Identification, Localization in the Central Nervous System and Novel Myostimulatory Effect of Allatostatins in Tenebrio molitor Beetle


Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
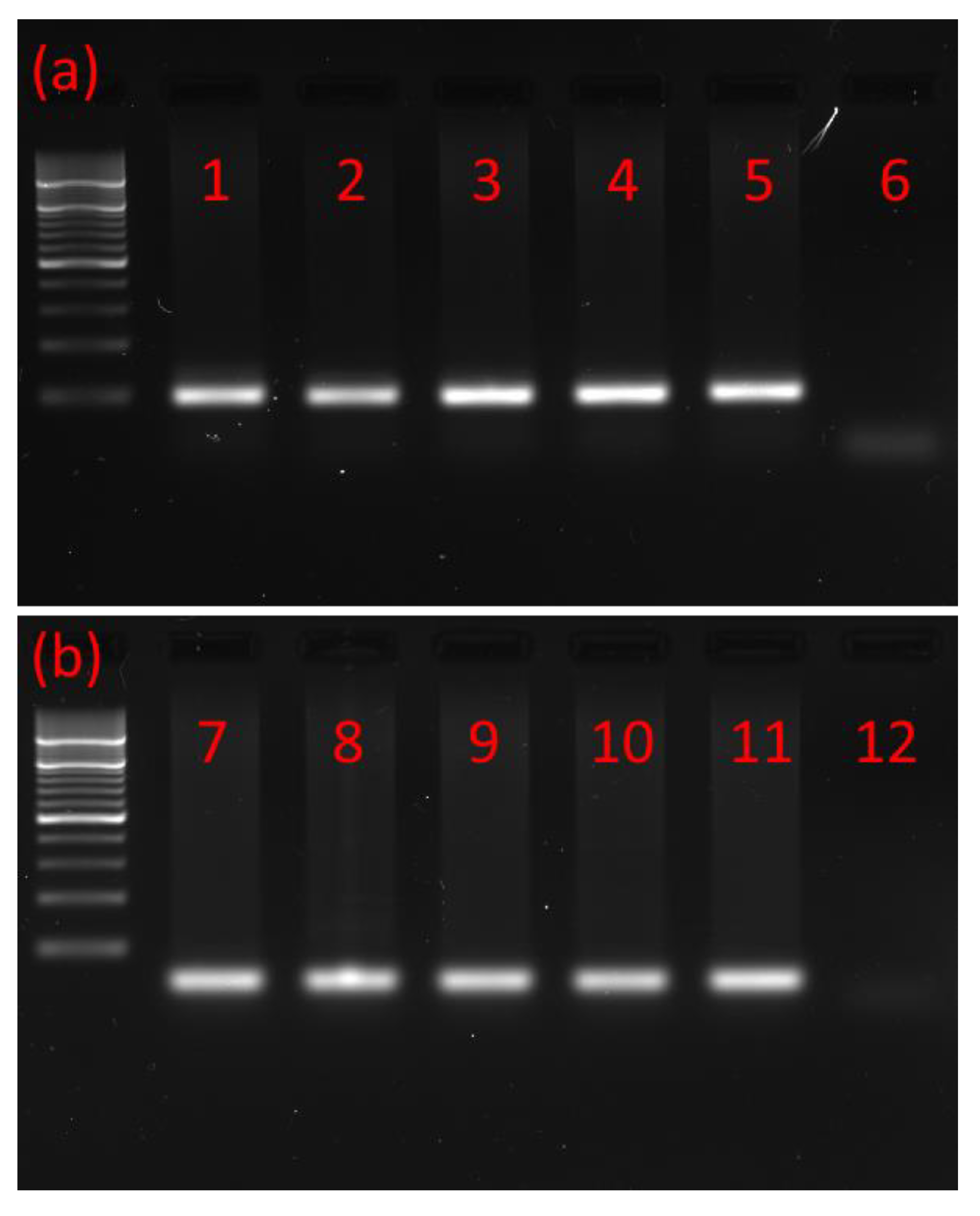
2.1. mRNA Identification
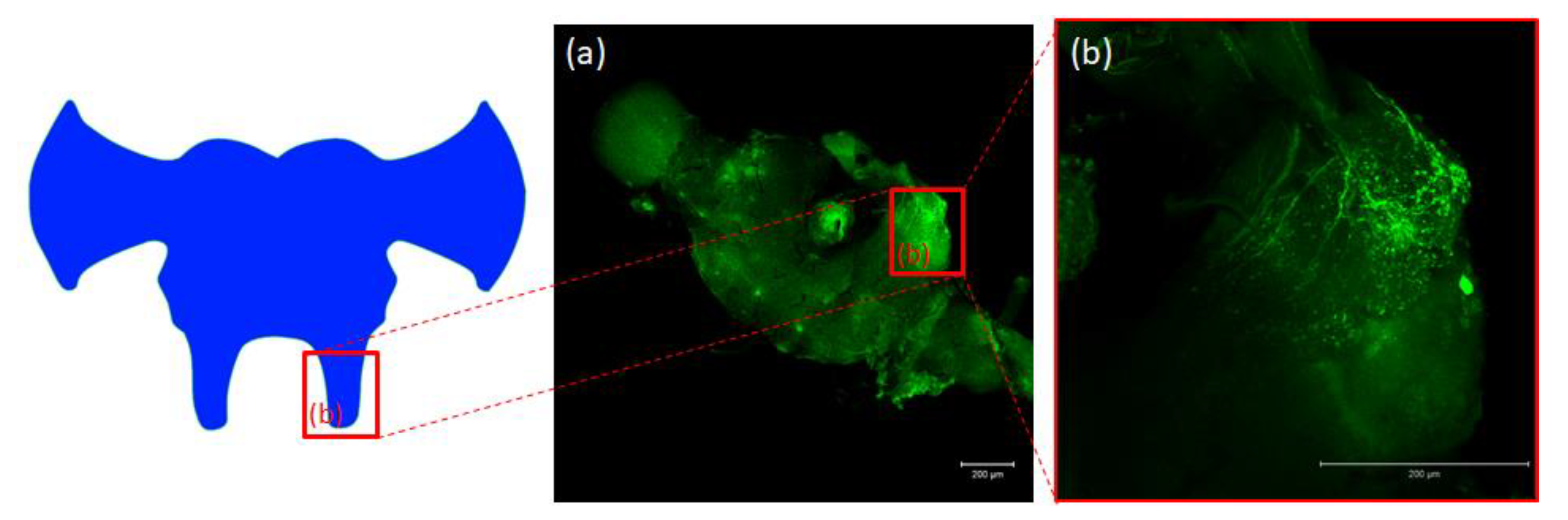
2.2. Immunolocalization
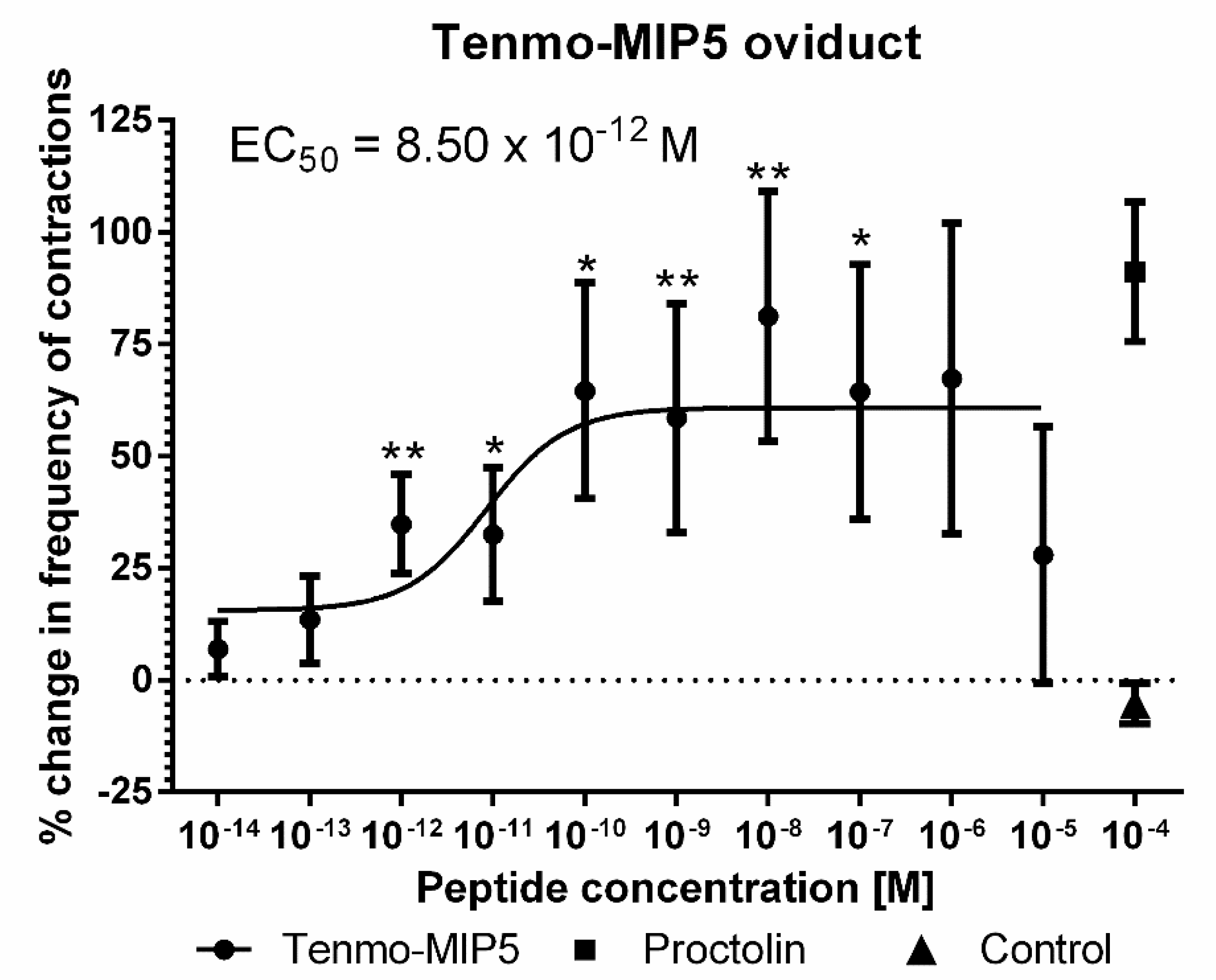
2.3. Oviduct Contractions
3. Discussion
4. Materials and Methods
4.1. Insects
4.2. Isolation of mRNA and Generation of cDNA
4.3. Primer Design
4.4. BLAST Search
4.5. Immunohistochemistry
4.6. In Vitro Oviduct Bioassay
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scherkenbeck, J.; Zdobinsky, T. Insect neuropeptides: Structures, chemical modifications and potential for insect control. Bioorg. Med. Chem. 2009, 17, 4071–4084. [Google Scholar] [CrossRef]
- Taghert, P.H.; Nitabach, M.N. Peptide neuromodulation in invertebrate model systems. Neuron 2012, 76, 82–97. [Google Scholar] [CrossRef]
- Nassel, D.R.; Zandawala, M. Recent advances in neuropeptide signaling in Drosophila, from genes to physiology and behavior. Prog. Neurobiol. 2019, 101607. [Google Scholar] [CrossRef]
- Chowanski, S.; Adamski, Z.; Lubawy, J.; Marciniak, P.; Pacholska-Bogalska, J.; Slocinska, M.; Spochacz, M.; Szymczak, M.; Urbanski, A.; Walkowiak-Nowicka, K.; et al. Insect Peptides—Perspectives in Human Diseases Treatment. Curr. Med. Chem. 2017, 24, 3116–3152. [Google Scholar] [CrossRef]
- Chowanski, S.; Lubawy, J.; Urbanski, A.; Rosinski, G. Cardioregulatory Functions of Neuropeptides and Peptide Hormones in Insects. Protein Pept. Lett. 2016, 23, 913–931. [Google Scholar] [CrossRef]
- Adamski, Z.; Bufo, S.A.; Chowanski, S.; Falabella, P.; Lubawy, J.; Marciniak, P.; Pacholska-Bogalska, J.; Salvia, R.; Scrano, L.; Slocinska, M.; et al. Beetles as Model Organisms in Physiological, Biomedical and Environmental Studies—A Review. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Hartenstein, V. The neuroendocrine system of invertebrates: A developmental and evolutionary perspective. J. Endocrinol. 2006, 190, 555–570. [Google Scholar] [CrossRef]
- Urbanski, A.; Lubawy, J.; Marciniak, P.; Rosinski, G. Myotropic activity and immunolocalization of selected neuropeptides of the burying beetle Nicrophorus vespilloides (Coleoptera: Silphidae). Insect Sci. 2019, 26, 656–670. [Google Scholar] [CrossRef]
- Verlinden, H.; Gijbels, M.; Lismont, E.; Lenaerts, C.; Broeck, J.V.; Marchal, E. The pleiotropic allatoregulatory neuropeptides and their receptors: A mini-review. J. Insect Physiol. 2015, 80, 2–14. [Google Scholar] [CrossRef]
- Coast, G.M.; Schooley, D.A. Toward a consensus nomenclature for insect neuropeptides and peptide hormones. Peptides 2011, 32, 620–631. [Google Scholar] [CrossRef]
- Marciniak, P.; Szymczak, M.; Rosinski, G. Hormony peptydowe owadów—Przegląd najważniejszych rodzin. Post. Biol. Komórki 2011, 38, 43–63. [Google Scholar]
- Bendena, W.G.; Tobe, S.S. Families of allatoregulator sequences: A 2011 perspective 1 1 This review is part of a virtual symposium on recent advances in understanding a variety of complex regulatory processes in insect physiology and endocrinology, including development, metabolism, cold hardiness, food intake and digestion, and diuresis, through the use of omics technologies in the postgenomic era. Can. J. Zool. 2012, 90, 521–544. [Google Scholar]
- Audsley, N.; Vandersmissen, H.P.; Weaver, R.; Dani, P.; Matthews, J.; Down, R.; Vuerinckx, K.; Kim, Y.J.; Vanden Broeck, J. Characterisation and tissue distribution of the PISCF allatostatin receptor in the red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 2013, 43, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Bachtel, N.D.; Hovsepian, G.A.; Nixon, D.F.; Eleftherianos, I. Allatostatin C modulates nociception and immunity in Drosophila. Sci. Rep. 2018, 8, 7501. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Piulachs, M.D.; Belles, X. Inhibition of vitellogenin production by allatostatin in the German cockroach. Mol. Cell. Endocrinol. 1996, 121, 191–196. [Google Scholar] [CrossRef]
- Martin, D.; Piulachs, M.D.; Belles, X. Allatostatin inhibits vitellogenin release in a cockroach. Ann. N. Y. Acad. Sci. 1998, 839, 341–342. [Google Scholar] [CrossRef]
- Fuse, M.; Zhang, J.R.; Partridge, E.; Nachman, R.J.; Orchard, I.; Bendena, W.G.; Tobe, S.S. Effects of an allatostatin and a myosuppressin on midgut carbohydrate enzyme activity in the cockroach Diploptera punctata. Peptides 1999, 20, 1285–1293. [Google Scholar] [CrossRef]
- Lange, A.B.; Alim, U.; Vandersmissen, H.P.; Mizoguchi, A.; Vanden Broeck, J.; Orchard, I. The distribution and physiological effects of the myoinhibiting peptides in the kissing bug, Rhodnius prolixus. Front. Neurosci. 2012, 6, 98. [Google Scholar] [CrossRef]
- Lorenz, M.W.; Kellner, R.; Hoffmann, K.H. A family of neuropeptides that inhibit juvenile-hormone biosynthesis in the cricket, Gryllus-bimaculatus. J. Biol. Chem. 1995, 270, 21103–21108. [Google Scholar] [CrossRef]
- Lubawy, J.; Czarniewska, E.; Kuczer, M.; Rosinski, G. Allatostatins—Pleiotropic insect neurohormones. Post. Biol. Komórki 2013, 40, 385–400. [Google Scholar]
- Audsley, N.; Matthews, J.; Weaver, R.J. Neuropeptides associated with the frontal ganglion of larval Lepidoptera. Peptides 2005, 26, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, M.B.; Jaffe, H.; Kochansky, J.; Raina, A.K. Identification of four additional myoinhibitory peptides (MIPs) from the ventral nerve cord of Manduca sexta. Arch. Insect Biochem. Physiol. 2001, 48, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, M.B.; Wagner, R.M.; Kochansky, J.P.; Harrison, D.J.; Thomas-Laemont, P.; Raina, A.K. The identification of two myoinhibitory peptides, with sequence similarities to the galanins, isolated from the ventral nerve cord of Manduca sexta. Regul. Pept. 1995, 57, 213–219. [Google Scholar] [CrossRef]
- Sedra, L.; Haddad, A.S.; Lange, A.B. Myoinhibitors controlling oviduct contraction within the female blood-gorging insect, Rhodnius prolixus. Gen. Comp. Endocrinol. 2015, 211, 62–68. [Google Scholar] [CrossRef]
- Matthews, H.J.; Audsley, N.; Weaver, R.J. Interactions between allatostatins and allatotropin on spontaneous contractions of the foregut of larval Lacanobia oleracea. J. Insect Physiol. 2007, 53, 75–83. [Google Scholar] [CrossRef]
- Price, M.D.; Merte, J.; Nichols, R.; Koladich, P.M.; Tobe, S.S.; Bendena, W.G. Drosophila melanogaster flatline encodes a myotropin orthologue to Manduca sexta allatostatin. Peptides 2002, 23, 787–794. [Google Scholar] [CrossRef]
- Diaz, M.M.; Schlichting, M.; Abruzzi, K.C.; Long, X.; Rosbash, M. Allatostatin-C/AstC-R2 is a Novel Pathway to Modulate the Circadian Activity Pattern in Drosophila. Curr. Biol. 2019, 29, 13–22. [Google Scholar] [CrossRef]
- Veenstra, J.A. Allatostatin C and its paralog allatostatin double C: The arthropod somatostatins. Insect Biochem. Mol. Biol. 2009, 39, 161–170. [Google Scholar] [CrossRef]
- Veenstra, J.A. Allatostatins C, double C and triple C, the result of a local gene triplication in an ancestral arthropod. Gen. Comp. Endocrinol. 2016, 230–231, 153–157. [Google Scholar] [CrossRef]
- Kreienkamp, H.J.; Larusson, H.J.; Witte, I.; Roeder, T.; Birgul, N.; Honck, H.H.; Harder, S.; Ellinghausen, G.; Buck, F.; Richter, D. Functional annotation of two orphan G-protein-coupled receptors, Drostar1 and -2, from Drosophila melanogaster and their ligands by reverse pharmacology. J. Biol. Chem. 2002, 277, 39937–39943. [Google Scholar] [CrossRef]
- Birgul, N.; Weise, C.; Kreienkamp, H.J.; Richter, D. Reverse physiology in drosophila: Identification of a novel allatostatin-like neuropeptide and its cognate receptor structurally related to the mammalian somatostatin/galanin/opioid receptor family. EMBO J. 1999, 18, 5892–5900. [Google Scholar] [CrossRef]
- Urlacher, E.; Soustelle, L.; Parmentier, M.-L.; Verlinden, H.; Gherardi, M.-J.; Fourmy, D.; Mercer, A.R.; Devaud, J.-M.; Massou, I. Honey Bee Allatostatins Target Galanin/Somatostatin-Like Receptors and Modulate Learning: A Conserved Function? PLoS ONE 2016, 11, e0146248. [Google Scholar] [CrossRef] [PubMed]
- De Loof, A.; Lindemans, M.; Liu, F.; De Groef, B.; Schoofs, L. Endocrine archeology: Do insects retain ancestrally inherited counterparts of the vertebrate releasing hormones GnRH, GHRH, TRH, and CRF? Gen. Comp. Endocrinol. 2012, 177, 18–27. [Google Scholar] [CrossRef] [PubMed]
- De Loof, A.; Schoofs, L.; Huybrechts, R. The endocrine system controlling sexual reproduction in animals: Part of the evolutionary ancient but well conserved immune system? Gen. Comp. Endocrinol. 2016, 226, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Lubawy, J.; Marciniak, P.; Kuczer, M.; Rosiński, G. Myotropic activity of allatostatins in tenebrionid beetles. Neuropeptides 2018. [Google Scholar] [CrossRef]
- Williams, E.A.; Conzelmann, M.; Jekely, G. Myoinhibitory peptide regulates feeding in the marine annelid Platynereis. Front. Zool. 2015, 12, 1. [Google Scholar] [CrossRef]
- Alzugaray, M.E.; Hernandez-Martinez, S.; Ronderos, J.R. Somatostatin signaling system as an ancestral mechanism: Myoregulatory activity of an Allatostatin-C peptide in Hydra. Peptides 2016, 82, 67–75. [Google Scholar] [CrossRef]
- Ito, M.; Masuda, N.; Shinomiya, K.; Endo, K.; Ito, K. Systematic analysis of neural projections reveals clonal composition of the Drosophila brain. Curr. Biol. 2013, 23, 644–655. [Google Scholar] [CrossRef]
- Weaver, R.J.; Audsley, N. Neuropeptide regulators of juvenile hormone synthesis: Structures, functions, distribution, and unanswered questions. Ann. N. Y. Acad. Sci. 2009, 1163, 316–329. [Google Scholar] [CrossRef]
- Matsumoto, K.; Suetsugu, Y.; Tanaka, Y.; Kotaki, T.; Goto, S.G.; Shinoda, T.; Shiga, S. Identification of allatostatins in the brown-winged green bug Plautia stali. J. Insect Physiol. 2017, 96, 21–28. [Google Scholar] [CrossRef]
- Audsley, N.; Matthews, H.J.; Price, N.R.; Weaver, R.J. Allatoregulatory peptides in Lepidoptera, structures, distribution and functions. J. Insect Physiol. 2008, 54, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Schoofs, L.; Holman, G.M.; Hayes, T.K.; Nachman, R.J.; De Loof, A. Isolation, identification and synthesis of locustamyoinhibiting peptide (LOM-MIP), a novel biologically active neuropeptide from Locusta migratoria. Regul. Pept. 1991, 36, 111–119. [Google Scholar] [CrossRef]
- Taghert, P.H.; Veenstra, J.A. Drosophila neuropeptide signaling. Adv. Genet. 2003, 49, 1–65. [Google Scholar] [PubMed]
- Tsukamoto, Y.; Nagata, S. Newly identified allatostatin Bs and their receptor in the two-spotted cricket, Gryllus bimaculatus. Peptides 2016, 80, 25–31. [Google Scholar] [CrossRef]
- Veenstra, J.A. Coleoptera genome and transcriptome sequences reveal numerous differences in neuropeptide signaling between species. Peerj 2019, 7. [Google Scholar] [CrossRef]
- Pandit, A.A.; Davies, S.A.; Smagghe, G.; Dow, J.A.T. Evolutionary trends of neuropeptide signaling in beetles—A comparative analysis of Coleopteran transcriptomic and genomic data. Insect Biochem. Mol. Biol. 2019, 114, 103227. [Google Scholar] [CrossRef]
- Li, B.; Predel, R.; Neupert, S.; Hauser, F.; Tanaka, Y.; Cazzamali, G.; Williamson, M.; Arakane, Y.; Verleyen, P.; Schoofs, L.; et al. Genomics, transcriptomics, and peptidomics of neuropeptides and protein hormones in the red flour beetle Tribolium castaneum. Genome Res. 2008, 18, 113–122. [Google Scholar] [CrossRef]
- Predel, R.; Rapus, J.; Eckert, M. Myoinhibitory neuropeptides in the American cockroach. Peptides 2001, 22, 199–208. [Google Scholar] [CrossRef]
- Schulze, J.; Neupert, S.; Schmidt, L.; Predel, R.; Lamkemeyer, T.; Homberg, U.; Stengl, M. Myoinhibitory peptides in the brain of the cockroach Leucophaea maderae and colocalization with pigment-dispersing factor in circadian pacemaker cells. J. Comp. Neurol. 2012, 520, 1078–1097. [Google Scholar] [CrossRef]
- Weaver, R.J.; Audsley, N. Neuropeptides of the beetle, Tenebrio molitor identified using MALDI-TOF mass spectrometry and deduced sequences from the Tribolium castaneum genome. Peptides 2008, 29, 168–178. [Google Scholar] [CrossRef]
- Marciniak, P.; Audsley, N.; Kuczer, M.; Rosinski, G. Identification of myotropic neuropeptides from the brain and corpus cardiacum-corpus allatum complex of the beetle, Zophobas atratus. J. Insect Sci. 2010, 10, 156. [Google Scholar] [CrossRef] [PubMed]
- Heuer, C.M.; Kollmann, M.; Binzer, M.; Schachtner, J. Neuropeptides in insect mushroom bodies. Arthropod Struct. Dev. 2012, 41, 199–226. [Google Scholar] [CrossRef] [PubMed]
- Chapman, R.F. The insects: Structure and function. In Biological Science Texts; Cambridge University Press: New York, NY, USA, 2006; Volume xii, p. 819. [Google Scholar]
- Liessem, S.; Ragionieri, L.; Neupert, S.; Buschges, A.; Predel, R. Transcriptomic and Neuropeptidomic Analysis of the Stick Insect, Carausius morosus. J. Proteome Res. 2018, 17, 2192–2204. [Google Scholar] [CrossRef] [PubMed]
- Abdel-latief, M.; Hoffmann, K.H. Neuropeptide regulators of the juvenile hormone biosynthesis (in vitro) in the beetle, Tenebrio molitor (Coleoptera, Tenebrionidae). Arch. Insect Biochem. Physiol. 2010, 74, 135–146. [Google Scholar] [CrossRef]
- Zars, T. Behavioral functions of the insect mushroom bodies. Curr. Opin. Neurobiol. 2000, 10, 790–795. [Google Scholar] [CrossRef]
- Hergarden, A.C.; Tayler, T.D.; Anderson, D.J. Allatostatin-A neurons inhibit feeding behavior in adult Drosophila. Proc. Natl. Acad. Sci. USA 2012, 109, 3967–3972. [Google Scholar] [CrossRef]
- Schoofs, L.; Veelaert, D.; Broeck, J.V.; De Loof, A. Immunocytochemical distribution of locustamyoinhibiting peptide (Lom-MIP) in the nervous system of Locusta migratoria. Regul. Pept. 1996, 63, 171–179. [Google Scholar]
- Marciniak, P.; Kuczer, M.; Rosinski, G. New physiological activities of myosuppressin, sulfakinin and NVP-like peptide in Zophobas atratus beetle. J. Comp. Physiol. B 2011, 181, 721–730. [Google Scholar] [CrossRef][Green Version]
- Konopinska, D.; Rosinski, G. Proctolin, an insect neuropeptide. J. Pept. Sci. 1999, 5, 533–546. [Google Scholar] [CrossRef]
- Down, R.E.; Matthews, H.J.; Audsley, N. Effects of Manduca sexta allatostatin and an analog on the pea aphid Acyrthosiphon pisum (Hemiptera: Aphididae) and degradation by enzymes from the aphid gut. Peptides 2010, 31, 489–497. [Google Scholar] [CrossRef]
- Marciniak, P.; Szymczak, M.; Pacholska-Bogalska, J.; Audsley, N.; Rosinski, G. Identification and localisation of selected myotropic neuropeptides in the ventral nerve cord of tenebrionid beetles. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 166, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Morales-Ramos, J.; Rojas, M.; Shapiro-Ilan, D.; Tedders, W. Developmental plasticity in Tenebrio molitor (Coleoptera: Tenebrionidae): Analysis of instar variation in number and development time under different diets. J. Entomol. Sci. 2010, 45, 75–90. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lubawy, J.; Marciniak, P.; Rosiński, G. Identification, Localization in the Central Nervous System and Novel Myostimulatory Effect of Allatostatins in Tenebrio molitor Beetle. Int. J. Mol. Sci. 2020, 21, 3510. https://doi.org/10.3390/ijms21103510
Lubawy J, Marciniak P, Rosiński G. Identification, Localization in the Central Nervous System and Novel Myostimulatory Effect of Allatostatins in Tenebrio molitor Beetle. International Journal of Molecular Sciences. 2020; 21(10):3510. https://doi.org/10.3390/ijms21103510
Chicago/Turabian StyleLubawy, Jan, Paweł Marciniak, and Grzegorz Rosiński. 2020. "Identification, Localization in the Central Nervous System and Novel Myostimulatory Effect of Allatostatins in Tenebrio molitor Beetle" International Journal of Molecular Sciences 21, no. 10: 3510. https://doi.org/10.3390/ijms21103510
APA StyleLubawy, J., Marciniak, P., & Rosiński, G. (2020). Identification, Localization in the Central Nervous System and Novel Myostimulatory Effect of Allatostatins in Tenebrio molitor Beetle. International Journal of Molecular Sciences, 21(10), 3510. https://doi.org/10.3390/ijms21103510