Physiological Importance of Pectin Modifying Genes During Rice Pollen Development

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Effects of a PME and PME-Inhibiting Catechin Extract on In Vitro Pollen Germination in Rice
2.2. Alteration of Immunolabeling Patterns of Pectins by Exogenous Treatment of PME and Polyphenon 60
2.3. Identification and In Silico Characterization of Rice Pollen-Specific PME and PMEI Genes
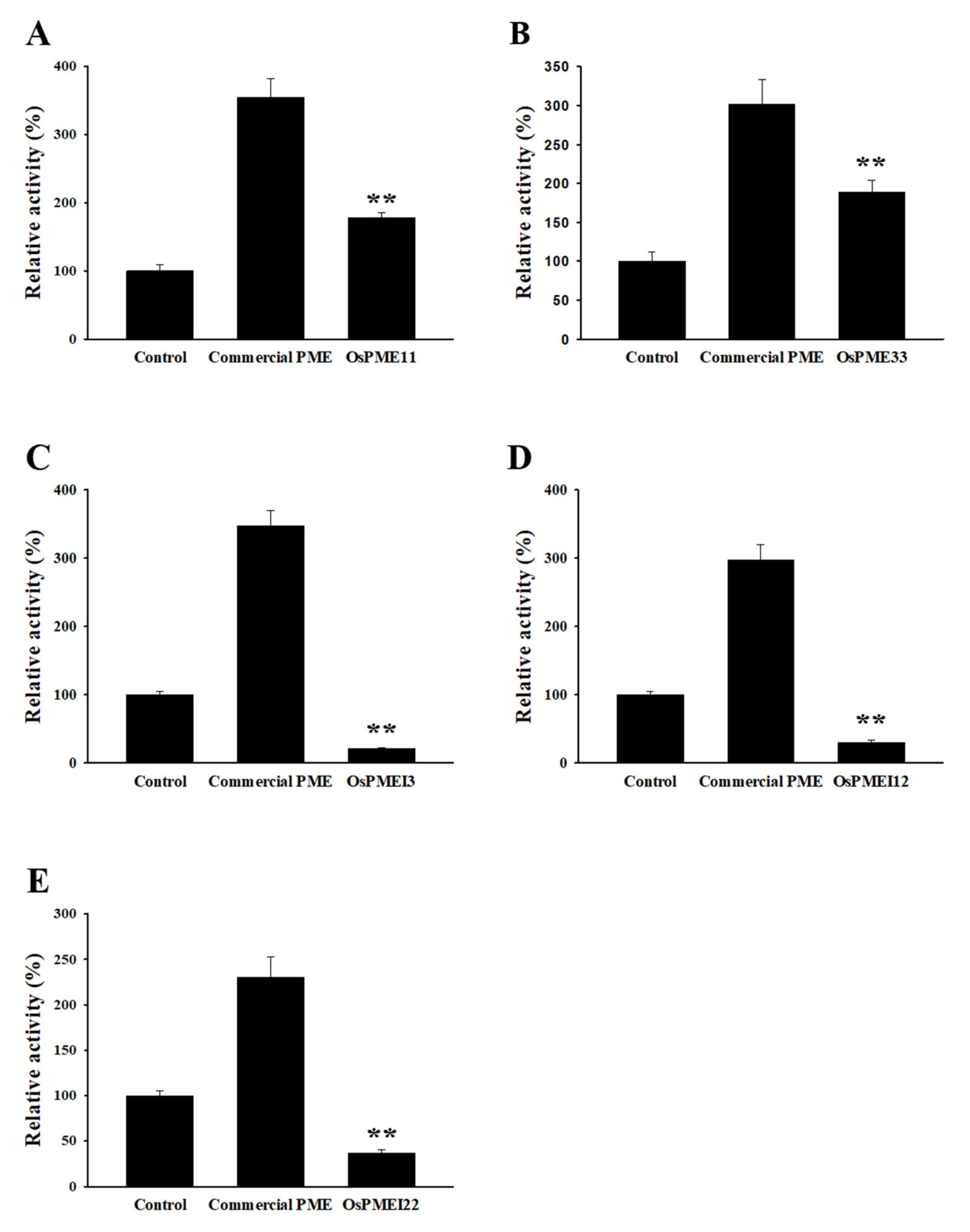
2.4. Enzyme Activity Assay Using the Tobacco Infiltration System
2.5. Subcellular Localization of Rice Pollen Specific PMEs and PMEIs
3. Discussion
3.1. Fine-Tuning of PME and PMEI Activities is Required for the Proper Development of Pollen Tubes
3.2. Several PME and PMEI Genes are Specifically Expressed in the Late Stage of Pollen Development in Rice
3.3. Pollen-Specific PMEs and PMEIs Possess Unique Enzymatic Activities in Tobacco Leaves
4. Materials and Methods
4.1. Plant Growth and In Vitro Pollen Germination
4.2. Immunolocalization of Pollen Tubes
4.3. Genome-Wide Phylogenomic Analysis
4.4. Tissue Expression Analysis
4.5. Vector Construction and Subcellular Localization Analysis
4.6. PME and PMEI Activity Assay
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| HGA | Homogalacturonan |
| PME | Pectin methylesterase |
| PMEI | Pectin methylesterase inhibitor |
| GalUA | galacturonic acid |
References
- Rounds, C.M.; Bezanilla, M. Growth Mechanisms in Tip-Growing Plant Cells. Annu. Rev. Plant Biol. 2013, 64, 243–265. [Google Scholar] [CrossRef]
- Offringa, R.; Kleine-Vehn, J. Cell Polarity and Development. J. Integr. Plant Biol. 2013, 55, 786–788. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Zhang, D.; Jung, K.-H. Molecular Basis of Pollen Germination in Cereals. Trends Plant Sci. 2019, 24, 1126–1136. [Google Scholar] [CrossRef]
- Minic, Z.; Jouanin, L. Plant glycoside hydrolases involved in cell wall polysaccharide degradation. Plant Physiol. Biochem. 2006, 44, 435–449. [Google Scholar] [CrossRef]
- Oh, C.-S.; Kim, H.; Lee, C. Rice cell wall polysaccharides: Structure and biosynthesis. J. Plant Biol. 2013, 56, 274–282. [Google Scholar] [CrossRef]
- Mohnen, D. Pectin structure and biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.Y.; Nguyen, H.P.; Lee, C. Genome-wide identification and expression analysis of rice pectin methylesterases: Implication of functional roles of pectin modification in rice physiology. J. Plant Physiol. 2015, 183, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J. Unique aspects of the grass cell wall. Curr. Opin. Plant Biol. 2008, 11, 301–307. [Google Scholar] [CrossRef]
- Harholt, J.; Suttangkakul, A.; Scheller, H.V. Biosynthesis of Pectin1. Plant Physiol. 2010, 153, 384–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelloux, J.; Rusterucci, C.; Mellerowicz, E. New insights into pectin methylesterase structure and function. Trends Plant Sci. 2007, 12, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-Q.; Ye, D. Roles of Pectin Methylesterases in Pollen-Tube Growth. J. Integr. Plant Biol. 2007, 49, 94–98. [Google Scholar] [CrossRef]
- Wallace, S.; Williams, J. Evolutionary origins of pectin methylesterase genes associated with novel aspects of angiosperm pollen tube walls. Biochem. Biophys. Res. Commun. 2017, 487, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Rausch, T.; Greiner, S. The N-terminal pro region mediates retention of unprocessed type-I PME in the Golgi apparatus. Plant J. 2009, 58, 361–375. [Google Scholar] [CrossRef]
- Di Matteo, A.; Giovane, A.; Raiola, A.; Camardella, L.; Bonivento, D.; De Lorenzo, G.; Cervone, F.; Bellincampi, D.; Tsernoglou, D. Structural Basis for the Interaction between Pectin Methylesterase and a Specific Inhibitor Protein. Plant Cell 2005, 17, 849–858. [Google Scholar] [CrossRef] [Green Version]
- Hocq, L.; Sénéchal, F.; Lefebvre, V.; Lehner, A.; Domon, J.-M.; Mollet, J.-C.; Dehors, J.; Pageau, K.; Marcelo, P.; Guerineau, F.; et al. Combined Experimental and Computational Approaches Reveal Distinct pH Dependence of Pectin Methylesterase Inhibitors1. Plant Physiol. 2016, 173, 1075–1093. [Google Scholar] [CrossRef] [Green Version]
- Rãckel, N.; Wolf, S.; Kost, B.; Rausch, T.; Greiner, S.; Röckel, N. Elaborate spatial patterning of cell-wall PME and PMEI at the pollen tube tip involves PMEI endocytosis, and reflects the distribution of esterified and de-esterified pectins. Plant J. 2008, 53, 133–143. [Google Scholar] [CrossRef]
- Wormit, A.; Usadel, B. The Multifaceted Role of Pectin Methylesterase Inhibitors (PMEIs). Int. J. Mol. Sci. 2018, 19, 2878. [Google Scholar] [CrossRef] [Green Version]
- Grebnev, G.; Ntefidou, M.; Kost, B. Secretion and Endocytosis in Pollen Tubes: Models of Tip Growth in the Spot Light. Front. Plant Sci. 2017, 8, 1055. [Google Scholar] [CrossRef]
- Parre, E.; Geitmann, A. Pectin and the role of the physical properties of the cell wall in pollen tube growth of Solanum chacoense. Planta 2004, 220, 582–592. [Google Scholar] [CrossRef]
- Nguyen, H.P.; Jeong, H.Y.; Kim, H.; Kim, Y.C.; Lee, C. Molecular and biochemical characterization of rice pectin methylesterase inhibitors (OsPMEIs). Plant Physiol. Biochem. 2016, 101, 105–112. [Google Scholar] [CrossRef]
- Szumlanski, A.L.; Nielsen, E. The Rab GTPase RabA4d Regulates Pollen Tube Tip Growth in Arabidopsis thaliana[W]. Plant Cell 2009, 21, 526–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, M.; Cheung, A.Y.; Hepler, P.K. Pectin Methylesterase, a Regulator of Pollen Tube Growth1[w]. Plant Physiol. 2005, 138, 1334–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, K.E.; Lam, S.Y.; Copenhaver, G.P. Separation of Arabidopsis Pollen Tetrads Is Regulated by QUARTET1, a Pectin Methylesterase Gene1. Plant Physiol. 2006, 142, 1004–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, K.C.; Selzer, T.; Shahar, C.; Udi, Y.; Tworowski, D.; Sagi, I. Inhibition of pectin methyl esterase activity by green tea catechins. Phytochemistry 2008, 69, 2586–2592. [Google Scholar] [CrossRef] [Green Version]
- Schoenaers, S.; Balcerowicz, D.; Breen, G.; Hill, K.; Zdanio, M.; Mouille, G.; Holman, T.J.; Oh, J.; Wilson, M.; Nikonorova, N.; et al. The Auxin-Regulated CrRLK1L Kinase ERULUS Controls Cell Wall Composition during Root Hair Tip Growth. Curr. Biol. 2018, 28, 722–732. [Google Scholar] [CrossRef] [Green Version]
- Hong, G.; Wang, J.; Hochstetter, D.; Gao, Y.; Xu, P.; Wang, Y. Epigallocatechin-3-gallate functions as a physiological regulator by modulating the jasmonic acid pathway. Physiol. Plant. 2014, 153, 432–439. [Google Scholar] [CrossRef]
- Moon, S.; Oo, M.M.; Kim, B.; Koh, H.-J.; Oh, S.A.; Yi, G.; An, G.; Park, S.K.; Jung, K.-H. Genome-wide analyses of late pollen-preferred genes conserved in various rice cultivars and functional identification of a gene involved in the key processes of late pollen development. Rice 2018, 11, 28. [Google Scholar] [CrossRef] [Green Version]
- Hongo, S.; Sato, K.; Yokoyama, R.; Nishitani, K. Demethylesterification of the Primary Wall by PECTIN METHYLESTERASE35 Provides Mechanical Support to the Arabidopsis Stem. Plant Cell 2012, 24, 2624–2634. [Google Scholar] [CrossRef] [Green Version]
- Jolie, R.P.; Duvetter, T.; Van Loey, A.; Hendrickx, M. Pectin methylesterase and its proteinaceous inhibitor: A review. Carbohydr. Res. 2010, 345, 2583–2595. [Google Scholar] [CrossRef]
- Saez-Aguayo, S.; Ralet, M.-C.; Berger, A.; Botran, L.; Ropartz, D.; Marion-Poll, A.; North, H.M. PECTIN METHYLESTERASE INHIBITOR6 Promotes Arabidopsis Mucilage Release by Limiting Methylesterification of Homogalacturonan in Seed Coat Epidermal Cells. Plant Cell 2013, 25, 308–323. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.A.; Harper, J.F.; Palanivelu, R. A Fruitful Journey: Pollen Tube Navigation from Germination to Fertilization. Annu. Rev. Plant Biol. 2019, 70, 809–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woriedh, M.; Wolf, S.; Márton, M.L.; Hinze, A.; Gahrtz, M.; Becker, D.; Dresselhaus, T. External application of gametophyte-specific ZmPMEI1 induces pollen tube burst in maize. Plant Reprod. 2013, 26, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Gou, J.-Y.; Miller, L.M.; Hou, G.; Yu, X.-H.; Chen, X.; Liu, C.-J. Acetylesterase-Mediated Deacetylation of Pectin Impairs Cell Elongation, Pollen Germination, and Plant Reproduction. Plant Cell 2012, 24, 50–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, S.Y.; Osborne, E.; Poindexter, P.D.; Somerville, C.R. Microspore Separation in the quartet 3 Mutants of Arabidopsis Is Impaired by a Defect in a Developmentally Regulated Polygalacturonase Required for Pollen Mother Cell Wall Degradation1. Plant Physiol. 2003, 133, 1170–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Yang, X.; Zhang, X.; Song, Y.; Liang, W.; Zhang, D. Rice Morphology Determinant-Mediated Actin Filament Organization Contributes to Pollen Tube Growth. Plant Physiol. 2018, 177, 255–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef]
- Grsic-Rausch, S.; Rausch, T. A coupled spectrophotometric enzyme assay for the determination of pectin methylesterase activity and its inhibition by proteinaceous inhibitors. Anal. Biochem. 2004, 333, 14–18. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.-J.; Jeong, H.Y.; Kang, S.-Y.; Silva, J.; Kim, E.-J.; Park, S.K.; Jung, K.-H.; Lee, C. Physiological Importance of Pectin Modifying Genes During Rice Pollen Development. Int. J. Mol. Sci. 2020, 21, 4840. https://doi.org/10.3390/ijms21144840
Kim Y-J, Jeong HY, Kang S-Y, Silva J, Kim E-J, Park SK, Jung K-H, Lee C. Physiological Importance of Pectin Modifying Genes During Rice Pollen Development. International Journal of Molecular Sciences. 2020; 21(14):4840. https://doi.org/10.3390/ijms21144840
Chicago/Turabian StyleKim, Yu-Jin, Ho Young Jeong, Seung-Yeon Kang, Jeniffer Silva, Eui-Jung Kim, Soon Ki Park, Ki-Hong Jung, and Chanhui Lee. 2020. "Physiological Importance of Pectin Modifying Genes During Rice Pollen Development" International Journal of Molecular Sciences 21, no. 14: 4840. https://doi.org/10.3390/ijms21144840
APA StyleKim, Y.-J., Jeong, H. Y., Kang, S.-Y., Silva, J., Kim, E.-J., Park, S. K., Jung, K.-H., & Lee, C. (2020). Physiological Importance of Pectin Modifying Genes During Rice Pollen Development. International Journal of Molecular Sciences, 21(14), 4840. https://doi.org/10.3390/ijms21144840