Unconventional Myosins: How Regulation Meets Function


Abstract
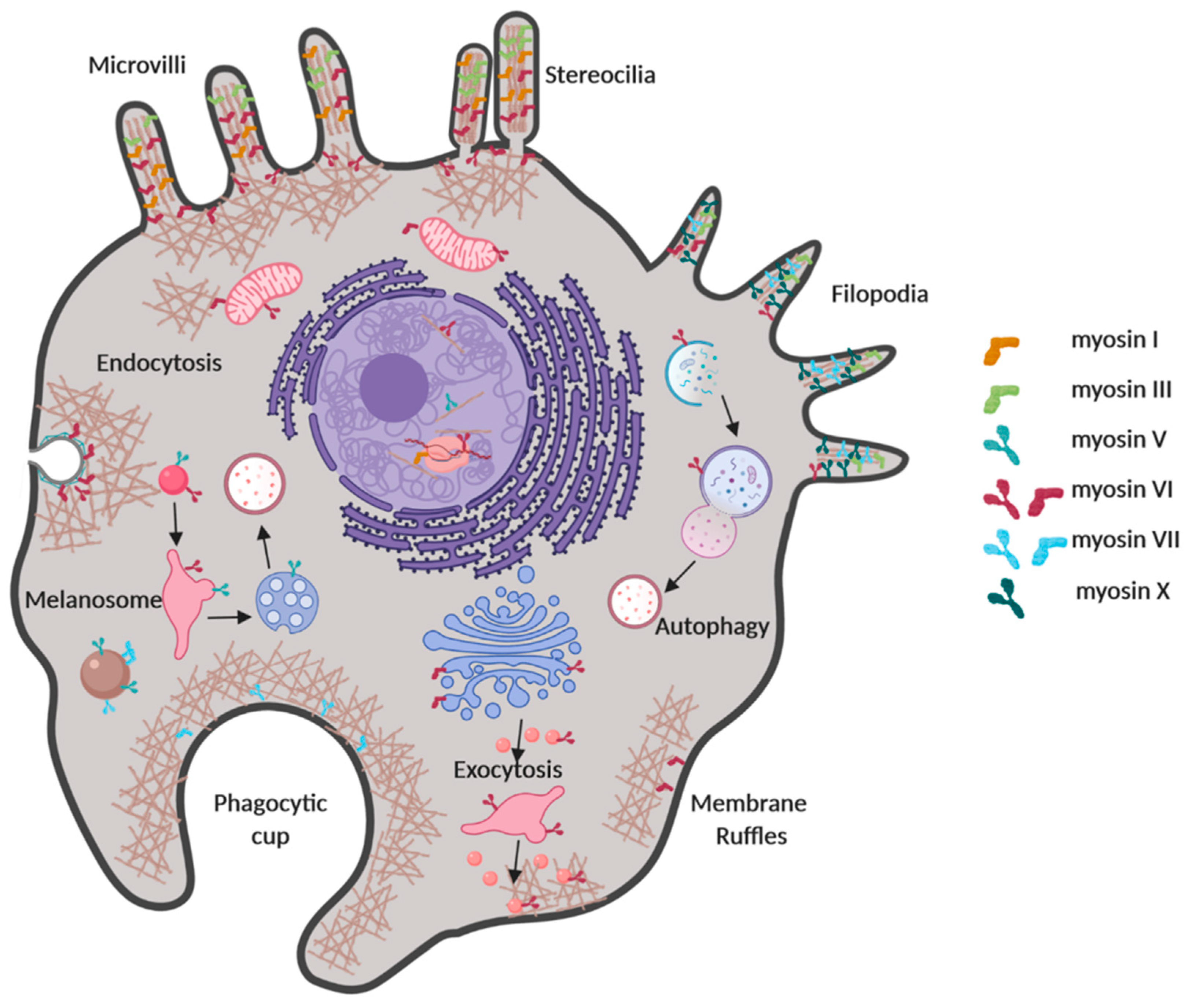
1. Introduction
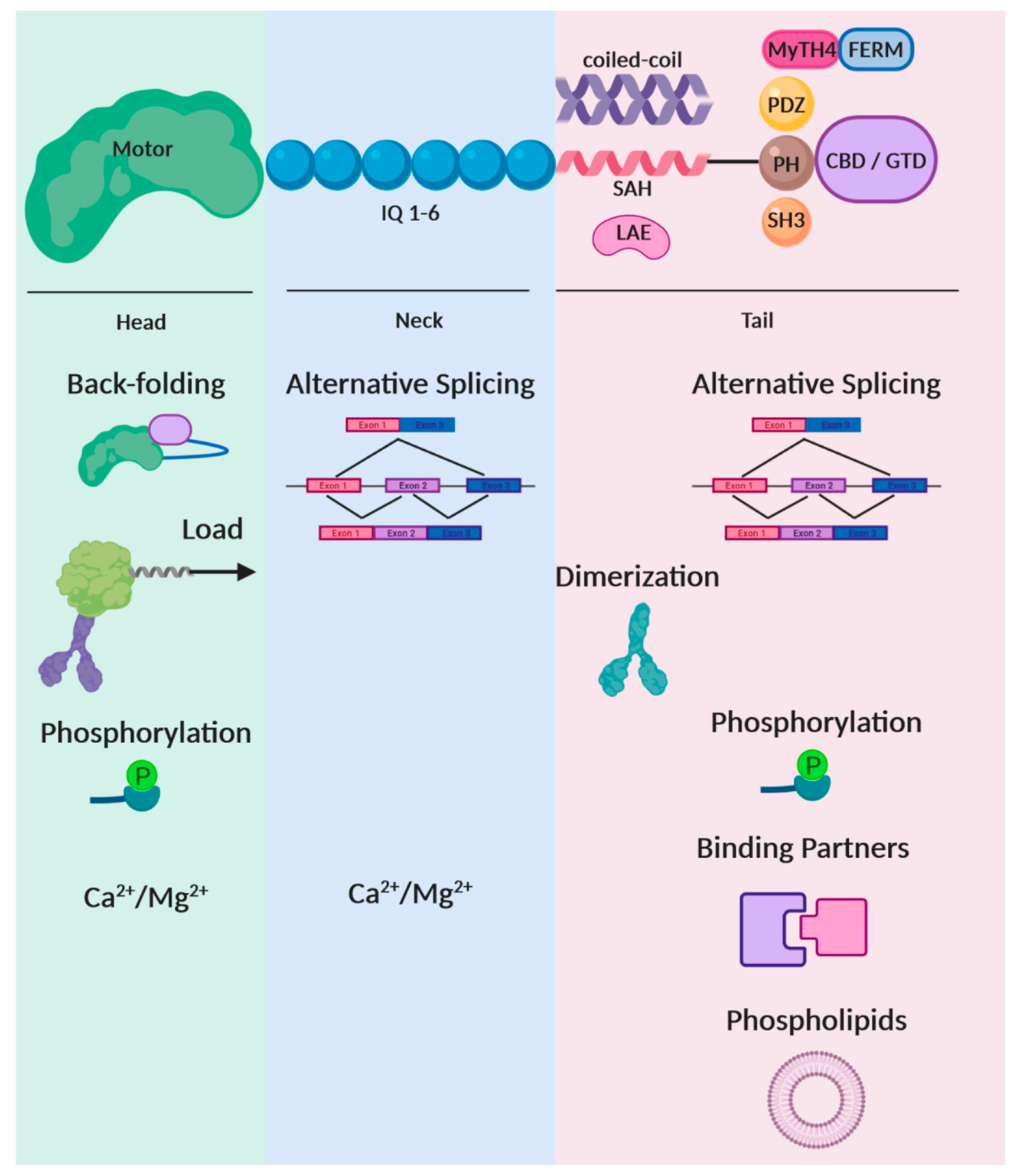
2. Regulation by Alternative Spicing
3. How the Load Upon a Myosin Modulates Its Function
4. Phosphorylation: A Versatile Regulatory Cue
5. Regulation Within the Local Environment: Divalent Cations
6. Regulation Within the Local Environment: Membranes and Phospholipids
7. Tail Dependent Regulation Through the Interaction with Binding Partners
7.1. Regulation by Binding Partners: Myosin V
7.2. Regulation by Binding Partners: Myosin VII
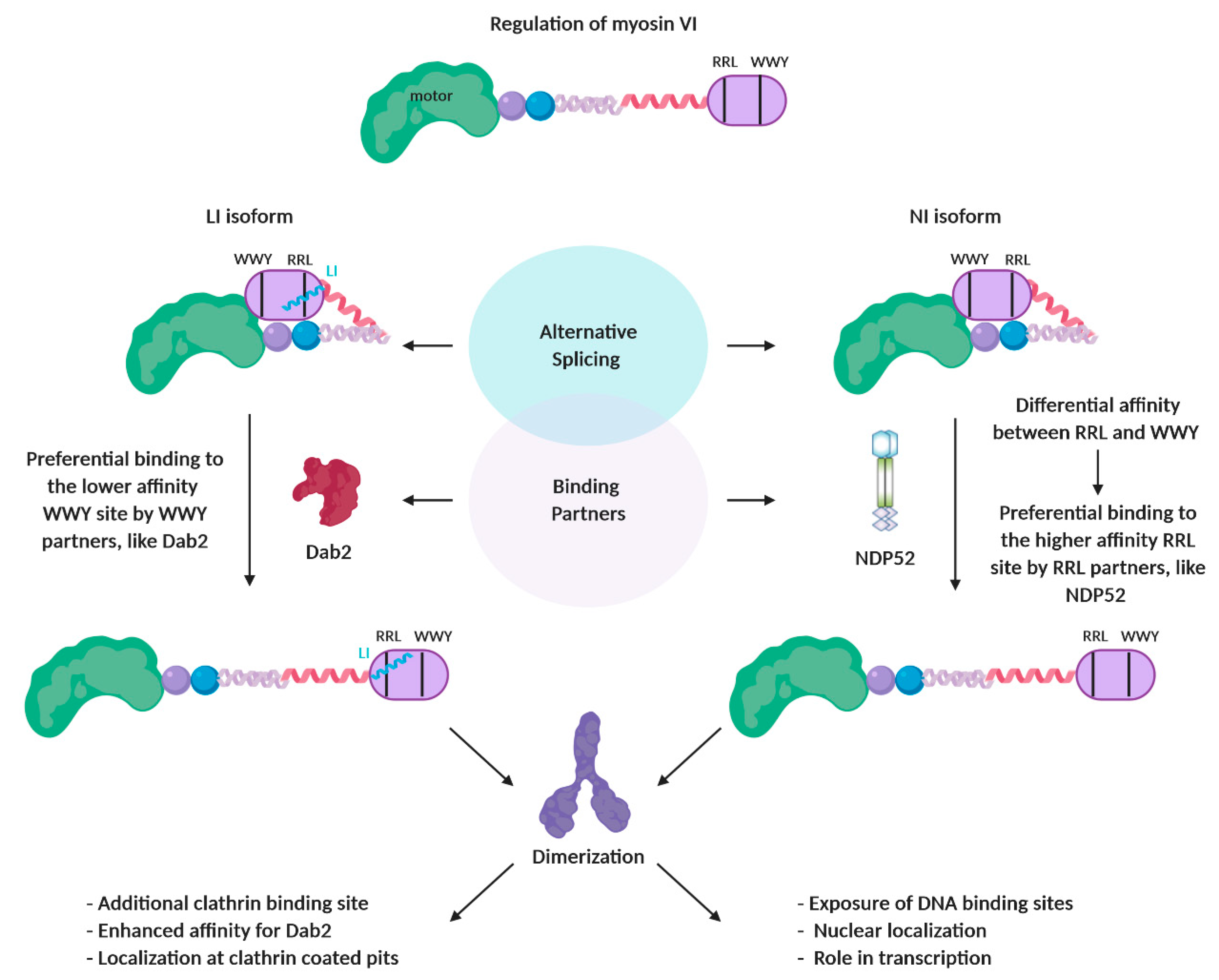
7.3. Regulation by Binding Partners: Myosin VI
8. Conclusion and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Foth, B.J.; Goedecke, M.C.; Soldati, D. New insights into myosin evolution and classification. Proc. Natl. Acad. Sci. USA 2006, 103, 3681–3686. [Google Scholar] [CrossRef] [PubMed]
- Odronitz, F.; Kollmar, M. Drawing the tree of eukaryotic life based on the analysis of 2,269 manually annotated myosins from 328 species. Genome Biol. 2007, 8, R196. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.S.; Powell, B.C.; Cheney, R.E. A millennial myosin census. Mol. Biol. Cell 2001, 12, 780–794. [Google Scholar] [CrossRef] [PubMed]
- de Lanerolle, P.; Serebryannyy, L. Nuclear actin and myosins: Life without filaments. Nat. Cell Biol. 2011, 13, 1282–1288. [Google Scholar] [CrossRef]
- Hartman, M.A.; Finan, D.; Sivaramakrishnan, S.; Spudich, J.A. Principles of unconventional myosin function and targeting. Annu. Rev. Cell Dev. Biol. 2011, 27, 133–155. [Google Scholar] [CrossRef]
- Li, J.; Lu, Q.; Zhang, M. Structural Basis of Cargo Recognition by Unconventional Myosins in Cellular Trafficking. Traffic 2016, 17, 822–838. [Google Scholar] [CrossRef]
- Lu, Q.; Li, J.; Zhang, M. Cargo recognition and cargo-mediated regulation of unconventional myosins. Acc. Chem. Res. 2014, 47, 3061–3070. [Google Scholar] [CrossRef]
- Houdusse, A.; Sweeney, H.L. How Myosin Generates Force on Actin Filaments. Trends Biochem. Sci. 2016, 41, 989–997. [Google Scholar] [CrossRef]
- Heissler, S.M.; Sellers, J.R. Various Themes of Myosin Regulation. J. Mol. Biol. 2016, 428, 1927–1946. [Google Scholar] [CrossRef]
- Purcell, T.J.; Morris, C.; Spudich, J.A.; Sweeney, H.L. Role of the lever arm in the processive stepping of myosin V. Proc. Natl. Acad. Sci. USA 2002, 99, 14159–14164. [Google Scholar] [CrossRef]
- Heissler, S.M.; Sellers, J.R. Myosin light chains: Teaching old dogs new tricks. Bioarchitecture 2014, 4, 169–188. [Google Scholar] [CrossRef] [PubMed]
- Cheney, R.E.; O’Shea, M.K.; Heuser, J.E.; Coelho, M.V.; Wolenski, J.S.; Espreafico, E.M.; Forscher, P.; Larson, R.E.; Mooseker, M.S. Brain myosin-V is a two-headed unconventional myosin with motor activity. Cell 1993, 75, 13–23. [Google Scholar] [CrossRef]
- Lu, Q.; Ye, F.; Wei, Z.; Wen, Z.; Zhang, M. Antiparallel coiled-coil-mediated dimerization of myosin X. Proc. Natl. Acad. Sci. USA 2012, 109, 17388–17393. [Google Scholar] [CrossRef] [PubMed]
- Fili, N.; Hari-Gupta, Y.; Dos Santos, A.; Cook, A.; Poland, S.; Ameer-Beg, S.M.; Parsons, M.; Toseland, C.P. NDP52 activates nuclear myosin VI to enhance RNA polymerase II transcription. Nat. Commun. 2017, 8, 1871. [Google Scholar] [CrossRef]
- Mukherjea, M.; Llinas, P.; Kim, H.; Travaglia, M.; Safer, D.; Menetrey, J.; Franzini-Armstrong, C.; Selvin, P.R.; Houdusse, A.; Sweeney, H.L. Myosin VI dimerization triggers an unfolding of a three-helix bundle in order to extend its reach. Mol. Cell 2009, 35, 305–315. [Google Scholar] [CrossRef]
- Spink, B.J.; Sivaramakrishnan, S.; Lipfert, J.; Doniach, S.; Spudich, J.A. Long single alpha-helical tail domains bridge the gap between structure and function of myosin VI. Nat. Struct. Mol. Biol. 2008, 15, 591–597. [Google Scholar] [CrossRef]
- Sakai, T.; Umeki, N.; Ikebe, R.; Ikebe, M. Cargo binding activates myosin VIIA motor function in cells. Proc. Natl. Acad. Sci. USA 2011, 108, 7028–7033. [Google Scholar] [CrossRef]
- Knight, P.J.; Thirumurugan, K.; Xu, Y.; Wang, F.; Kalverda, A.P.; Stafford, W.F.; Sellers, J.R.; Peckham, M. The predicted coiled-coil domain of myosin 10 forms a novel elongated domain that lengthens the head. J. Biol. Chem. 2005, 280, 34702–34708. [Google Scholar] [CrossRef]
- Liu, J.; Taylor, D.W.; Krementsova, E.B.; Trybus, K.M.; Taylor, K.A. Three-dimensional structure of the myosin V inhibited state by cryoelectron tomography. Nature 2006, 442, 208–211. [Google Scholar] [CrossRef]
- Thirumurugan, K.; Sakamoto, T.; Hammer, J.A.; Sellers, J.R.; Knight, P.J. The cargo-binding domain regulates structure and activity of myosin 5. Nature 2006, 442, 212–215. [Google Scholar] [CrossRef]
- Wang, F.; Thirumurugan, K.; Stafford, W.F.; Hammer, J.A.; Knight, P.J.; Sellers, J.R. Regulated conformation of myosin V. J. Biol. Chem. 2004, 279, 2333–2336. [Google Scholar] [CrossRef] [PubMed]
- Sato, O.; Li, X.D.; Ikebe, M. Myosin Va becomes a low duty ratio motor in the inhibited form. J. Biol. Chem. 2007, 282, 13228–13239. [Google Scholar] [CrossRef] [PubMed]
- Umeki, N.; Jung, H.S.; Watanabe, S.; Sakai, T.; Li, X.D.; Ikebe, R.; Craig, R.; Ikebe, M. The tail binds to the head-neck domain, inhibiting ATPase activity of myosin VIIA. Proc. Natl. Acad. Sci. USA 2009, 106, 8483–8488. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Baboolal, T.G.; Siththanandan, V.; Chen, M.; Walker, M.L.; Knight, P.J.; Peckham, M.; Sellers, J.R. A FERM domain autoregulates Drosophila myosin 7a activity. Proc. Natl. Acad. Sci. USA 2009, 106, 4189–4194. [Google Scholar] [CrossRef] [PubMed]
- Umeki, N.; Jung, H.S.; Sakai, T.; Sato, O.; Ikebe, R.; Ikebe, M. Phospholipid-dependent regulation of the motor activity of myosin X. Nat. Struct. Mol. Biol. 2011, 18, 783–788. [Google Scholar] [CrossRef] [PubMed]
- Buss, F.; Arden, S.D.; Lindsay, M.; Luzio, J.P.; Kendrick-Jones, J. Myosin VI isoform localized to clathrin-coated vesicles with a role in clathrin-mediated endocytosis. EMBO J. 2001, 20, 3676–3684. [Google Scholar] [CrossRef] [PubMed]
- Tomatis, V.M.; Papadopulos, A.; Malintan, N.T.; Martin, S.; Wallis, T.; Gormal, R.S.; Kendrick-Jones, J.; Buss, F.; Meunier, F.A. Myosin VI small insert isoform maintains exocytosis by tethering secretory granules to the cortical actin. J. Cell Biol. 2013, 200, 301–320. [Google Scholar] [CrossRef]
- Dance, A.L.; Miller, M.; Seragaki, S.; Aryal, P.; White, B.; Aschenbrenner, L.; Hasson, T. Regulation of myosin-VI targeting to endocytic compartments. Traffic 2004, 5, 798–813. [Google Scholar] [CrossRef]
- Au, J.S.; Puri, C.; Ihrke, G.; Kendrick-Jones, J.; Buss, F. Myosin VI is required for sorting of AP-1B-dependent cargo to the basolateral domain in polarized MDCK cells. J. Cell Biol. 2007, 177, 103–114. [Google Scholar] [CrossRef]
- Wollscheid, H.P.; Biancospino, M.; He, F.; Magistrati, E.; Molteni, E.; Lupia, M.; Soffientini, P.; Rottner, K.; Cavallaro, U.; Pozzoli, U.; et al. Diverse functions of myosin VI elucidated by an isoform-specific alpha-helix domain. Nat. Struct. Mol. Biol. 2016, 23, 300–308. [Google Scholar] [CrossRef]
- Fili, N.; Hari-Gupta, Y.; Aston, B.; Dos Santos, A.; Gough, R.E.; Alamad, B.; Wang, L.; Martin-Fernandez, M.L.; Toseland, C.P. Competition between two high- and low-affinity protein-binding sites in myosin VI controls its cellular function. J. Biol. Chem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Spudich, G.; Chibalina, M.V.; Au, J.S.; Arden, S.D.; Buss, F.; Kendrick-Jones, J. Myosin VI targeting to clathrin-coated structures and dimerization is mediated by binding to Disabled-2 and PtdIns (4,5) P2. Nat. Cell Biol. 2007, 9, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.D.; Mermall, V.; Strobel, M.C.; Russell, L.B.; Mooseker, M.S.; Copeland, N.G.; Jenkins, N.A. Molecular genetic dissection of mouse unconventional myosin-VA: Tail region mutations. Genetics 1998, 148, 1963–1972. [Google Scholar] [PubMed]
- Lambert, J.; Naeyaert, J.M.; Callens, T.; De Paepe, A.; Messiaen, L. Human myosin V gene produces different transcripts in a cell type-specific manner. Biochem. Biophys. Res. Commun. 1998, 252, 329–333. [Google Scholar] [CrossRef]
- Mercer, J.A.; Seperack, P.K.; Strobel, M.C.; Copeland, N.G.; Jenkins, N.A. Novel myosin heavy chain encoded by murine dilute coat colour locus. Nature 1991, 349, 709–713. [Google Scholar] [CrossRef]
- Seperack, P.K.; Mercer, J.A.; Strobel, M.C.; Copeland, N.G.; Jenkins, N.A. Retroviral sequences located within an intron of the dilute gene alter dilute expression in a tissue-specific manner. EMBO J. 1995, 14, 2326–2332. [Google Scholar] [CrossRef]
- Roland, J.T.; Lapierre, L.A.; Goldenring, J.R. Alternative splicing in class V myosins determines association with Rab10. J. Biol. Chem. 2009, 284, 1213–1223. [Google Scholar] [CrossRef]
- Au, J.S.; Huang, J.D. A tissue-specific exon of myosin Va is responsible for selective cargo binding in melanocytes. Cell Motil. Cytoskelet. 2002, 53, 89–102. [Google Scholar] [CrossRef]
- Wu, X.S.; Rao, K.; Zhang, H.; Wang, F.; Sellers, J.R.; Matesic, L.E.; Copeland, N.G.; Jenkins, N.A.; Hammer, J.A. Identification of an organelle receptor for myosin-Va. Nat. Cell Biol. 2002, 4, 271–278. [Google Scholar] [CrossRef]
- Li, X.D.; Ikebe, R.; Ikebe, M. Activation of myosin Va function by melanophilin, a specific docking partner of myosin Va. J. Biol. Chem. 2005, 280, 17815–17822. [Google Scholar] [CrossRef]
- Hodi, Z.; Nemeth, A.L.; Radnai, L.; Hetenyi, C.; Schlett, K.; Bodor, A.; Perczel, A.; Nyitray, L. Alternatively spliced exon B of myosin Va is essential for binding the tail-associated light chain shared by dynein. Biochemistry 2006, 45, 12582–12595. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.; Fodor, E.; Ginsburg, A.; Hammer, J.A. The binding of DYNLL2 to myosin Va requires alternatively spliced exon B and stabilizes a portion of the myosin’s coiled-coil domain. Biochemistry 2006, 45, 11564–11577. [Google Scholar] [CrossRef] [PubMed]
- Ruppert, C.; Kroschewski, R.; Bahler, M. Identification, characterization and cloning of myr 1, a mammalian myosin-I. J. Cell Biol. 1993, 120, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Tang, N.; Ostap, E.M. Biochemical and motile properties of Myo1b splice isoforms. J. Biol. Chem. 2005, 280, 41562–41567. [Google Scholar] [CrossRef]
- Laakso, J.M.; Lewis, J.H.; Shuman, H.; Ostap, E.M. Control of myosin-I force sensing by alternative splicing. Proc. Natl. Acad. Sci. USA 2010, 107, 698–702. [Google Scholar] [CrossRef]
- Mori, K.; Furusawa, T.; Okubo, T.; Inoue, T.; Ikawa, S.; Yanai, N.; Mori, K.J.; Obinata, M. Genome structure and differential expression of two isoforms of a novel PDZ-containing myosin (MysPDZ) (Myo18A). J. Biochem. 2003, 133, 405–413. [Google Scholar] [CrossRef]
- Dose, A.C.; Hillman, D.W.; Wong, C.; Sohlberg, L.; Lin-Jones, J.; Burnside, B. Myo3A, one of two class III myosin genes expressed in vertebrate retina, is localized to the calycal processes of rod and cone photoreceptors and is expressed in the sacculus. Mol. Biol. Cell 2003, 14, 1058–1073. [Google Scholar] [CrossRef]
- Nal, N.; Ahmed, Z.M.; Erkal, E.; Alper, O.M.; Luleci, G.; Dinc, O.; Waryah, A.M.; Ain, Q.; Tasneem, S.; Husnain, T.; et al. Mutational spectrum of MYO15A: The large N-terminal extension of myosin XVA is required for hearing. Hum. Mutat. 2007, 28, 1014–1019. [Google Scholar] [CrossRef]
- Laakso, J.M.; Lewis, J.H.; Shuman, H.; Ostap, E.M. Myosin I can act as a molecular force sensor. Science 2008, 321, 133–136. [Google Scholar] [CrossRef]
- Veigel, C.; Coluccio, L.M.; Jontes, J.D.; Sparrow, J.C.; Milligan, R.A.; Molloy, J.E. The motor protein myosin-I produces its working stroke in two steps. Nature 1999, 398, 530–533. [Google Scholar] [CrossRef]
- Batters, C.; Arthur, C.P.; Lin, A.; Porter, J.; Geeves, M.A.; Milligan, R.A.; Molloy, J.E.; Coluccio, L.M. Myo1c is designed for the adaptation response in the inner ear. EMBO J. 2004, 23, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Li, J.; Ye, F.; Zhang, M. Structure of myosin-1c tail bound to calmodulin provides insights into calcium-mediated conformational coupling. Nat. Struct. Mol. Biol. 2015, 22, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Nambiar, R.; McConnell, R.E.; Tyska, M.J. Control of cell membrane tension by myosin-I. Proc. Natl. Acad. Sci. USA 2009, 106, 11972–11977. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.A.; Yee, A.G.; Gillespie, P.G.; Corey, D.P. Localization of myosin-Ibeta near both ends of tip links in frog saccular hair cells. J. Neurosci. 1998, 18, 8637–8647. [Google Scholar] [CrossRef] [PubMed]
- Holt, J.R.; Gillespie, S.K.; Provance, D.W.; Shah, K.; Shokat, K.M.; Corey, D.P.; Mercer, J.A.; Gillespie, P.G. A chemical-genetic strategy implicates myosin-1c in adaptation by hair cells. Cell 2002, 108, 371–381. [Google Scholar] [CrossRef]
- Stauffer, E.A.; Scarborough, J.D.; Hirono, M.; Miller, E.D.; Shah, K.; Mercer, J.A.; Holt, J.R.; Gillespie, P.G. Fast adaptation in vestibular hair cells requires myosin-1c activity. Neuron 2005, 47, 541–553. [Google Scholar] [CrossRef]
- Bose, A.; Robida, S.; Furcinitti, P.S.; Chawla, A.; Fogarty, K.; Corvera, S.; Czech, M.P. Unconventional myosin Myo1c promotes membrane fusion in a regulated exocytic pathway. Mol. Cell Biol. 2004, 24, 5447–5458. [Google Scholar] [CrossRef]
- Altman, D.; Sweeney, H.L.; Spudich, J.A. The mechanism of myosin VI translocation and its load-induced anchoring. Cell 2004, 116, 737–749. [Google Scholar] [CrossRef]
- Cramer, L.P. Myosin VI: Roles for a minus end-directed actin motor in cells. J. Cell Biol. 2000, 150, F121–F126. [Google Scholar] [CrossRef]
- Self, T.; Sobe, T.; Copeland, N.G.; Jenkins, N.A.; Avraham, K.B.; Steel, K.P. Role of myosin VI in the differentiation of cochlear hair cells. Dev. Biol. 1999, 214, 331–341. [Google Scholar] [CrossRef]
- Seiler, C.; Ben-David, O.; Sidi, S.; Hendrich, O.; Rusch, A.; Burnside, B.; Avraham, K.B.; Nicolson, T. Myosin VI is required for structural integrity of the apical surface of sensory hair cells in zebrafish. Dev. Biol. 2004, 272, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Lenartowska, M.; Miller, K.G. Myosin VI stabilizes an actin network during Drosophila spermatid individualization. Mol. Biol. Cell 2006, 17, 2559–2571. [Google Scholar] [CrossRef] [PubMed]
- Hicks, J.L.; Deng, W.M.; Rogat, A.D.; Miller, K.G.; Bownes, M. Class VI unconventional myosin is required for spermatogenesis in Drosophila. Mol. Biol. Cell 1999, 10, 4341–4353. [Google Scholar] [CrossRef] [PubMed]
- Naccache, S.N.; Hasson, T. Myosin VI altered at threonine 406 stabilizes actin filaments in vivo. Cell Motil. Cytoskelet. 2006, 63, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Maddugoda, M.P.; Crampton, M.S.; Shewan, A.M.; Yap, A.S. Myosin VI and vinculin cooperate during the morphogenesis of cadherin cell cell contacts in mammalian epithelial cells. J. Cell Biol. 2007, 178, 529–540. [Google Scholar] [CrossRef]
- Pathak, D.; Sepp, K.J.; Hollenbeck, P.J. Evidence that myosin activity opposes microtubule-based axonal transport of mitochondria. J. Neurosci. 2010, 30, 8984–8992. [Google Scholar] [CrossRef]
- Kruppa, A.J.; Kishi-Itakura, C.; Masters, T.A.; Rorbach, J.E.; Grice, G.L.; Kendrick-Jones, J.; Nathan, J.A.; Minczuk, M.; Buss, F. Myosin VI-Dependent Actin Cages Encapsulate Parkin-Positive Damaged Mitochondria. Dev. Cell 2018, 44, 484–499. [Google Scholar] [CrossRef]
- Desnos, C.; Huet, S.; Fanget, I.; Chapuis, C.; Bottiger, C.; Racine, V.; Sibarita, J.B.; Henry, J.P.; Darchen, F. Myosin Va mediates docking of secretory granules at the plasma membrane. J. Neurosci. 2007, 27, 10636–10645. [Google Scholar] [CrossRef]
- Levi, V.; Gelfand, V.I.; Serpinskaya, A.S.; Gratton, E. Melanosomes transported by myosin-V in Xenopus melanophores perform slow 35 nm steps. Biophys. J. 2006, 90, L07–L09. [Google Scholar] [CrossRef][Green Version]
- Provance, D.W., Jr.; Addison, E.J.; Wood, P.R.; Chen, D.Z.; Silan, C.M.; Mercer, J.A. Myosin-Vb functions as a dynamic tether for peripheral endocytic compartments during transferrin trafficking. BMC Cell Biol. 2008, 9, 44. [Google Scholar] [CrossRef]
- Wu, X.; Bowers, B.; Rao, K.; Wei, Q.; Hammer, J.A. Visualization of melanosome dynamics within wild-type and dilute melanocytes suggests a paradigm for myosin V function in vivo. J. Cell Biol. 1998, 143, 1899–1918. [Google Scholar] [CrossRef] [PubMed]
- Bement, W.M.; Mooseker, M.S. TEDS rule: A molecular rationale for differential regulation of myosins by phosphorylation of the heavy chain head. Cell Motil. Cytoskelet. 1995, 31, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Albanesi, J.P.; Hammer, J.A.; Korn, E.D. The interaction of F-actin with phosphorylated and unphosphorylated myosins IA and IB from Acanthamoeba castellanii. J. Biol. Chem. 1983, 258, 10176–10181. [Google Scholar] [PubMed]
- Brzeska, H.; Knaus, U.G.; Wang, Z.Y.; Bokoch, G.M.; Korn, E.D. p21-activated kinase has substrate specificity similar to Acanthamoeba myosin I heavy chain kinase and activates Acanthamoeba myosin I. Proc. Natl. Acad. Sci. USA 1997, 94, 1092–1095. [Google Scholar] [CrossRef] [PubMed]
- Brzeska, H.; Lynch, T.J.; Martin, B.; Korn, E.D. The localization and sequence of the phosphorylation sites of Acanthamoeba myosins I. An improved method for locating the phosphorylated amino acid. J. Biol. Chem. 1989, 264, 19340–19348. [Google Scholar]
- Durrwang, U.; Fujita-Becker, S.; Erent, M.; Kull, F.J.; Tsiavaliaris, G.; Geeves, M.A.; Manstein, D.J. Dictyostelium myosin-IE is a fast molecular motor involved in phagocytosis. J. Cell Sci. 2006, 119, 550–558. [Google Scholar] [CrossRef]
- Fujita-Becker, S.; Durrwang, U.; Erent, M.; Clark, R.J.; Geeves, M.A.; Manstein, D.J. Changes in Mg2+ ion concentration and heavy chain phosphorylation regulate the motor activity of a class I myosin. J. Biol. Chem. 2005, 280, 6064–6071. [Google Scholar] [CrossRef]
- Grosshans, B.L.; Grotsch, H.; Mukhopadhyay, D.; Fernandez, I.M.; Pfannstiel, J.; Idrissi, F.Z.; Lechner, J.; Riezman, H.; Geli, M.I. TEDS site phosphorylation of the yeast myosins I is required for ligand-induced but not for constitutive endocytosis of the G protein-coupled receptor Ste2p. J. Biol. Chem. 2006, 281, 11104–11114. [Google Scholar] [CrossRef]
- Attanapola, S.L.; Alexander, C.J.; Mulvihill, D.P. Ste20-kinase-dependent TEDS-site phosphorylation modulates the dynamic localisation and endocytic function of the fission yeast class I myosin, Myo1. J. Cell Sci. 2009, 122, 3856–3861. [Google Scholar] [CrossRef]
- Yoshimura, M.; Homma, K.; Saito, J.; Inoue, A.; Ikebe, R.; Ikebe, M. Dual regulation of mammalian myosin VI motor function. J. Biol. Chem. 2001, 276, 39600–39607. [Google Scholar] [CrossRef]
- Buss, F.; Kendrick-Jones, J.; Lionne, C.; Knight, A.E.; Cote, G.P.; Paul Luzio, J. The localization of myosin VI at the golgi complex and leading edge of fibroblasts and its phosphorylation and recruitment into membrane ruffles of A431 cells after growth factor stimulation. J. Cell Biol. 1998, 143, 1535–1545. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.A.; Wells, A.L.; Yang, Z.; Chen, L.Q.; Baldacchino, C.V.; Sweeney, H.L. Calcium functionally uncouples the heads of myosin VI. J. Biol. Chem. 2003, 278, 23324–23330. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.B.; Humphreys, D.; Singh, V.; Davidson, A.C.; Arden, S.D.; Buss, F.; Koronakis, V. MYO6 is targeted by Salmonella virulence effectors to trigger PI3-kinase signaling and pathogen invasion into host cells. Proc. Natl. Acad. Sci. USA 2017, 114, 3915–3920. [Google Scholar] [CrossRef] [PubMed]
- De La Cruz, E.M.; Ostap, E.M.; Sweeney, H.L. Kinetic mechanism and regulation of myosin VI. J. Biol. Chem. 2001, 276, 32373–32381. [Google Scholar] [CrossRef]
- Schneider, M.E.; Dose, A.C.; Salles, F.T.; Chang, W.; Erickson, F.L.; Burnside, B.; Kachar, B. A new compartment at stereocilia tips defined by spatial and temporal patterns of myosin IIIa expression. J. Neurosci. 2006, 26, 10243–10252. [Google Scholar] [CrossRef]
- Dose, A.C.; Burnside, B. Cloning and chromosomal localization of a human class III myosin. Genomics 2000, 67, 333–342. [Google Scholar] [CrossRef]
- Cardasis, H.L.; Stevens, S.M., Jr.; McClung, S.; Kempler, K.E.; Powell, D.H.; Eyler, J.R.; Battelle, B.A. The actin-binding interface of a myosin III is phosphorylated in vivo in response to signals from a circadian clock. Biochemistry 2007, 46, 13907–13919. [Google Scholar] [CrossRef]
- Dalal, J.S.; Stevens, S.M., Jr.; Alvarez, S.; Munoz, N.; Kempler, K.E.; Dose, A.C.; Burnside, B.; Battelle, B.A. Mouse class III myosins: Kinase activity and phosphorylation sites. J. Neurochem. 2011, 119, 772–784. [Google Scholar] [CrossRef]
- Kempler, K.; Toth, J.; Yamashita, R.; Mapel, G.; Robinson, K.; Cardasis, H.; Stevens, S.; Sellers, J.R.; Battelle, B.A. Loop 2 of limulus myosin III is phosphorylated by protein kinase A and autophosphorylation. Biochemistry 2007, 46, 4280–4293. [Google Scholar] [CrossRef]
- Dose, A.C.; Ananthanarayanan, S.; Moore, J.E.; Burnside, B.; Yengo, C.M. Kinetic mechanism of human myosin IIIA. J. Biol. Chem. 2007, 282, 216–231. [Google Scholar] [CrossRef]
- Komaba, S.; Watanabe, S.; Umeki, N.; Sato, O.; Ikebe, M. Effect of phosphorylation in the motor domain of human myosin IIIA on its ATP hydrolysis cycle. Biochemistry 2010, 49, 3695–3702. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Quintero, O.A.; Moore, J.E.; Unrath, W.C.; Manor, U.; Salles, F.T.; Grati, M.; Kachar, B.; Yengo, C.M. Intermolecular autophosphorylation regulates myosin IIIa activity and localization in parallel actin bundles. J. Biol. Chem. 2010, 285, 35770–35782. [Google Scholar] [CrossRef] [PubMed]
- An, B.C.; Sakai, T.; Komaba, S.; Kishi, H.; Kobayashi, S.; Kim, J.Y.; Ikebe, R.; Ikebe, M. Phosphorylation of the kinase domain regulates autophosphorylation of myosin IIIA and its translocation in microvilli. Biochemistry 2014, 53, 7835–7845. [Google Scholar] [CrossRef] [PubMed]
- Yoshizaki, T.; Imamura, T.; Babendure, J.L.; Lu, J.C.; Sonoda, N.; Olefsky, J.M. Myosin 5a is an insulin-stimulated Akt2 (protein kinase Bbeta) substrate modulating GLUT4 vesicle translocation. Mol. Cell Biol. 2007, 27, 5172–5183. [Google Scholar] [CrossRef]
- Karcher, R.L.; Roland, J.T.; Zappacosta, F.; Huddleston, M.J.; Annan, R.S.; Carr, S.A.; Gelfand, V.I. Cell cycle regulation of myosin-V by calcium/calmodulin-dependent protein kinase II. Science 2001, 293, 1317–1320. [Google Scholar] [CrossRef]
- Pranchevicius, M.C.; Baqui, M.M.; Ishikawa-Ankerhold, H.C.; Lourenco, E.V.; Leao, R.M.; Banzi, S.R.; dos Santos, C.T.; Roque-Barreira, M.C.; Espreafico, E.M.; Larson, R.E. Myosin Va phosphorylated on Ser1650 is found in nuclear speckles and redistributes to nucleoli upon inhibition of transcription. Cell Motil. Cytoskelet. 2008, 65, 441–456. [Google Scholar] [CrossRef]
- Fernandez-Golbano, I.M.; Idrissi, F.Z.; Giblin, J.P.; Grosshans, B.L.; Robles, V.; Grotsch, H.; Borras Mdel, M.; Geli, M.I. Crosstalk between PI (4,5) P (2) and CK2 modulates actin polymerization during endocytic uptake. Dev. Cell 2014, 30, 746–758. [Google Scholar] [CrossRef]
- Sahlender, D.A.; Roberts, R.C.; Arden, S.D.; Spudich, G.; Taylor, M.J.; Luzio, J.P.; Kendrick-Jones, J.; Buss, F. Optineurin links myosin VI to the Golgi complex and is involved in Golgi organization and exocytosis. J. Cell Biol. 2005, 169, 285–295. [Google Scholar] [CrossRef]
- McLeish, K.R.; Merchant, M.L.; Creed, T.M.; Tandon, S.; Barati, M.T.; Uriarte, S.M.; Ward, R.A. Frontline Science: Tumor necrosis factor-alpha stimulation and priming of human neutrophil granule exocytosis. J. Leukoc. Biol. 2017, 102, 19–29. [Google Scholar] [CrossRef]
- Rosenfeld, S.S.; Houdusse, A.; Sweeney, H.L. Magnesium regulates ADP dissociation from myosin V. J. Biol. Chem. 2005, 280, 6072–6079. [Google Scholar] [CrossRef]
- Swenson, A.M.; Trivedi, D.V.; Rauscher, A.A.; Wang, Y.; Takagi, Y.; Palmer, B.M.; Malnasi-Csizmadia, A.; Debold, E.P.; Yengo, C.M. Magnesium modulates actin binding and ADP release in myosin motors. J. Biol. Chem. 2014, 289, 23977–23991. [Google Scholar] [CrossRef] [PubMed]
- Heissler, S.M.; Selvadurai, J.; Bond, L.M.; Fedorov, R.; Kendrick-Jones, J.; Buss, F.; Manstein, D.J. Kinetic properties and small-molecule inhibition of human myosin-6. FEBS Lett. 2012, 586, 3208–3214. [Google Scholar] [CrossRef] [PubMed]
- Heissler, S.M.; Manstein, D.J. Comparative kinetic and functional characterization of the motor domains of human nonmuscle myosin-2C isoforms. J. Biol. Chem. 2011, 286, 21191–21202. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, D.V.; Muretta, J.M.; Swenson, A.M.; Thomas, D.D.; Yengo, C.M. Magnesium impacts myosin V motor activity by altering key conformational changes in the mechanochemical cycle. Biochemistry 2013, 52, 4710–4722. [Google Scholar] [CrossRef]
- Adamek, N.; Coluccio, L.M.; Geeves, M.A. Calcium sensitivity of the cross-bridge cycle of Myo1c, the adaptation motor in the inner ear. Proc. Natl. Acad. Sci. USA 2008, 105, 5710–5715. [Google Scholar] [CrossRef]
- Coluccio, L.M.; Geeves, M.A. Transient kinetic analysis of the 130-kDa myosin I (MYR-1 gene product) from rat liver. A myosin I designed for maintenance of tension? J. Biol. Chem. 1999, 274, 21575–21580. [Google Scholar] [CrossRef]
- Geeves, M.A.; Perreault-Micale, C.; Coluccio, L.M. Kinetic analyses of a truncated mammalian myosin I suggest a novel isomerization event preceding nucleotide binding. J. Biol. Chem. 2000, 275, 21624–21630. [Google Scholar] [CrossRef]
- Munnich, S.; Taft, M.H.; Manstein, D.J. Crystal structure of human myosin 1c—The motor in GLUT4 exocytosis: Implications for Ca2+ regulation and 14-3-3 binding. J. Mol. Biol. 2014, 426, 2070–2081. [Google Scholar] [CrossRef]
- Shuman, H.; Greenberg, M.J.; Zwolak, A.; Lin, T.; Sindelar, C.V.; Dominguez, R.; Ostap, E.M. A vertebrate myosin-I structure reveals unique insights into myosin mechanochemical tuning. Proc. Natl. Acad. Sci. USA 2014, 111, 2116–2121. [Google Scholar] [CrossRef]
- McConnell, R.E.; Tyska, M.J. Leveraging the membrane - cytoskeleton interface with myosin-1. Trends Cell Biol. 2010, 20, 418–426. [Google Scholar] [CrossRef]
- Li, J.; Chen, Y.; Deng, Y.; Unarta, I.C.; Lu, Q.; Huang, X.; Zhang, M. Ca2+-Induced Rigidity Change of the Myosin VIIa IQ Motif-Single alpha Helix Lever Arm Extension. Structure 2017, 25, 579–591.e4. [Google Scholar] [CrossRef] [PubMed]
- Li, X.D.; Mabuchi, K.; Ikebe, R.; Ikebe, M. Ca2+-induced activation of ATPase activity of myosin Va is accompanied with a large conformational change. Biochem. Biophys. Res. Commun. 2004, 315, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Li, X.D.; Jung, H.S.; Mabuchi, K.; Craig, R.; Ikebe, M. The globular tail domain of myosin Va functions as an inhibitor of the myosin Va motor. J. Biol. Chem. 2006, 281, 21789–21798. [Google Scholar] [CrossRef] [PubMed]
- Li, X.D.; Jung, H.S.; Wang, Q.; Ikebe, R.; Craig, R.; Ikebe, M. The globular tail domain puts on the brake to stop the ATPase cycle of myosin Va. Proc. Natl. Acad. Sci. USA 2008, 105, 1140–1145. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Shen, M.; Cao, Y.; Zhang, H.M.; Yao, L.L.; Li, X.D. Calmodulin bound to the first IQ motif is responsible for calcium-dependent regulation of myosin 5a. J. Biol. Chem. 2012, 287, 16530–16540. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Zhang, N.; Zheng, S.; Zhang, W.B.; Zhang, H.M.; Lu, Z.; Su, Q.P.; Sun, Y.; Ye, K.; Li, X.D. Calmodulin in complex with the first IQ motif of myosin-5a functions as an intact calcium sensor. Proc. Natl. Acad. Sci. USA 2016, 113, E5812–E5820. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Edwards, J.G.; Riley, N.; Provance, D.W., Jr.; Karcher, R.; Li, X.D.; Davison, I.G.; Ikebe, M.; Mercer, J.A.; Kauer, J.A.; et al. Myosin Vb mobilizes recycling endosomes and AMPA receptors for postsynaptic plasticity. Cell 2008, 135, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Gardini, L.; Heissler, S.M.; Arbore, C.; Yang, Y.; Sellers, J.R.; Pavone, F.S.; Capitanio, M. Dissecting myosin-5B mechanosensitivity and calcium regulation at the single molecule level. Nat. Commun. 2018, 9, 2844. [Google Scholar] [CrossRef]
- Nguyen, H.; Higuchi, H. Motility of myosin V regulated by the dissociation of single calmodulin. Nat. Struct. Mol. Biol. 2005, 12, 127–132. [Google Scholar] [CrossRef]
- Trybus, K.M.; Gushchin, M.I.; Lui, H.; Hazelwood, L.; Krementsova, E.B.; Volkmann, N.; Hanein, D. Effect of calcium on calmodulin bound to the IQ motifs of myosin V. J. Biol. Chem. 2007, 282, 23316–23325. [Google Scholar] [CrossRef]
- Batters, C.; Brack, D.; Ellrich, H.; Averbeck, B.; Veigel, C. Calcium can mobilize and activate myosin-VI. Proc. Natl. Acad. Sci. USA 2016, 113, E1162–E1169. [Google Scholar] [CrossRef] [PubMed]
- Feeser, E.A.; Ignacio, C.M.; Krendel, M.; Ostap, E.M. Myo1e binds anionic phospholipids with high affinity. Biochemistry 2010, 49, 9353–9360. [Google Scholar] [CrossRef] [PubMed]
- Hokanson, D.E.; Laakso, J.M.; Lin, T.; Sept, D.; Ostap, E.M. Myo1c binds phosphoinositides through a putative pleckstrin homology domain. Mol. Biol. Cell 2006, 17, 4856–4865. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Yu, J.; Yan, J.; Wei, Z.; Zhang, M. Structural basis of the myosin X PH1 (N)-PH2-PH1 (C) tandem as a specific and acute cellular PI (3,4,5) P (3) sensor. Mol. Biol. Cell 2011, 22, 4268–4278. [Google Scholar] [CrossRef]
- Kerber, M.L.; Cheney, R.E. Myosin-X: A MyTH-FERM myosin at the tips of filopodia. J. Cell Sci. 2011, 124, 3733–3741. [Google Scholar] [CrossRef]
- Berg, J.S.; Derfler, B.H.; Pennisi, C.M.; Corey, D.P.; Cheney, R.E. Myosin-X, a novel myosin with pleckstrin homology domains, associates with regions of dynamic actin. J. Cell Sci. 2000, 113 Pt 19, 3439–3451. [Google Scholar]
- Yu, C.; Lou, J.; Wu, J.; Pan, L.; Feng, W.; Zhang, M. Membrane-induced lever arm expansion allows myosin VI to walk with large and variable step sizes. J. Biol. Chem. 2012, 287, 35021–35035. [Google Scholar] [CrossRef]
- McKenna, J.M.; Ostap, E.M. Kinetics of the interaction of myo1c with phosphoinositides. J. Biol. Chem. 2009, 284, 28650–28659. [Google Scholar] [CrossRef]
- Komaba, S.; Coluccio, L.M. Localization of myosin 1b to actin protrusions requires phosphoinositide binding. J. Biol. Chem. 2010, 285, 27686–27693. [Google Scholar] [CrossRef]
- Olety, B.; Walte, M.; Honnert, U.; Schillers, H.; Bahler, M. Myosin 1G (Myo1G) is a haematopoietic specific myosin that localises to the plasma membrane and regulates cell elasticity. FEBS Lett. 2010, 584, 493–499. [Google Scholar] [CrossRef]
- Patino-Lopez, G.; Aravind, L.; Dong, X.; Kruhlak, M.J.; Ostap, E.M.; Shaw, S. Myosin 1G is an abundant class I myosin in lymphocytes whose localization at the plasma membrane depends on its ancient divergent pleckstrin homology (PH) domain (Myo1PH). J. Biol. Chem. 2010, 285, 8675–8686. [Google Scholar] [CrossRef] [PubMed]
- Nevzorov, I.; Sidorenko, E.; Wang, W.; Zhao, H.; Vartiainen, M.K. Myosin-1C uses a novel phosphoinositide-dependent pathway for nuclear localization. EMBO Rep. 2018, 19, 290–304. [Google Scholar] [CrossRef] [PubMed]
- Pyrpassopoulos, S.; Feeser, E.A.; Mazerik, J.N.; Tyska, M.J.; Ostap, E.M. Membrane-bound myo1c powers asymmetric motility of actin filaments. Curr. Biol. 2012, 22, 1688–1692. [Google Scholar] [CrossRef] [PubMed]
- Pylypenko, O.; Attanda, W.; Gauquelin, C.; Lahmani, M.; Coulibaly, D.; Baron, B.; Hoos, S.; Titus, M.A.; England, P.; Houdusse, A.M. Structural basis of myosin V Rab GTPase-dependent cargo recognition. Proc. Natl. Acad. Sci. USA 2013, 110, 20443–20448. [Google Scholar] [CrossRef]
- Wei, Z.; Liu, X.; Yu, C.; Zhang, M. Structural basis of cargo recognitions for class V myosins. Proc. Natl. Acad. Sci. USA 2013, 110, 11314–11319. [Google Scholar] [CrossRef]
- Fukuda, M.; Kuroda, T.S.; Mikoshiba, K. Slac2-a/melanophilin, the missing link between Rab27 and myosin Va: Implications of a tripartite protein complex for melanosome transport. J. Biol. Chem. 2002, 277, 12432–12436. [Google Scholar] [CrossRef]
- Strom, M.; Hume, A.N.; Tarafder, A.K.; Barkagianni, E.; Seabra, M.C. A family of Rab27-binding proteins. Melanophilin links Rab27a and myosin Va function in melanosome transport. J. Biol. Chem. 2002, 277, 25423–25430. [Google Scholar] [CrossRef]
- Yao, L.L.; Cao, Q.J.; Zhang, H.M.; Zhang, J.; Cao, Y.; Li, X.D. Melanophilin Stimulates Myosin-5a Motor Function by Allosterically Inhibiting the Interaction between the Head and Tail of Myosin-5a. Sci. Rep. 2015, 5, 10874. [Google Scholar] [CrossRef]
- Wu, X.; Wang, F.; Rao, K.; Sellers, J.R.; Hammer, J.A. Rab27a is an essential component of melanosome receptor for myosin Va. Mol. Biol. Cell 2002, 13, 1735–1749. [Google Scholar] [CrossRef]
- Sckolnick, M.; Krementsova, E.B.; Warshaw, D.M.; Trybus, K.M. More than just a cargo adapter, melanophilin prolongs and slows processive runs of myosin Va. J. Biol. Chem. 2013, 288, 29313–29322. [Google Scholar] [CrossRef]
- Matsui, T.; Ohbayashi, N.; Fukuda, M. The Rab interacting lysosomal protein (RILP) homology domain functions as a novel effector domain for small GTPase Rab36: Rab36 regulates retrograde melanosome transport in melanocytes. J. Biol. Chem. 2012, 287, 28619–28631. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, O.C.; Cheney, R.E. Human myosin-Vc is a novel class V myosin expressed in epithelial cells. J. Cell Sci. 2002, 115, 991–1004. [Google Scholar] [PubMed]
- Roland, J.T.; Kenworthy, A.K.; Peranen, J.; Caplan, S.; Goldenring, J.R. Myosin Vb interacts with Rab8a on a tubular network containing EHD1 and EHD3. Mol. Biol. Cell 2007, 18, 2828–2837. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.H.; Zhang, H.M.; Shen, M.; Yao, L.L.; Li, X.D. The motor function of Drosophila melanogaster myosin-5 is activated by calcium and cargo-binding protein dRab11. Biochem. J. 2015, 469, 135–144. [Google Scholar] [CrossRef]
- Dunn, B.D.; Sakamoto, T.; Hong, M.S.; Sellers, J.R.; Takizawa, P.A. Myo4p is a monomeric myosin with motility uniquely adapted to transport mRNA. J. Cell Biol. 2007, 178, 1193–1206. [Google Scholar] [CrossRef]
- Bertrand, E.; Chartrand, P.; Schaefer, M.; Shenoy, S.M.; Singer, R.H.; Long, R.M. Localization of ASH1 mRNA particles in living yeast. Mol. Cell 1998, 2, 437–445. [Google Scholar] [CrossRef]
- Hodges, A.R.; Krementsova, E.B.; Trybus, K.M. She3p binds to the rod of yeast myosin V and prevents it from dimerizing, forming a single-headed motor complex. J. Biol. Chem. 2008, 283, 6906–6914. [Google Scholar] [CrossRef]
- Krementsova, E.B.; Hodges, A.R.; Bookwalter, C.S.; Sladewski, T.E.; Travaglia, M.; Sweeney, H.L.; Trybus, K.M. Two single-headed myosin V motors bound to a tetrameric adapter protein form a processive complex. J. Cell Biol. 2011, 195, 631–641. [Google Scholar] [CrossRef]
- Sladewski, T.E.; Bookwalter, C.S.; Hong, M.S.; Trybus, K.M. Single-molecule reconstitution of mRNA transport by a class V myosin. Nat. Struct. Mol. Biol. 2013, 20, 952–957. [Google Scholar] [CrossRef]
- El-Amraoui, A.; Schonn, J.S.; Kussel-Andermann, P.; Blanchard, S.; Desnos, C.; Henry, J.P.; Wolfrum, U.; Darchen, F.; Petit, C. MyRIP, a novel Rab effector, enables myosin VIIa recruitment to retinal melanosomes. EMBO Rep. 2002, 3, 463–470. [Google Scholar] [CrossRef]
- Sivaramakrishnan, S.; Spudich, J.A. Coupled myosin VI motors facilitate unidirectional movement on an F-actin network. J. Cell Biol. 2009, 187, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, H.L.; Houdusse, A. Myosin VI rewrites the rules for myosin motors. Cell 2010, 141, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Tumbarello, D.A.; Kendrick-Jones, J.; Buss, F. Myosin VI and its cargo adaptors-linking endocytosis and autophagy. J. Cell Sci. 2013, 126, 2561–2570. [Google Scholar] [CrossRef] [PubMed]
- Vreugde, S.; Ferrai, C.; Miluzio, A.; Hauben, E.; Marchisio, P.C.; Crippa, M.P.; Bussi, M.; Biffo, S. Nuclear myosin VI enhances RNA polymerase II-dependent transcription. Mol. Cell 2006, 23, 749–755. [Google Scholar] [CrossRef]
- Lister, I.; Schmitz, S.; Walker, M.; Trinick, J.; Buss, F.; Veigel, C.; Kendrick-Jones, J. A monomeric myosin VI with a large working stroke. EMBO J. 2004, 23, 1729–1738. [Google Scholar] [CrossRef]
- Song, C.F.; Sader, K.; White, H.; Kendrick-Jones, J.; Trinick, J. Nucleotide-dependent shape changes in the reverse direction motor, myosin VI. Biophys. J. 2010, 99, 3336–3344. [Google Scholar] [CrossRef]
- Morriswood, B.; Ryzhakov, G.; Puri, C.; Arden, S.D.; Roberts, R.; Dendrou, C.; Kendrick-Jones, J.; Buss, F. T6BP and NDP52 are myosin VI binding partners with potential roles in cytokine signalling and cell adhesion. J. Cell Sci. 2007, 120, 2574–2585. [Google Scholar] [CrossRef]
- Park, H.; Ramamurthy, B.; Travaglia, M.; Safer, D.; Chen, L.Q.; Franzini-Armstrong, C.; Selvin, P.R.; Sweeney, H.L. Full-length myosin VI dimerizes and moves processively along actin filaments upon monomer clustering. Mol. Cell 2006, 21, 331–336. [Google Scholar] [CrossRef]
- Altman, D.; Goswami, D.; Hasson, T.; Spudich, J.A.; Mayor, S. Precise positioning of myosin VI on endocytic vesicles in vivo. PLoS Biol. 2007, 5, e210. [Google Scholar] [CrossRef]
- Yu, C.; Feng, W.; Wei, Z.; Miyanoiri, Y.; Wen, W.; Zhao, Y.; Zhang, M. Myosin VI undergoes cargo-mediated dimerization. Cell 2009, 138, 537–548. [Google Scholar] [CrossRef]
- Nishikawa, S.; Homma, K.; Komori, Y.; Iwaki, M.; Wazawa, T.; Hikikoshi Iwane, A.; Saito, J.; Ikebe, R.; Katayama, E.; Yanagida, T.; et al. Class VI myosin moves processively along actin filaments backward with large steps. Biochem. Biophys. Res. Commun. 2002, 290, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Rock, R.S.; Rice, S.E.; Wells, A.L.; Purcell, T.J.; Spudich, J.A.; Sweeney, H.L. Myosin VI is a processive motor with a large step size. Proc. Natl. Acad. Sci. USA 2001, 98, 13655–13659. [Google Scholar] [CrossRef] [PubMed]
- Naccache, S.N.; Hasson, T.; Horowitz, A. Binding of internalized receptors to the PDZ domain of GIPC/synectin recruits myosin VI to endocytic vesicles. Proc. Natl. Acad. Sci. USA 2006, 103, 12735–12740. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Myosin | Mode of Regulation | Properties |
|---|---|---|
| Class I | Alternative splicing |
|
| Applied load |
| |
| Motor domain phosphorylation |
| |
| Phosphorylation within the tail |
| |
| Mg2+ |
| |
| Ca2+ |
| |
| Phospholipids |
| |
| Class III | Alternative splicing within the neck and tail region (myosin IIIb) |
|
| Motor domain phosphorylation |
| |
| Class V | Alternative splicing in the region between the neck and the GTD |
|
| Phosphorylation within the tail |
| |
| Mg2+ |
| |
| Ca2+ |
| |
| Binding partners |
| |
| Class VI | Alternative splicing within the CBD |
|
| Applied load |
| |
| Motor domain phosphorylation |
| |
| Phosphorylation within CBD |
| |
| Mg2+ |
| |
| Ca2+ |
| |
| Phospholipids (PtdIns(4,5)P2 |
| |
| Binding partners |
| |
| Class VII | Mg2+ |
|
| Ca2+ |
| |
| Binding partners |
| |
| Class X | Phospholipids (PtdIns(3,4,5)P3) |
|
| Class XV | Alternative splicing N-terminal to the motor domain (myosin XVa) | None defined |
| Class XVIII | Alternative splicing within the tail |
|
| Myosin | Interplay between Modes of Regulation: Effect on Myosin Function |
|---|---|
| Class I | Force sensing:
|
| Class V | Association with cargo:
|
| Class VI | Intracellular localization and function:
|
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fili, N.; Toseland, C.P. Unconventional Myosins: How Regulation Meets Function. Int. J. Mol. Sci. 2020, 21, 67. https://doi.org/10.3390/ijms21010067
Fili N, Toseland CP. Unconventional Myosins: How Regulation Meets Function. International Journal of Molecular Sciences. 2020; 21(1):67. https://doi.org/10.3390/ijms21010067
Chicago/Turabian StyleFili, Natalia, and Christopher P. Toseland. 2020. "Unconventional Myosins: How Regulation Meets Function" International Journal of Molecular Sciences 21, no. 1: 67. https://doi.org/10.3390/ijms21010067
APA StyleFili, N., & Toseland, C. P. (2020). Unconventional Myosins: How Regulation Meets Function. International Journal of Molecular Sciences, 21(1), 67. https://doi.org/10.3390/ijms21010067