Syringaresinol Inhibits UVA-Induced MMP-1 Expression by Suppression of MAPK/AP-1 Signaling in HaCaT Keratinocytes and Human Dermal Fibroblasts


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
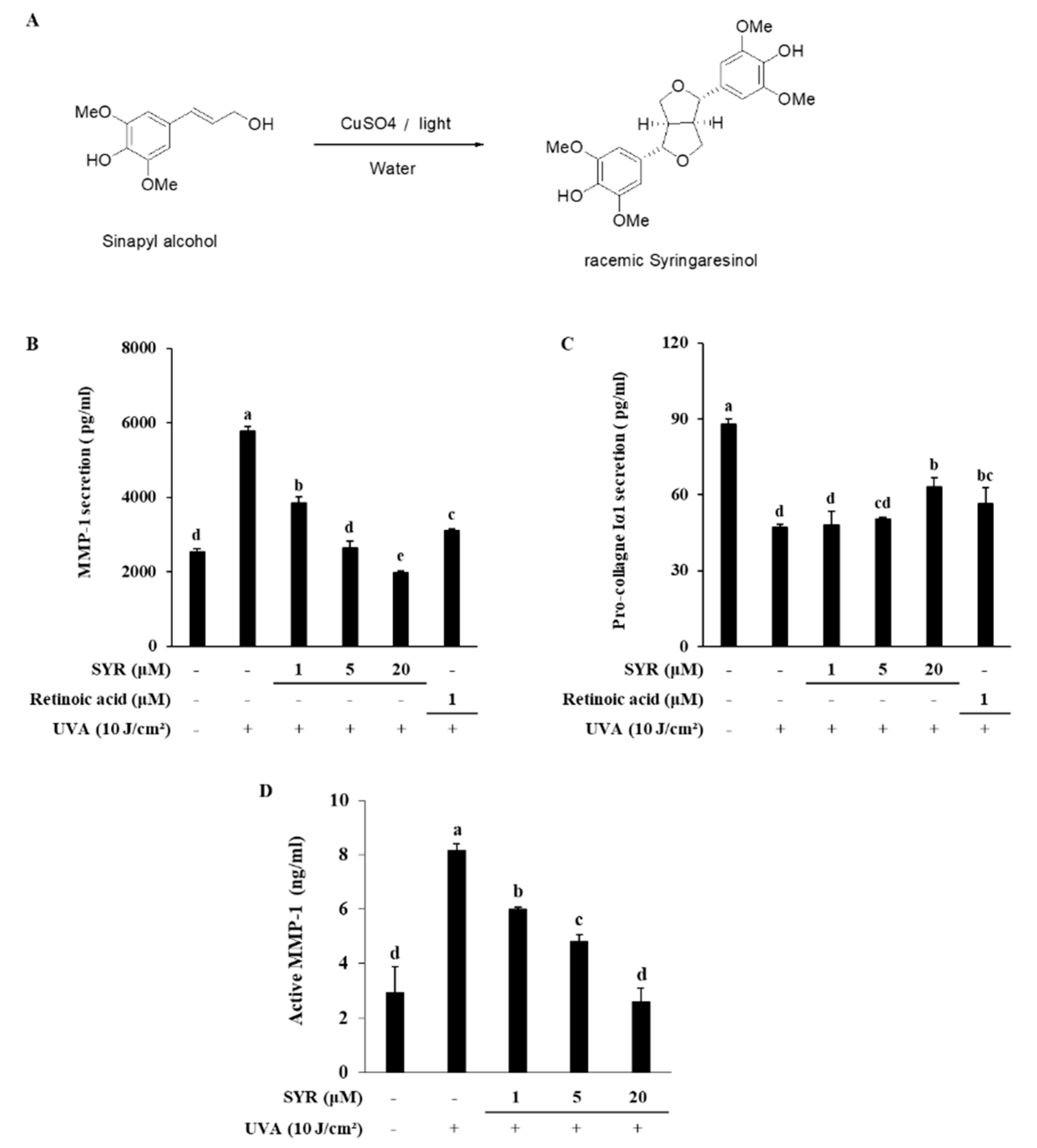
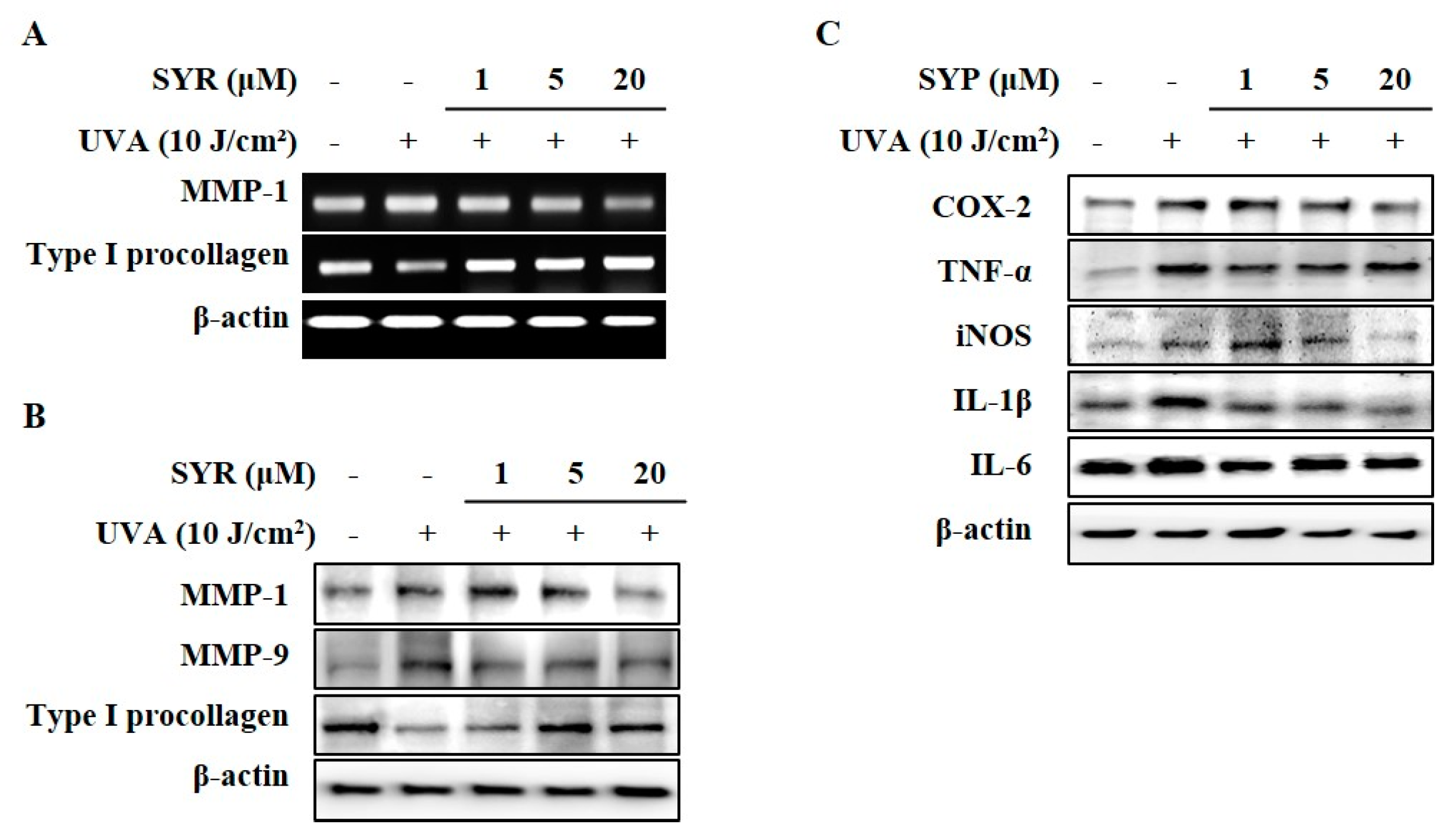
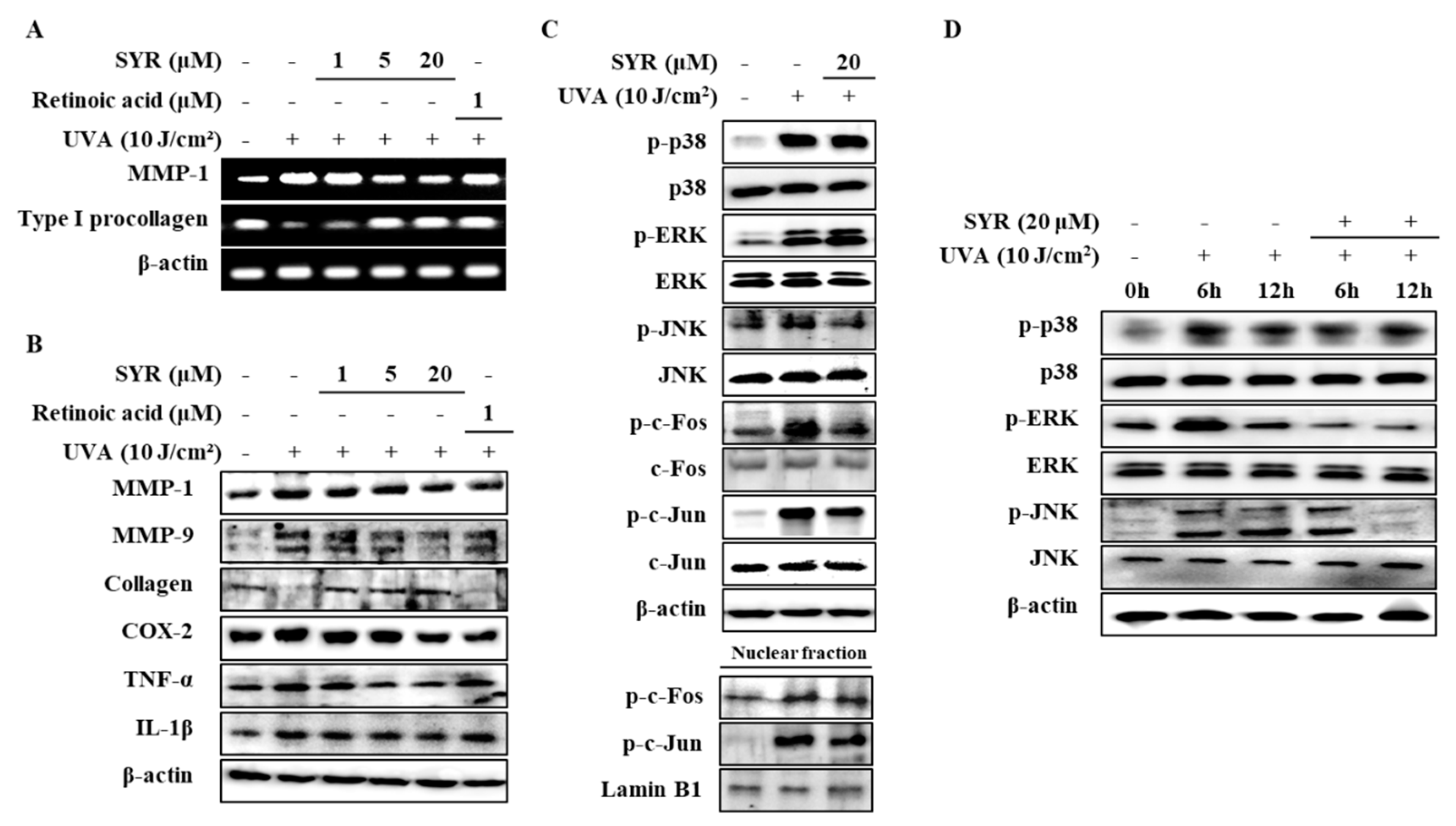
2.1. SYR Reversed UVA-Induced Changes of MMP-1 and Procollagen Iα1 Production in HaCaT Keratinocytes
2.2. UVA-Induced Inflammation in HaCaT Keratinocytes is Ameliorated by SYR Treatment
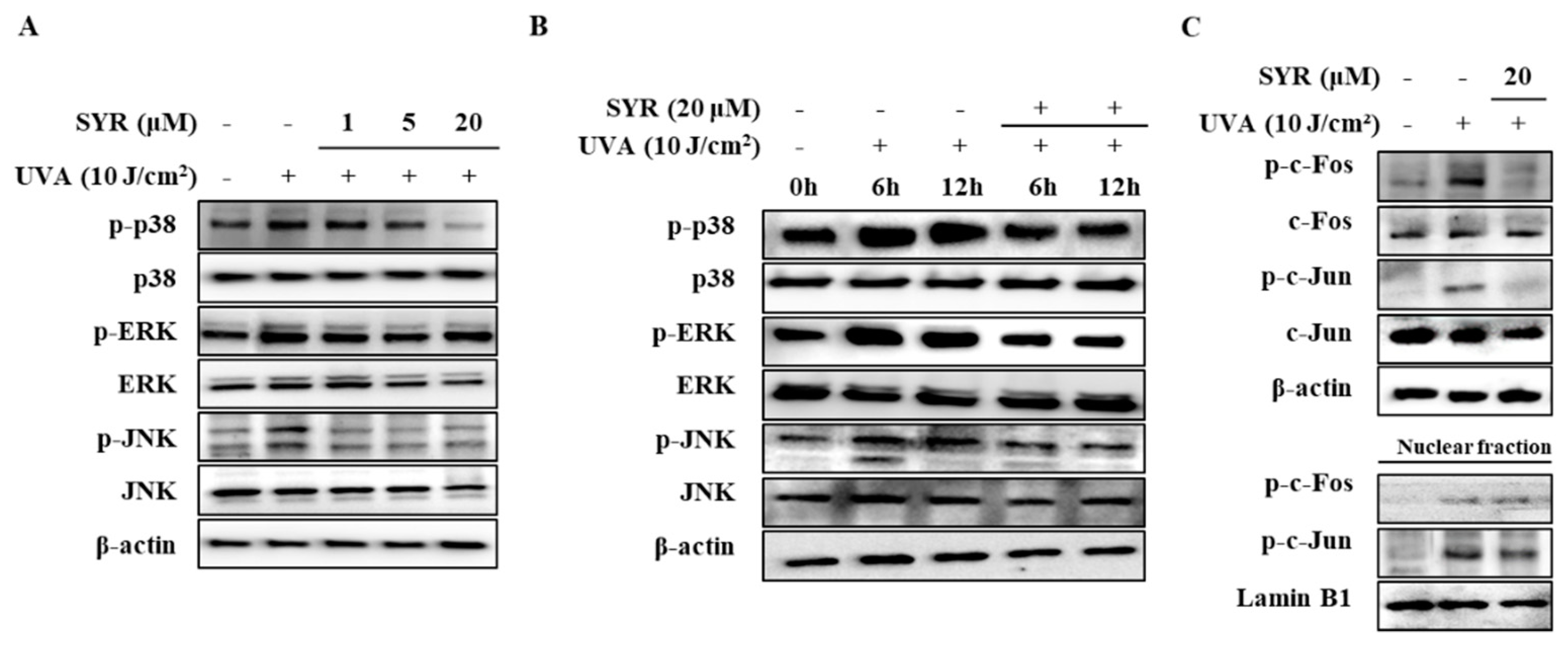
2.3. SYR Suppresses the UVA-Induced Activation of MAPK/AP-1 Signaling in Keratinocytes
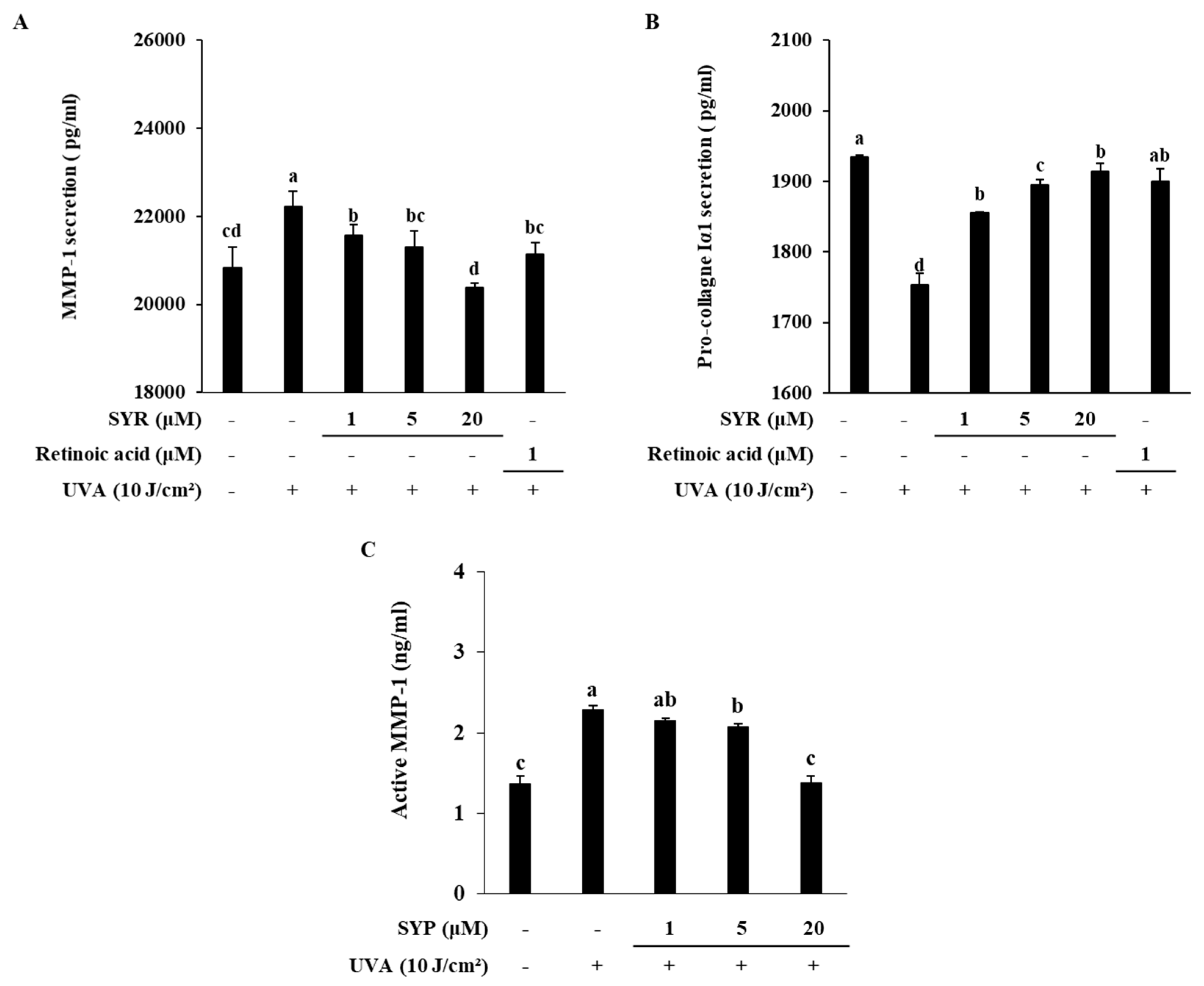
2.4. SYR Inhibits the UVA-Induced MMP-1 Production in Human Dermal Fibroblasts (HDFs) by Inhibiting MAPK-Mediated Pro-Inflammatory Response and AP-1 Activation
3. Discussion
4. Materials and Methods
4.1. Synthesis of (±)-Syringaresinol (SYR)
4.2. Cell Culture
4.3. UVA Irradiation of HaCaT Keratinocytes and HDFs
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Determination of MMP-1 Activation
4.6. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.7. Western Blotting
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rittié, L.; Fisher, G.J. UV-light-induced signal cascades and skin aging. Ageing Res. Rev. 2002, 1, 705–720. [Google Scholar] [CrossRef]
- Fisher, G.J.; Kang, S.; Varani, J.; Bata-Csorgo, Z.; Wan, Y.; Datta, S.; Voorhees, J.J. Mechanisms of photoaging and chronological skin aging. Arch. Dermatol. 2002, 138, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The skin aging exposome. J. Dermatol. Sci. 2017, 85, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, R.P.; Lee, T.K. Adverse effects of ultraviolet radiation: A brief review. Prog. Biophys. Mol. Biol. 2006, 92, 119–131. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How UV light touches the brain and endocrine system through skin, and why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef] [PubMed]
- Bocheva, G.; Slominski, R.M.; Slominski, A.T. Neuroendocrine aspects of skin aging. Int. J. Mol. Sci. 2019, 20, 2798. [Google Scholar] [CrossRef] [PubMed]
- Yaar, M.; Gilchrest, B.A. Photoageing: Mechanism, prevention and therapy. Br. J. Dermatol. 2007, 157, 874–887. [Google Scholar] [CrossRef]
- Rabe, J.H.; Mamelak, A.J.; McElgunn, P.J.S.; Morison, W.L.; Sauder, D.N. Photoaging: Mechanisms and repair. J. Am. Acad. Dermatol. 2006, 55, 1–19. [Google Scholar] [CrossRef]
- Silvers, A.L.; Bachelor, M.A.; Bowden, G.T. The role of JNK and p38 MAPK activities in UVA-induced signaling pathways leading to AP-1 activation and c-Fos expression. Neoplasia 2003, 5, 319–329. [Google Scholar] [CrossRef]
- Pardo, A.; Selman, M. MMP-1: The elder of the family. Int. J. Biochem. Cell Biol. 2005, 37, 283–288. [Google Scholar] [CrossRef]
- Altemimi, A.; Lakhssassi, N.; Baharlouei, A.; Watson, D.; Lightfoot, D. Phytochemicals: Extraction, isolation, and identification of bioactive compounds from plant extracts. Plants 2017, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Teponno, R.B.; Kusari, S.; Spiteller, M. Recent advances in research on lignans and neolignans. Nat. Prod. Rep. 2016, 33, 1044–1092. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-G.; Watanabe, N.; Sano, Y.; Uraki, Y.; Sano, Y.O. Extractives of kitakobusi Magnolia kobus DC. var. borealis Sarg. III.: Antibacterial and antifungal activity of extractives. Bull. Hokkaido Univ. For. 1998, 55, 63–73. [Google Scholar]
- Salleh, W.M.N.H.W.; Ahmad, F.; Heng Yen, K.; Zulkifli, R.M. Anticholinesterase and anti-inflammatory constituents from Beilschmiedia pulverulenta kosterm. Nat. Prod. Sci. 2016, 22, 225–230. [Google Scholar] [CrossRef]
- Wang, H.M.; Chou, Y.T.; Hong, Z.L.; Chen, H.A.; Chang, Y.C.; Yang, W.L.; Chang, H.C.; Mai, C.T.; Chen, C.Y. Bioconstituents from stems of Synsepalum dulcificum Daniell (Sapotaceae) inhibit human melanoma proliferation, reduce mushroom tyrosinase activity and have antioxidant properties. J. Taiwan Inst. Chem. Eng. 2011, 42, 204–211. [Google Scholar] [CrossRef]
- Honda, A.; Abe, R.; Makino, T.; Norisugi, O.; Fujita, Y.; Watanabe, H.; Nishihira, J.; Iwakura, Y.; Yamagishi, S.-I.; Shimizu, H.; et al. Interleukin-1β and macrophage migration inhibitory factor (MIF) in dermal fibroblasts mediate UVA-induced matrix metalloproteinase-1 expression. J. Dermatol. Sci. 2008, 49, 63–72. [Google Scholar] [CrossRef]
- Muthusamy, V.; Piva, T.J. The UV response of the skin: A review of the MAPK, NFκB and TNFα signal transduction pathways. Arch. Dermatol. Res. 2010, 302, 5–17. [Google Scholar] [CrossRef]
- Benbow, U.; Brinckerhoff, C.E. The AP-1 site and MMP gene regulation: What is all the fuss about? Matrix Biol. 1997, 15, 519–526. [Google Scholar] [CrossRef]
- de Jager, T.L.; Cockrell, A.E.; Du Plessis, S.S. Ultraviolet light induced generation of reactive oxygen species. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2017; Volume 996, pp. 15–23. [Google Scholar]
- Lee, B.; Moon, K.M.; Son, S.; Yun, H.Y.; Han, Y.K.; Ha, Y.M.; Kim, D.H.; Chung, K.W.; Lee, E.K.; An, H.J.; et al. (2R/S,4R)-2-(2,4-Dihydroxyphenyl)thiazolidine-4-carboxylic acid prevents UV-induced wrinkle formation through inhibiting NF-κB-mediated inflammation. J. Dermatol. Sci. 2015, 79, 313–316. [Google Scholar] [CrossRef]
- Yang, Y.; Kim, H.J.; Woo, K.-J.; Cho, D.; Bang, S.I. Lipo-PGE1 suppresses collagen production in human dermal fibroblasts via the ERK/Ets-1 signaling pathway. PLoS ONE 2017, 12, e0179614. [Google Scholar] [CrossRef]
- Bhogal, R.K.; Bona, C.A. Regulatory effect of extracellular signal-regulated kinases (ERK) on type I collagen synthesis in human dermal fibroblasts stimulated by IL-4 and IL-13. Int. Rev. Immunol. 2008, 27, 472–496. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-García, C.; Sánchez-Quesada, C.; Toledo, E.; Delgado-Rodríguez, M.; Gaforio, J.J. Naturally lignan-rich foods: A dietary tool for health promotion? Molecules 2019, 24, 917. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Hu, X. Advances in the synthesis of lignan natural products. Molecules 2018, 23, 3385. [Google Scholar] [CrossRef]
- Kim, J.; Cho, S.Y.; Kim, S.H.; Cho, D.; Kim, S.; Park, C.W.; Shimizu, T.; Cho, J.Y.; Seo, D.B.; Shin, S.S. Effects of Korean ginseng berry on skin antipigmentation and antiaging via FoxO3a activation. J. Ginseng Res. 2017, 41, 277–283. [Google Scholar] [CrossRef]
- Kim, J.; Toda, T.; Watanabe, K.; Shibuya, S.; Ozawa, Y.; Izuo, N.; Cho, S.; Seo, D.B.; Yokote, K.; Shimizu, T.J. Syringaresinol Reverses Age-Related Skin Atrophy by Suppressing FoxO3a-Mediated Matrix Metalloproteinase—2 Activation in Copper/Zinc Superoxide Dismutase—Deficient Mice. J. Investig. Dermatol. 2019, 139, 648–655. [Google Scholar]
- Kun, N.; Tsuchiya, K.; Nakajima, S. New anti-aging cosmetic ingredients. J. Soc. Cosmet. Chem. Jpn. 1999, 33, 245–253. [Google Scholar] [CrossRef]
- Oh, J.H.; Seo, Y.; Kong, C. Anti-photoaging effects of solvent-partitioned fractions from Portulaca oleracea L. on UVB-stressed human keratinocytes. J. Food Biochem. 2019, 43, e12814. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, J.H.; Joo, Y.H.; Karadeniz, F.; Ko, J.; Kong, C.-S. Syringaresinol Inhibits UVA-Induced MMP-1 Expression by Suppression of MAPK/AP-1 Signaling in HaCaT Keratinocytes and Human Dermal Fibroblasts. Int. J. Mol. Sci. 2020, 21, 3981. https://doi.org/10.3390/ijms21113981
Oh JH, Joo YH, Karadeniz F, Ko J, Kong C-S. Syringaresinol Inhibits UVA-Induced MMP-1 Expression by Suppression of MAPK/AP-1 Signaling in HaCaT Keratinocytes and Human Dermal Fibroblasts. International Journal of Molecular Sciences. 2020; 21(11):3981. https://doi.org/10.3390/ijms21113981
Chicago/Turabian StyleOh, Jung Hwan, Yung Hyup Joo, Fatih Karadeniz, Jaeyoung Ko, and Chang-Suk Kong. 2020. "Syringaresinol Inhibits UVA-Induced MMP-1 Expression by Suppression of MAPK/AP-1 Signaling in HaCaT Keratinocytes and Human Dermal Fibroblasts" International Journal of Molecular Sciences 21, no. 11: 3981. https://doi.org/10.3390/ijms21113981
APA StyleOh, J. H., Joo, Y. H., Karadeniz, F., Ko, J., & Kong, C.-S. (2020). Syringaresinol Inhibits UVA-Induced MMP-1 Expression by Suppression of MAPK/AP-1 Signaling in HaCaT Keratinocytes and Human Dermal Fibroblasts. International Journal of Molecular Sciences, 21(11), 3981. https://doi.org/10.3390/ijms21113981