastoma Tumours. Int. J. Mol. Sci. 2020, 21, 39. https://doi.org/10.3390/ijms21010039
Huertas-Castaño C, Gómez-Muñoz MA, Pardal R, Vega FM. Hypoxia in the Initiation and Progression of Neuroblastoma Tumours. International Journal of Molecular Sciences. 2020; 21(1):39. https://doi.org/10.3390/ijms21010039
Chicago/Turabian StyleHuertas-Castaño, Carlos, María A. Gómez-Muñoz, Ricardo Pardal, and Francisco M. Vega. 2020. "Hypoxia in the Initiation and Progression of Neuroblastoma Tumours" International Journal of Molecular Sciences 21, no. 1: 39. https://doi.org/10.3390/ijms21010039
APA StyleHuertas-Castaño, C., Gómez-Muñoz, M. A., Pardal, R., & Vega, F. M. (2020). Hypoxia in the Initiation and Progression of Neuroblastoma Tumours. International Journal of Molecular Sciences, 21(1), 39. https://doi.org/10.3390/ijms21010039