Adipose-Derived Stem Cells Promote Intussusceptive Lymphangiogenesis by Restricting Dermal Fibrosis in Irradiated Tissue of Mice


Abstract
:1. Introduction
2. Results
2.1. Macroscopic Observation of Hind Limb Swelling
2.2. The Number and Area of Lymphatic Vessels with LYVE-1 Immunoreactivity
2.3. Analysis of LEC Proliferative Activity
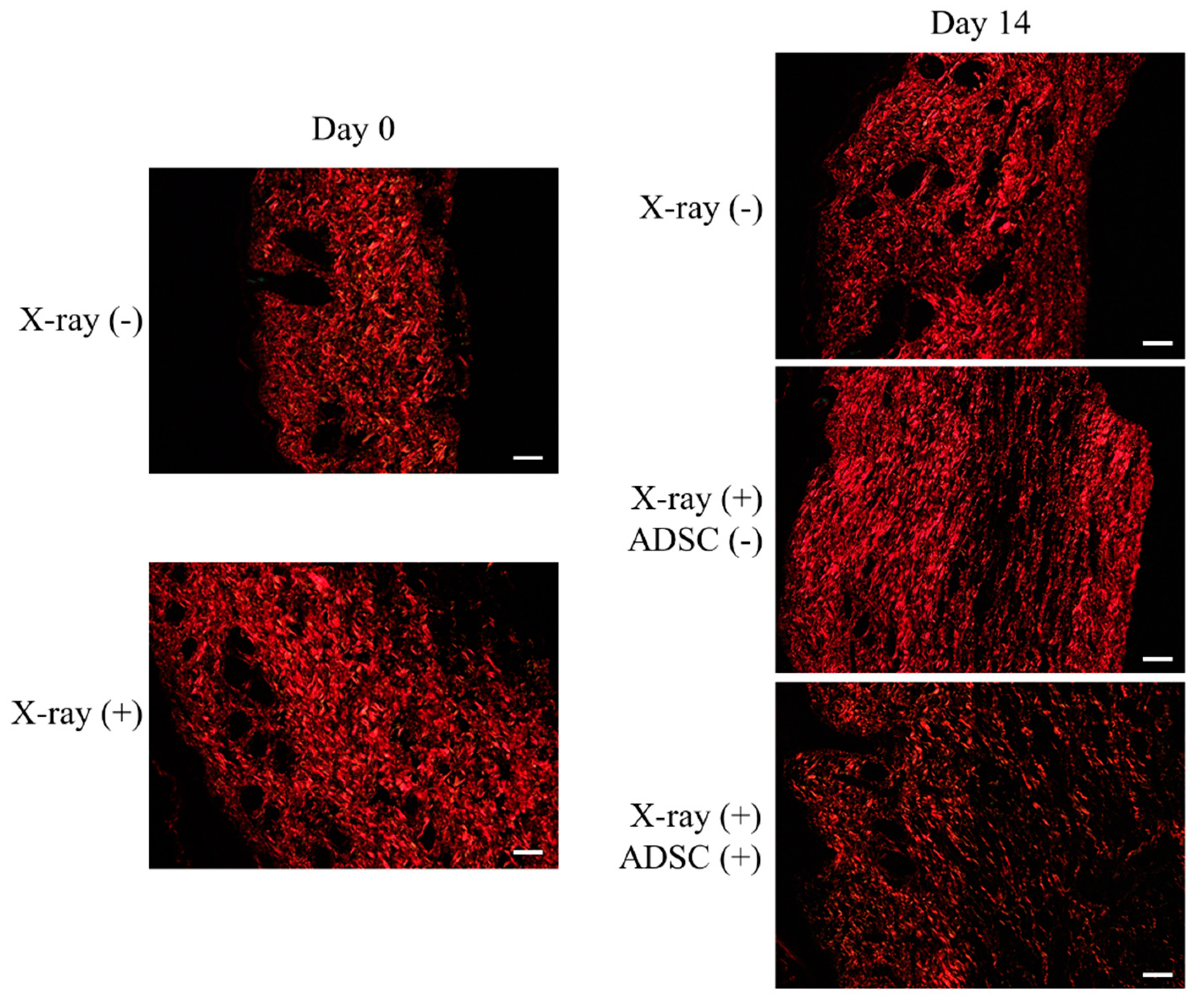
2.4. Evaluation of Fibrosis Using Picro-Sirius Red Staining
- X-ray/ADSC (−/−) group, day 0: Collagen fibers of type I randomly oriented and type III spread throughout the specimen.
- X-ray/ADSC (−/−) group, day 14: Shorter and still randomly oriented type I collagen fibers; type III collagen fibers only in gaps of type I fibers.
- X-ray (+) groups (X-ray/ADSC (+/−) and X-ray/ADSC (+/+)), day 0: Collagen fibers of type I densely deposited and type III rarely found.
- X-ray/ADSC (+/−) group, day 14: Thicker and longer type I collagen fibers deposited in parallel. Type III collagen fibers only in places of a low density of type I collagen fibers.
- X-ray/ADSC (+/+) group, day 14: Randomly oriented type I collagen fibers with lower density compared to the other two groups. Type III collagen fibers spread throughout the specimen, mainly in gaps of type I collagen fibers.
2.5. Gene Expression Analysis
- X-irradiation did not change Prox1 expression (day 0, mean ± standard error; X-ray (−), X-ray (+): 1.00 ± 0.25, 0.64 ± 0.11, respectively). In the X-ray/ADSC (+/+) group, Prox1 expression tended to increase from day 2 and significantly increased at day 14 (day 14: 1.93 ± 0.26, p < 0.01 compared with day 0). In the X-ray/ADSC (−/−) group, it increased significantly at day 8 (day 8: 1.88 ± 0.23, p < 0.05 compared with day 0).
- X-irradiation significantly suppressed Vegfc expression (day 0; X-ray (−), X-ray (+): 1.00 ± 0.06, 0.47 ± 0.09, respectively). Lymphatic incision induced a steep decrease in Vegfc expression, and was sustained in the X-ray/ADSC (+/+) group (day14; X-ray/ADSC (−/−), X-ray/ADSC (+/−), X-ray/ADSC (+/+): 0.98 ± 0.13, 0.80 ± 0.12, 0.19 ± 0.14, respectively).
- X-irradiation did not change Fgf2 expression (day 0; X-ray (−), X-ray (+): 1.00 ± 0.23, 0.64 ± 0.09, respectively). In the X-ray/ADSC (+/+) group, Fgf2 expression increased significantly at day 14 compared with X-ray/ADSC (+/−) group (X-ray/ADSC (−/−), X-ray/ADSC (+/−), X-ray/ADSC (+/+): 0.90 ± 0.10, 0.35 ± 0.07, 0.86 ± 0.13, respectively).
- Expression of Hgf tended to increase, but not significantly, after x-irradiation (day 0; X-ray (−), X-ray (+): 1.00 ± 0.09, 2.27 ± 0.47, respectively). Hgf expression did not change significantly during both inter-day or inter-group analysis.
- Tgfb1 expression did not change by x-irradiation (day 0; X-ray (−), X-ray (+): 1.00 ± 0.19, 0.98 ± 0.10, respectively). Expression of Tgfb1 mRNA in the X-ray/ADSC (+/+) group significantly increased compared with the other groups from day 8, and increased approximately 3-fold at day 14 (day 14; X-ray/ADSC (−/−), X-ray/ADSC (+/−), X-ray/ADSC (+/+): 1.25 ± 0.17, 1.03 ± 0.18, 3.50 ± 0.37, respectively). In addition, in the X-ray/ADSC (−/−) group, it significantly increased at day 8 (X-ray/ADSC (−/−) day 8: 1.63 ± 0.23, p < 0.01 compared with day 0).
- Col1a1 expression did not change significantly by x-irradiation (day 0; X-ray (−), X-ray (+): 1.00 ± 0.12, 0.63 ± 0.21, respectively). At day 2, expression of Col1a1 mRNA markedly decreased in all groups (day 2; X-ray/ADSC (−/−), X-ray/ADSC (+/−), X-ray/ADSC (+/+): 0.01 ± 0.01, 0.02 ± 0.01, 0.02 ± 0.00, respectively). When compared with its expression at day 2, that in X-ray/ADSC (−/−) group significantly increased at day 8 (X-ray/ADSC (−/−) day 8: 0.62 ± 0.04, p < 0.01) and that in X-ray/ADSC (+/+) group significantly increased at day 14 (X-ray/ADSC (+/+) day 14: 0.61 ± 0.08, p < 0.01).
2.6. SEM Observation
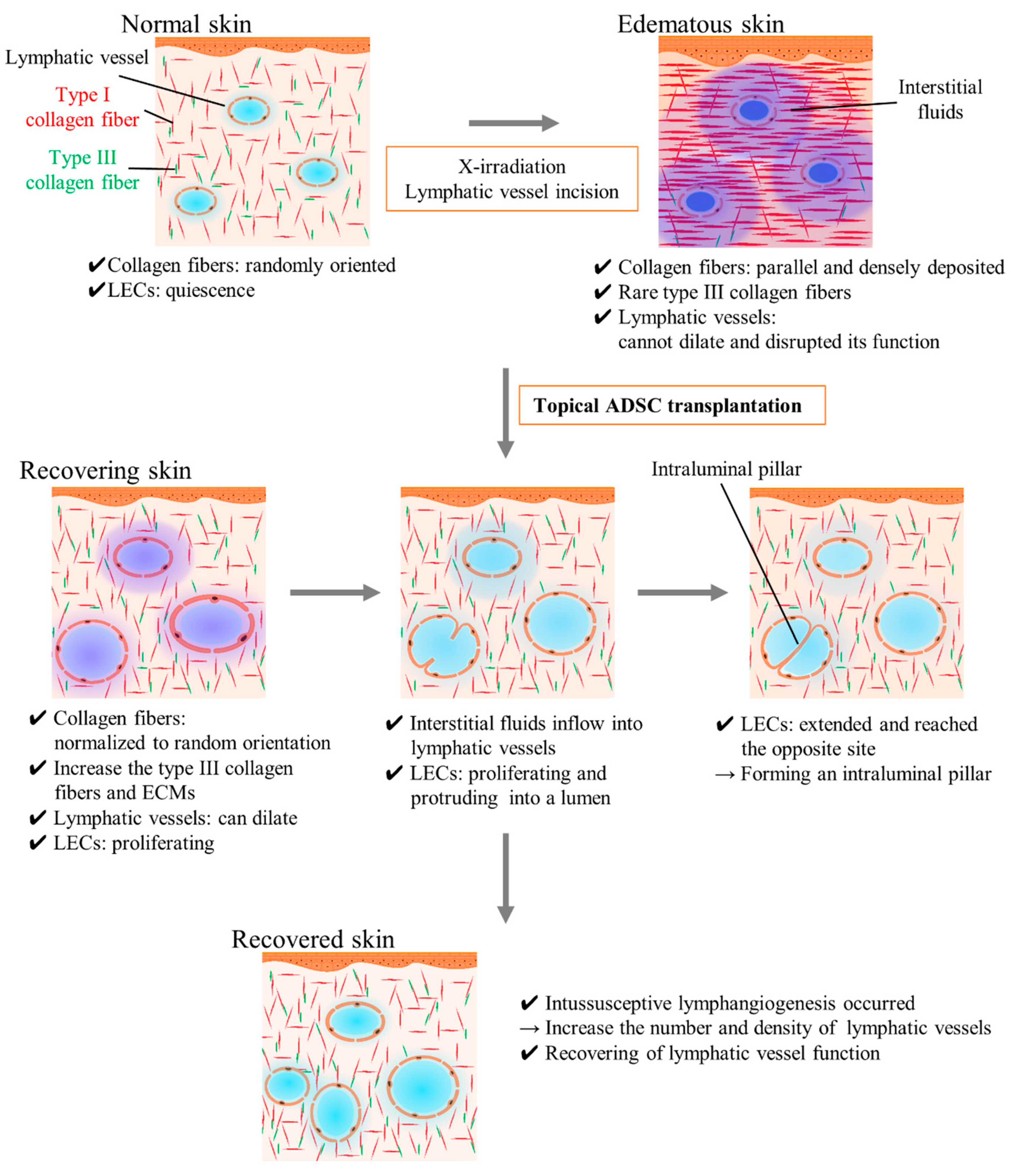
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Mouse Secondary Lymphedema Model and Surgical Preparation
4.3. Preparation of Adipose-Derived Stem Cells
4.4. Grouping of the Experimental Animals
- X-ray/ADSC (−/−) group: No-x-irradiated and mock injected group, n = 6.
- X-ray/ADSC (+/−) group: x-irradiated and mock injected group, n = 6.
- X-ray/ADSC (+/+) group: x-irradiated and ADSC transplanted group, n = 6.
4.5. Histological Examination
4.6. LYVE-1 Immunoreactivity
4.7. Measurement of LEC Proliferation Activity
4.8. Evaluation of Fibrosis Using Picro-Sirius Red Staining
4.9. Gene Expression Analysis
4.10. Observation Using SEM
4.11. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADSC | adipose-derived stem cell |
| BMSC | bone marrow derived stem cell |
| ECM | extracellular matrix |
| LEC | lymphatic endothelial cell |
| LYVE-1 | lymphatic vessel hyaluronan receptor 1 |
| MSC | mesenchymal stem cell |
| PCNA | proliferating cell nuclear antigen |
| SEM | scanning electron microscopy |
| TBS | tris(hydroxymethyl)aminomethane-buffered saline |
| TBS-T | tris(hydroxymethyl)aminomethane-buffered saline containing 0.1% Tween-20 |
| TGF-β | transforming growth factor-beta |
| VEGF | vascular endothelial growth factor |
| VEGFR | vascular endothelial growth factor receptor |
References
- Francois, M.; Caprini, A.; Hosking, B.; Orsenigo, F.; Wilhelm, D.; Browne, C.; Paavonen, K.; Karnezis, T.; Shayan, R.; Downes, M.; et al. Sox18 induces development of the Lymphatic vasculature in mice. Nature 2008, 456, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Nakagami, H.; Kaneda, Y.; Morishita, R. Lymphedema and therapeutic Lymphangiogenesis. BioMed Res. Int. 2013, 2013, 804675. [Google Scholar] [CrossRef] [Green Version]
- Ghezzi, F.; Uccella, S.; Cromi, A.; Bogani, G.; Robba, C.; Serati, M.; Bolis, P. Lymphoceles, Lymphorrhea, and Lymphedema after laparoscopic and open endometrial cancer staging. Ann. Surg. Oncol. 2011, 19, 259–267. [Google Scholar] [CrossRef]
- Avraham, T.; Clavin, N.W.; Daluvoy, S.V.; Fernandez, J.; Soares, M.A.; Cordeiro, A.P.; Mehrara, B.J. Fibrosis is a key inhibitor of lymphatic regeneration. Plast. Reconstr. Surg. 2009, 124, 438–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, C.; Assouad, J.; Riquet, M.; Hidden, G. Postmastectomy Lymphedema. Ann. Surg. 2006, 243, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Yamamoto, N.; Doi, K.; Oshima, A.; Yoshimatsu, H.; Todokoro, T.; Ogata, F.; Mihara, M.; Narushima, M.; Iida, T.; et al. Indocyanine green–enhanced Lymphography for upper extremity Lymphedema. Plast. Reconstr. Surg. 2011, 128, 941–947. [Google Scholar] [CrossRef]
- Koshima, I.; Narushima, M.; Yamamoto, Y.; Mihara, M.; Iida, T. Recent advancement on surgical treatments for Lymphedema. Ann. Vasc. Dis. 2012, 5, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Saaristo, A.M.; Niemi, T.; Viitanen, T.P.; Tervala, T.V.; Hartiala, P.; Suominen, E.A. Microvascular breast reconstruction and lymph node transfer for Postmastectomy Lymphedema patients. Ann. Surg. 2012, 255, 468–473. [Google Scholar] [CrossRef]
- Cheng, M.-H.; Chen, S.-C.; Henry, S.L.; Tan, B.K.; Lin, M.C.-Y.; Huang, J.-J. Vascularized groin lymph node flap transfer for Postmastectomy upper limb Lymphedema. Plast. Reconstr. Surg. 2013, 131, 1286–1298. [Google Scholar] [CrossRef]
- Chen, C.; Chiang, N.; Perng, C.; Ma, H.; Lin, C.-H. Review of preclinical and clinical studies of using cell-based therapy for secondary lymphedema. J. Surg. Oncol. 2019, 121. [Google Scholar] [CrossRef] [Green Version]
- Ongstad, E.L.; Bouta, E.M.; Roberts, J.E.; Uzarski, J.S.; Gibbs, S.E.; Sabel, M.S.; Cimmino, V.M.; Roberts, M.A.; Goldman, J. Lymphangiogenesis-independent resolution of experimental edema. Am. J. Physiol. Circ. Physiol. 2010, 299, H46–H54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oashi, K.; Furukawa, H.; Oyama, A.; Funayama, E.; Hayashi, T.; Saito, A.; Yamamoto, Y. A new model of acquired Lymphedema in the mouse hind limb. Ann. Plast. Surg. 2012, 69, 565–568. [Google Scholar] [CrossRef]
- Oashi, K.; Furukawa, H.; Nishihara, H.; Ozaki, M.; Oyama, A.; Funayama, E.; Hayashi, T.; Kuge, Y.; Yamamoto, Y. Pathophysiological characteristics of melanoma in-transit metastasis in a Lymphedema mouse model. J. Investig. Dermatol. 2013, 133, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.W.; Chong, S.Z.; Wong, F.H.S.; Evrard, M.; Tan, S.M.-L.; Keeble, J.; Kemeny, D.M.; Gascoigne, N.R.J.; Abastado, J.-P.; Angeli, V. Neutrophils contribute to inflammatory lymphangiogenesis by increasing VEGF-A bioavailability and secreting VEGF-D. Blood 2013, 122, 3666–3677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, D.; Yamamoto, Y.; Murao, N.; Oyama, A.; Funayama, E.; Furukawa, H. Establishment of an Acquired Lymphedema Model in the Mouse Hindlimb. Plast. Reconstr. Surg. 2017, 139, 67e–78e. [Google Scholar] [CrossRef] [PubMed]
- Tammela, T.; Saaristo, A.; Holopainen, T.; Lyytikkä, J.; Kotronen, A.; Pitkonen, M.; Abo-Ramadan, U.; Ylä-Herttuala, S.; Petrova, T.V.; Alitalo, K. Therapeutic differentiation and maturation of lymphatic vessels after lymph node dissection and transplantation. Nat. Med. 2007, 13, 1458–1466. [Google Scholar] [CrossRef] [PubMed]
- Conrad, C.; Niess, H.; Huss, R.; Huber, S.; von Luettichau, I.; Nelson, P.J.; Ott, H.C.; Jauch, K.-W.; Bruns, C.J. Multipotent mesenchymal stem cells acquire a lymphendothelial phenotype and enhance lymphatic regeneration in vivo. Circulation 2009, 119, 281–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oka, M.; Iwata, C.; Suzuki, H.I.; Kiyono, K.; Morishita, Y.; Watabe, T.; Komuro, A.; Kano, M.R.; Miyazono, K. Inhibition of endogenous TGF-β signaling enhances lymphangiogenesis. Blood 2008, 111, 4571–4579. [Google Scholar] [CrossRef] [Green Version]
- Serizawa, F.; Ito, K.; Matsubara, M.; Sato, A.; Shimokawa, H.; Satomi, S. Extracorporeal shock wave therapy induces therapeutic Lymphangiogenesis in a rat model of secondary lymphoedema. Eur. J. Vasc. Endovasc. Surg. 2011, 42, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Gousopoulos, E.; Proulx, S.T.; Bachmann, S.B.; Scholl, J.; Dionyssiou, D.; Demiri, E.; Halin, C.; Dieterich, L.C.; Detmar, M. Regulatory T cell transfer ameliorates lymphedema and promotes lymphatic vessel function. JCI Insight 2016, 1, e89081. [Google Scholar] [CrossRef] [Green Version]
- Mishima, T.; Ito, Y.; Nishizawa, N.; Amano, H.; Tsujikawa, K.; Miyaji, K.; Watanabe, M.; Majima, M. RAMP1 signaling improves lymphedema and promotes lymphangiogenesis in mice. J. Surg. Res. 2017, 219, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Jiang, Z.; Cui, C.; Sun, Y.; Lu, B.; Li, H.; Cao, W.; Chen, B.; Li, S.; Guo, L. The roles of podoplanin-positive/podoplanin-negative cells from adipose-derived stem cells in lymphatic regeneration. Plast. Reconstr. Surg. 2020, 145, 420–431. [Google Scholar] [CrossRef]
- Saijo, H.; Suzuki, K.; Yoshimoto, H.; Imamura, Y.; Yamashita, S.; Tanaka, K. Paracrine effects of adipose-derived stem cells promote lymphangiogenesis in irradiated lymphatic endothelial cells. Plast. Reconstr. Surg. 2019, 143, 1189e–1200e. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-H.; Shirado, T.; Mashiko, T.; Feng, J.; Asahi, R.; Kanayama, K.; Mori, M.; Chi, D.; Sunaga, A.; Sarukawa, S.; et al. Therapeutic effects of human adipose-derived products on impaired wound healing in irradiated tissue. Plast. Reconstr. Surg. 2018, 142, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Akita, S.; Yoshimoto, H.; Akino, K.; Ohtsuru, A.; Hayashida, K.; Hirano, A.; Suzuki, K.; Yamashita, S. Early experiences with stem cells in treating chronic wounds. Clin. Plast. Surg. 2012, 39, 281–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.-S.; Park, B.-S.; Park, S.-H.; Kim, H.-K.; Sung, J.-H. Antiwrinkle effect of adipose-derived stem cell: Activation of dermal fibroblast by secretory factors. J. Dermatol. Sci. 2009, 53, 96–102. [Google Scholar] [CrossRef]
- Murphy, M.B.; Moncivais, K.; Caplan, A. Mesenchymal stem cells: Environmentally responsive therapeutics for regenerative medicine. Exp. Mol. Med. 2013, 45, e54. [Google Scholar] [CrossRef] [Green Version]
- Quián, Y.P.; Ramirez, P.H.; Cuellar, J.F.B.; Pintado, A.P.; Pérez, M.A.C. Lymphoscintigraphy for the assessment of autologous stem cell implantation in chronic lymphedema. Clin. Nucl. Med. 2015, 40, 217–219. [Google Scholar] [CrossRef]
- Toyserkani, N.M.; Jensen, C.H.; Sheikh, S.P.; Sørensen, J.A. Cell-assisted lipotransfer using autologous adipose-derived stromal cells for alleviation of breast cancer-related lymphedema. STEM CELLS Transl. Med. 2016, 5, 857–859. [Google Scholar] [CrossRef] [Green Version]
- Toyserkani, N.M.; Jensen, C.H.; Andersen, D.C.; Sheikh, S.P.; Sørensen, J.A. Treatment of breast cancer-related lymphedema with adipose-derived regenerative cells and fat grafts: A feasibility and safety study. STEM CELLS Transl. Med. 2017, 6, 1666–1672. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Hamuy, R.; Hamada, Y.; Yoshimoto, H.; Hirano, A.; Akita, S. Adipose-derived stem cell transplantation for therapeutic lymphangiogenesis in a mouse secondary lymphedema model. Regen. Med. 2015, 10, 549–562. [Google Scholar] [CrossRef]
- Hayashida, K.; Yoshida, S.; Yoshimoto, H.; Fujioka, M.; Saijo, H.; Migita, K.; Kumaya, M.; Akita, S. Adipose-derived stem cells and vascularized lymph node transfers successfully treat mouse hindlimb secondary lymphedema by early reconnection of the lymphatic system and lymphangiogenesis. Plast. Reconstr. Surg. 2017, 139, 639–651. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Shibata, R.; Shintani, S.; Ishii, M.; Murohara, T. Therapeutic lymphangiogenesis with implantation of adipose-derived regenerative cells. J. Am. Hear. Assoc. 2012, 1, e000877. [Google Scholar] [CrossRef] [Green Version]
- Mendez, U.; Brown, E.M.; Ongstad, E.L.; Slis, J.R.; Goldman, J. Functional recovery of fluid drainage precedes lymphangiogenesis in acute murine foreleg lymphedema. Am. J. Physiol. Circ. Physiol. 2012, 302, H2250–H2256. [Google Scholar] [CrossRef]
- Avraham, T.; Yan, A.; Zampell, J.C.; Daluvoy, S.V.; Haimovitz-Friedman, A.; Cordeiro, A.P.; Mehrara, B. Radiation therapy causes loss of dermal lymphatic vessels and interferes with lymphatic function by TGF-beta1-mediated tissue fibrosis. Am. J. Physiol. Physiol. 2010, 299, C589–C605. [Google Scholar] [CrossRef] [Green Version]
- Zerbinati, N.; Calligaro, A. Calcium hydroxylapatite treatment of human skin: Evidence of collagen turnover through picrosirius red staining and circularly polarized microscopy. Clin. Cosmet. Investig. Dermatol. 2018, 11, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Clavin, N.W.; Avraham, T.; Fernandez, J.; Daluvoy, S.V.; Soares, M.A.; Chaudhry, A.; Mehrara, B.J. TGF-beta1 is a negative regulator of lymphatic regeneration during wound repair. Am J Physiol Heart Circ Physiol. 2008, 295, H2113–H2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamura, A.; Matsushita, T.; Komuro, A.; Kobayashi, T.; Maeda, S.; Hamaguchi, Y.; Takehara, K. Adipose-derived stromal/stem cells successfully attenuate the fibrosis of scleroderma mouse models. Int. J. Rheum. Dis. 2019, 23, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Tammela, T.; Alitalo, K. Lymphangiogenesis: Molecular mechanisms and future promise. Cell 2010, 140, 460–476. [Google Scholar] [CrossRef] [Green Version]
- Wigle, J.; Oliver, G. Prox1 function is required for the development of the murine lymphatic system. Cell 1999, 98, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, Z.; Wang, R.; Zhao, F.; Shi, P.; Jiang, Y.; Pang, X. Direct comparison of the potency of human mesenchymal stem cells derived from amnion tissue, bone marrow and adipose tissue at inducing dermal fibroblast responses to cutaneous wounds. Int. J. Mol. Med. 2012, 31, 407–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bliley, J.M.; Argenta, A.; Satish, L.; McLaughlin, M.M.; Dees, A.; Tompkins-Rhoades, C.; Marra, K.G.; Rubin, J.P. Administration of adipose-derived stem cells enhances vascularity, induces collagen deposition, and dermal adipogenesis in burn wounds. Burns 2016, 42, 1212–1222. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Rajantie, I.; Ilmonen, M.; Makinen, T.; Karkkainen, M.J.; Haiko, P.; Salven, P.; Alitalo, K. Preexisting lymphatic endothelium but not endothelial progenitor cells are essential for tumor lymphangiogenesis and lymphatic metastasis. Cancer Res. 2004, 64, 3737–3740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y. Vascular endothelial cell growth factor receptor 3-mediated activation of lymphatic endothelium is crucial for tumor cell entry and spread via lymphatic vessels. Cancer Res. 2005, 65, 4739–4746. [Google Scholar] [CrossRef] [Green Version]
- Bracher, A.; Cardona, A.S.; Tauber, S.; Fink, A.M.; Steiner, A.; Pehamberger, H.; Niederleithner, H.; Petzelbauer, P.; Gröger, M.; Loewe, R. Epidermal growth factor facilitates melanoma lymph node metastasis by influencing tumor lymphangiogenesis. J. Investig. Dermatol. 2013, 133, 230–238. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Lalani, A.S.; Harding, T.C.; Gonzalez, M.; Wu, W.-W.; Luan, B.; Tu, G.H.; Koprivnikar, K.; VanRoey, M.J.; He, Y.; et al. Inhibition of lymphogenous metastasis using adeno-associated virus-mediated gene transfer of a soluble VEGFR-3 decoy receptor. Cancer Res. 2005, 65, 6901–6909. [Google Scholar] [CrossRef] [Green Version]
- Jha, S.; Rauniyar, K.; Jeltsch, M. Key molecules in lymphatic development, function, and identification. Ann. Anat. Anat. Anz. 2018, 219, 25–34. [Google Scholar] [CrossRef]
- Vaahtomeri, K.; Karaman, S.; Makinen, T.; Alitalo, K. Lymphangiogenesis guidance by paracrine and pericellular factors. Genes Dev. 2017, 31, 1615–1634. [Google Scholar] [CrossRef]
- De Spiegelaere, W.; Casteleyn, C.; Broeck, W.V.D.; Plendl, J.; Bahramsoltani, M.; Simoens, P.; Djonov, V.; Cornillie, P. Intussusceptive angiogenesis: A biologically relevant form of angiogenesis. J. Vasc. Res. 2012, 49, 390–404. [Google Scholar] [CrossRef]
- Djonov, V.; Baum, O.; Burri, P.H. Vascular remodeling by intussusceptive angiogenesis. Cell and Tissue Res. 2003, 314, 107–117. [Google Scholar] [CrossRef]
- Baum, O.; Suter, F.; Gerber, B.; Tschanz, S.A.; Buergy, R.; Blank, F.; Hlushchuk, R.; Djonov, V. VEGF-A promotes intussusceptive angiogenesis in the developing chicken chorioallantoic membrane. Microcirculation 2010, 17. [Google Scholar] [CrossRef] [PubMed]
- Dimova, I.; Hlushchuk, R.; Makanya, A.; Styp-Rekowska, B.; Ceausu, A.R.; Flueckiger, S.; Lang, S.; Semela, D.; Le Noble, F.; Chatterjee, S.; et al. Inhibition of Notch signaling induces extensive intussusceptive neo-angiogenesis by recruitment of mononuclear cells. Angiogenesis 2013, 16, 921–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konerding, M.A.; Turhan, A.; Ravnic, D.J.; Lin, M.; Fuchs, C.; Secomb, T.; Tsuda, A.; Mentzer, S.J. Inflammation-induced intussusceptive angiogenesis in murine colitis. Anat. Rec. Adv. Integr. Anat. Evol. Boil. 2010, 293, 849–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvucci, O.; Yao, L.; Villalba, S.; Sajewicz, A.; Pittaluga, S.; Tosato, G. Regulation of endothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1. Blood 2002, 99, 2703–2711. [Google Scholar] [CrossRef] [Green Version]
- Dimova, I.; Karthik, S.; Makanya, A.; Hlushchuk, R.; Semela, D.; Volarevic, V.; Djonov, V. SDF-1/CXCR4 signalling is involved in blood vessel growth and remodelling by intussusception. J Cell Mol Med. 2019, 23, 3916–3926. [Google Scholar] [CrossRef]
- Vimalraj, S.; Bhuvaneswari, S.; Lakshmikirupa, S.; Jyothsna, G.; Chatterjee, S. Nitric oxide signaling regulates tumor-induced intussusceptive-like angiogenesis. Microvasc. Res. 2018, 119, 47–59. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; González-Gómez, M.; Carrasco, J.L.; Díaz-Flores, L. Intussusceptive lymphangiogenesis in the sinuses of developing human foetal lymph nodes. Ann. Anat. Anat. Anz. 2019, 226, 73–83. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.D.P.; Carrasco, J.L.; Sáez, F.; González-Gómez, M.; Madrid, J.F.; García, M.D.P. Intussusceptive lymphangiogenesis in lymphatic malformations/lymphangiomas. Anat. Rec. Adv. Integr. Anat. Evol. Boil. 2019, 302, 2003–2013. [Google Scholar] [CrossRef]
- Hassan, W.U.; Greiser, U.; Wang, W.-X. Role of adipose-derived stem cells in wound healing. Wound Repair Regen. 2014, 22, 313–325. [Google Scholar] [CrossRef]
- Hassanshahi, A.; Hassanshahi, M.; Khabbazi, S.; Hosseini-Khah, Z.; Peymanfar, Y.; Ghalamkari, S.; Su, Y.-W.; Xian, C.J.; Qhalamkari, S. Adipose-derived stem cells for wound healing. J. Cell. Physiol. 2018, 234, 7903–7914. [Google Scholar] [CrossRef]
- Ahmadzadeh, N.; Robering, J.W.; Kengelbach-Weigand, A.; Al-Abboodi, M.; Beier, J.P.; Horch, R.E.; Boos, A.M. Human adipose-derived stem cells support lymphangiogenesis in vitro by secretion of lymphangiogenic factors. Exp. Cell Res. 2020, 388, 111816. [Google Scholar] [CrossRef] [PubMed]
- Flanders, K.C.; Major, C.D.; Arabshahi, A.; Aburime, E.E.; Okada, M.H.; Fujii, M.; Blalock, T.D.; Schultz, G.S.; Sowers, A.; Anzano, M.A.; et al. Interference with transforming growth factor-beta/ Smad3 signaling results in accelerated healing of wounds in previously irradiated skin. Am. J. Pathol. 2003, 163, 2247–2257. [Google Scholar] [CrossRef]
- Shin, J.W.; Jurisic, G.; Detmar, M. Lymphatic-specific expression of dipeptidyl peptidase IV and its dual role in lymphatic endothelial function. Exp. Cell Res. 2008, 314, 3048–3056. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| X-ray/ADSC | Day 0 | Day 2 | Day 8 | Day 14 |
|---|---|---|---|---|
| −/− | 2.42 ± 0.26 | 4.25 ± 0.41 ** | 7.85 ± 0.69 **,†† | 4.77 ± 0.58 **,‡‡ |
| +/− | 2.21 ± 0.22 | 4.64 ± 0.55 ** | 3.51 ± 0.50 | 4.53 ± 0.90 ** |
| +/+ | 1.74 ± 0.24 | 3.49 ± 0.46 * | 4.81 ± 0.60 ** | 5.76 ± 0.74 ** |
| X-ray/ADSC | Day 0 | Day 2 | Day 8 | Day 14 |
|---|---|---|---|---|
| −/− | 0.25 ± 0.07 | 0.15 ± 0.06 | 0.31 ± 0.06 | 0.45 ± 0.09 |
| +/− | 0.01 ± 0.01 | 0.24 ± 0.04 ** | 0.14 ± 0.06 | 0.27 ± 0.04 ** |
| +/+ | 0.08 ± 0.03 | 0.26 ± 0.06 * | 0.56 ± 0.05 ** | 0.61 ± 0.04 ** |
| X-ray | Day 0 | X-ray/ADSC | Day 14 | ||
|---|---|---|---|---|---|
| − | Red | 21.95 ± 0.63 | −/− | Red | 19.73 ± 0.83 * |
| Green | 2.12 ± 0.16 | Green | 0.90 ± 0.14 * | ||
| + | Red | 25.42 ± 0.74 | +/− | Red | 15.06 ± 0.33 ** |
| Green | 0.43 ± 0.06 ** | ||||
| Green | 1.32 ± 0.10 | +/+ | Red | 24.68 ± 1.33 | |
| Green | 1.34 ± 0.20 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogino, R.; Hayashida, K.; Yamakawa, S.; Morita, E. Adipose-Derived Stem Cells Promote Intussusceptive Lymphangiogenesis by Restricting Dermal Fibrosis in Irradiated Tissue of Mice. Int. J. Mol. Sci. 2020, 21, 3885. https://doi.org/10.3390/ijms21113885
Ogino R, Hayashida K, Yamakawa S, Morita E. Adipose-Derived Stem Cells Promote Intussusceptive Lymphangiogenesis by Restricting Dermal Fibrosis in Irradiated Tissue of Mice. International Journal of Molecular Sciences. 2020; 21(11):3885. https://doi.org/10.3390/ijms21113885
Chicago/Turabian StyleOgino, Ryohei, Kenji Hayashida, Sho Yamakawa, and Eishin Morita. 2020. "Adipose-Derived Stem Cells Promote Intussusceptive Lymphangiogenesis by Restricting Dermal Fibrosis in Irradiated Tissue of Mice" International Journal of Molecular Sciences 21, no. 11: 3885. https://doi.org/10.3390/ijms21113885