Structural Study of Metal Binding and Coordination in Ancient Metallo-?-Lactamase PNGM-1 Variants

 and
and
Abstract
:1. Introduction
2. Results
2.1. Two Metal-Binding Sites in PNGM-1
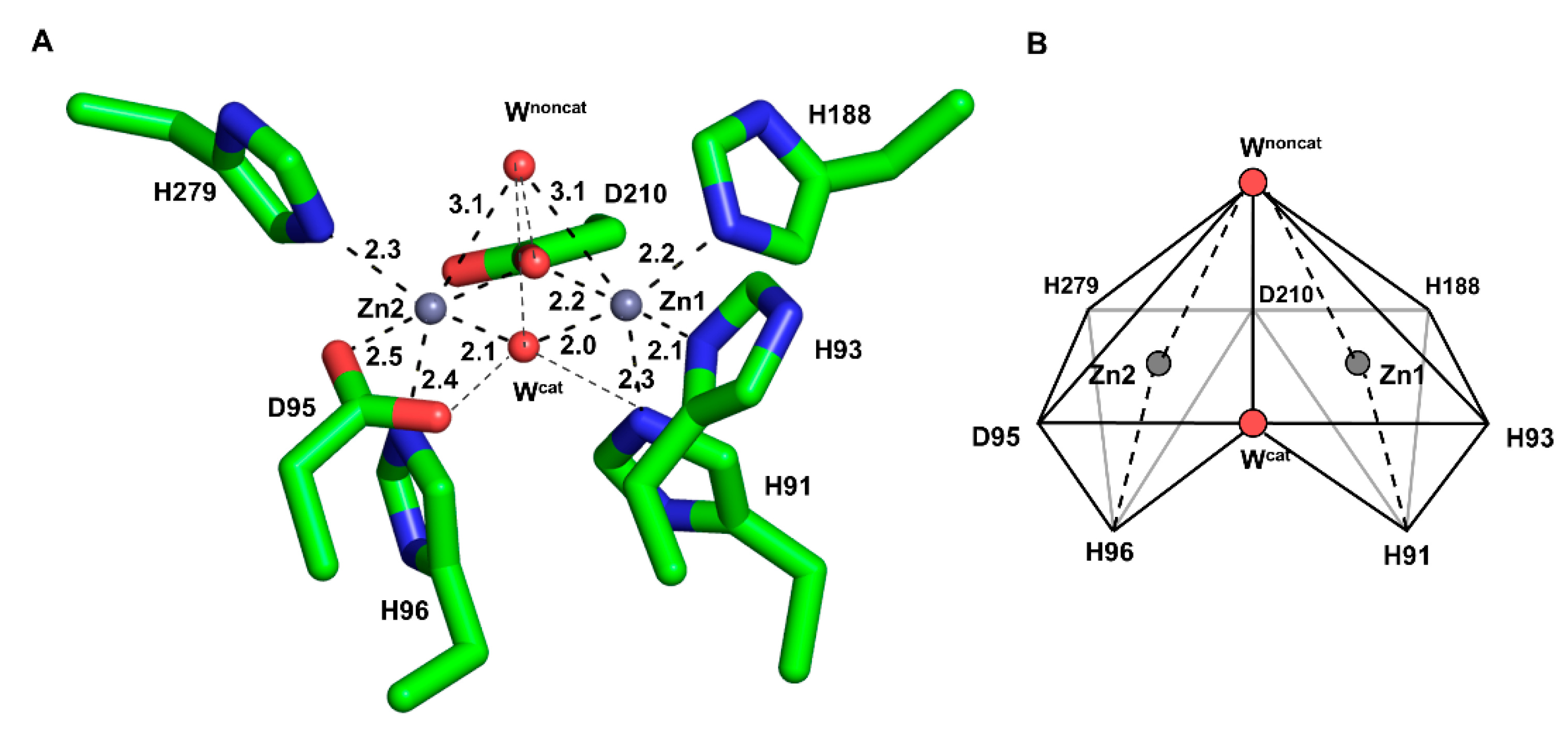
2.2. Metal Coordination Geometry of the MBSs
2.3. Metal Occupancy in Two MBSs
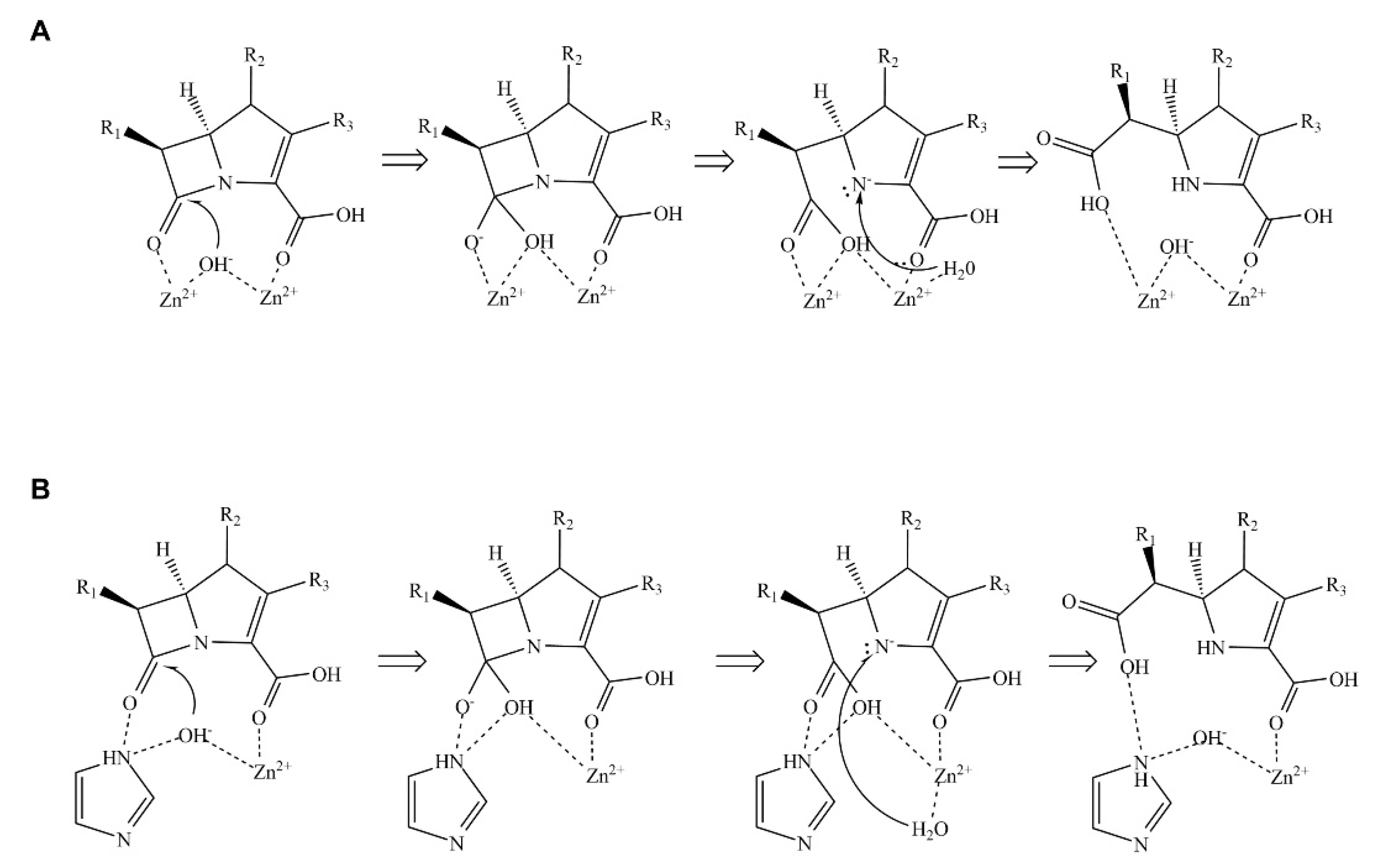
2.4. Catalytic Water Molecule between the Two Zinc Ions
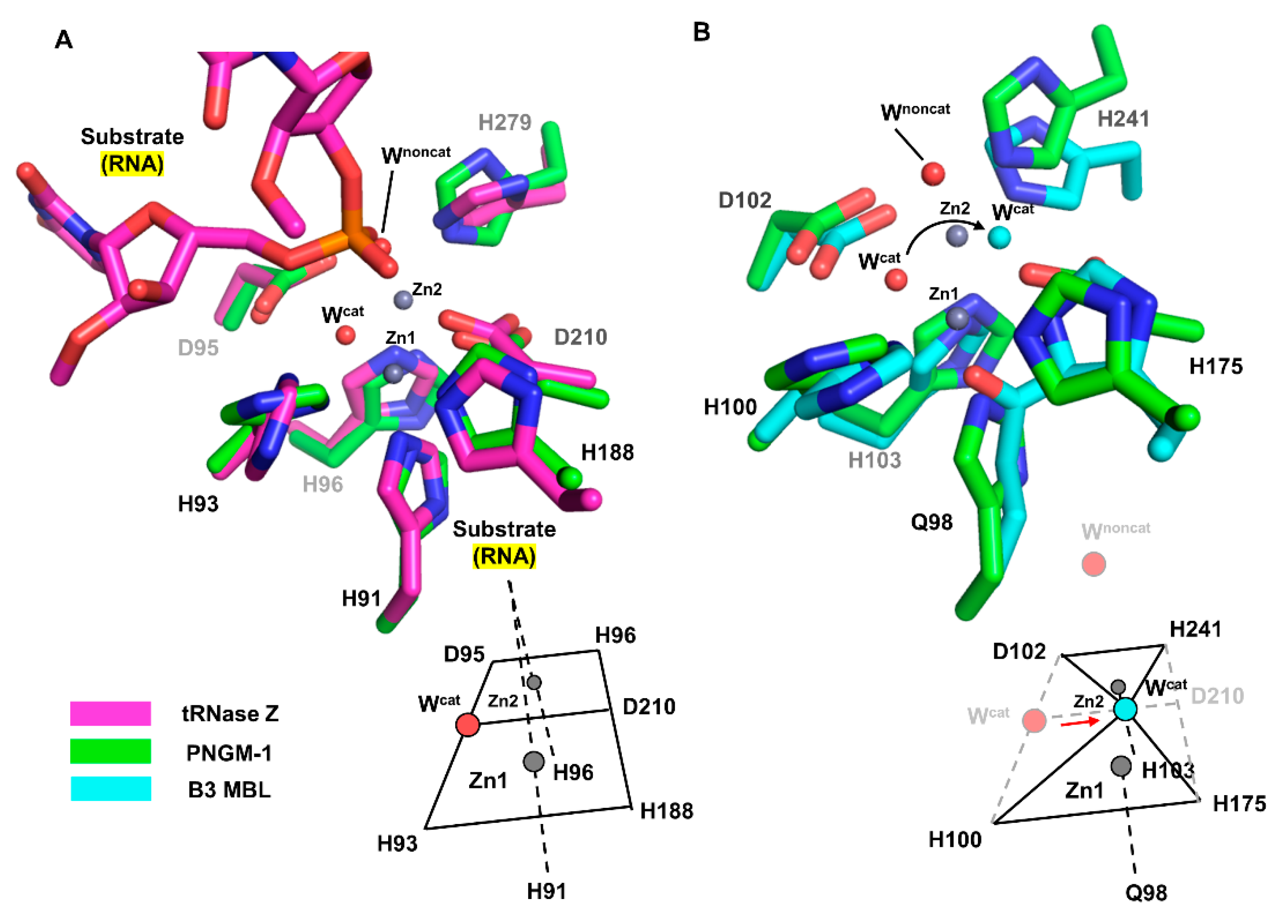
2.5. Structural Comparisons between PNGM-1 and Other B3 MBLs
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Gene Cloning and Site-Directed Mutagenesis
4.3. Expression and Purification of PNGM-1 Variants
4.4. Crystallization
4.5. Data Collection, Structure Determination, and Refinement
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MDR | Multidrug-resistant |
| MBL | Metallo-β-lactamase |
| PBP | Penicillin-binding protein |
| MBS | Metal-binding site |
| Zn1 | The first metal-binding site |
| Zn2 | The second metal-binding site |
| Wcat | Catalytic water |
| Wnoncat | Non-catalytic water |
References
- Hughes, D. Exploiting genomics, genetics and chemistry to combat antibiotic resistance. Nat. Rev. Genet. 2003, 4, 432–441. [Google Scholar] [CrossRef]
- Richter, M.F.; Hergenrother, P.J. The challenge of converting Gram-positive-only compounds into broad-spectrum antibiotics. Ann. N. Y. Acad. Sci. 2018, 1435, 18–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornejo-Juárez, P.; Vilar-Compte, D.; Pérez-Jiménez, C.; Ñamendys-Silva, S.A.; Sandoval-Hernández, S.; Volkow-Fernández, P. The impact of hospital-acquired infections with multidrug-resistant bacteria in an oncology intensive care unit. Int. J. Infect. Dis. 2015, 31, 31–34. [Google Scholar] [CrossRef] [Green Version]
- De Kraker, M.E.; Stewardson, A.J.; Harbarth, S. Will 10 Million People Die a Year due to Antimicrobial Resistance by 2050? PLoS Med. 2016, 13, e1002184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.-H.; Li, G.; Li, G.-B. Principles and current strategies targeting metallo-beta-lactamase mediated antibacterial resistance. Med. Res. Rev. 2020. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.M.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Queenan, A.M.; Bush, K. Carbapenemases: The Versatile beta-Lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Fast, W.; Benkovic, S.J. On the Mechanism of the Metallo-beta-lactamase from Bacteroides fragilis. Biochemestry 1999, 38, 10013–10023. [Google Scholar] [CrossRef]
- Brem, J.; Cain, R.; Cahill, S.T.; McDonough, M.A.; Clifton, I.J.; Jimenez-Castellanos, J.-C.; Avison, M.B.; Spencer, J.; Fishwick, C.W.; Schofield, C.J. Structural basis of metallo-beta-lactamase, serine-beta-lactamase and penicillin-binding protein inhibition by cyclic boronates. Nat. Commun. 2016, 7, 12406. [Google Scholar] [CrossRef] [Green Version]
- Orellano, E.G.; Girardini, J.E.; Cricco, J.A.; Ceccarelli, E.A.; Vila, A.J. Spectroscopic Characterization of a Binuclear Metal Site in Bacillus cereus beta-Lactamase II. Biochemestry 1998, 37, 10173–10180. [Google Scholar] [CrossRef]
- Wommer, S.; Rival, S.; Heinz, U.; Galleni, M.; Frère, J.-M.; Franceschini, N.; Amicosante, G.; Rasmussen, B.; Bauer, R.; Adolph, H.-W. Substrate-activated Zinc Binding of Metallo-beta-lactamases. J. Biol. Chem. 2002, 277, 24142–24147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.S.; Hong, M.-K.; Jeon, J.W.; Kim, J.H.; Jeon, J.H.; Lee, J.H.; Kim, T.Y.; Karim, A.M.; Malik, S.K.; Kang, L.-W.; et al. The novel metallo-beta-lactamase PNGM-1 from a deep-sea sediment metagenome: Crystallization and X-ray crystallographic analysis. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2018, 74, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Garau, G.; Di Guilmi, A.M.; Hall, B.G. Structure-Based Phylogeny of the Metallo-beta-Lactamases. Antimicrob. Agents Chemother. 2005, 49, 2778–2784. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Takahashi, M.; Jeon, J.H.; Kang, L.-W.; Seki, M.; Park, K.S.; Hong, M.-K.; Park, Y.S.; Kim, T.Y.; Karim, A.M.; et al. Dual activity of PNGM-1 pinpoints the evolutionary origin of subclass B3 metallo-beta-lactamases: A molecular and evolutionary study. Emerg. Microbes Infect. 2019, 8, 1688–1700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojica, M.F.; Bonomo, R.A.; Fast, W. B1-Metallo-beta-Lactamases: Where do we stand? Curr. Drug Targets 2016, 17, 1029–1050. [Google Scholar] [CrossRef]
- Pellegrini, O.; De La Sierra-Gallay, I.L.; Piton, J.; Gilet, L.; Condon, C. Activation of tRNA Maturation by Downstream Uracil Residues in B. subtilis. Structure 2012, 20, 1769–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morán-Barrio, J.; Lisa, M.-N.; Larrieux, N.; Drusin, S.I.; Viale, A.M.; Moreno, D.M.; Buschiazzo, A.; Vila, A.J. Crystal Structure of the Metallo-beta-Lactamase GOB in the Periplasmic Dizinc Form Reveals an Unusual Metal Site. Antimicrob. Agents Chemother. 2016, 60, 6013–6022. [Google Scholar] [CrossRef] [Green Version]
- Park, K.S.; Kim, T.Y.; Kim, J.H.; Lee, J.H.; Jeon, J.H.; Karim, A.M.; Malik, S.K.; Lee, S.H. PNGM-1, a novel subclass B3 metallo-beta-lactamase from a deep-sea sediment metagenome. J. Glob. Antimicrob. Resist. 2018, 14, 302–305. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar] [CrossRef]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murshudov, G.N.; Skubak, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vriend, G. WHAT IF: A molecular modeling and drug design program. J. Mol. Graph. 1990, 8, 52–56. [Google Scholar] [CrossRef]
- Brat, D.; Boles, E.; Wiedemann, B. Functional Expression of a Bacterial Xylose Isomerase in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2009, 75, 2304–2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrodinger, L. The PyMOL Molecular Graphics System Version 1.3r1. 2010. Available online: https://pymol.org/2/ (accessed on 1 July 2020).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H91A | H93A | H96A | H257A | H279A | |
|---|---|---|---|---|---|
| Data collection | |||||
| Wavelength (Å) | 0.9796 | 0.9796 | 0.9796 | 0.9796 | 0.9796 |
| Resolution range | 48.0–1.9 (2.01–1.94) | 40.3–1.6 (1.67–1.61) | 45.8–2.5 (2.54–2.45) | 48.6–2.0 (2.07–2.0) | 48.0–2.2 (2.23–2.16) |
| Space group | P 1 21 1 | P 1 21 1 | C 1 2 1 | P 1 21 1 | C 1 2 1 |
| Unit cell a, b, c (Å) α, β, γ (°) | 121.4, 83.1, 163.6 90, 110.7, 90 | 121.8, 82.8, 164.0 90, 111.1, 90 | 121.6, 82.5, 163.7 90, 110.8, 90 | 79.8, 143.7, 79.8 90, 111.8, 90 | 121.9, 83.0, 164.0 90, 110.9, 90 |
| Total reflections | 1082322 | 2483252 | 255443 | 524840 | 397308 |
| Unique reflections | 214810 (16994) | 388130 (37693) | 50474 (4821) | 109298 (10188) | 79596 (7627) |
| Multiplicity | 5.1 (4.0) | 6.4 (5.5) | 5.1 (4.7) | 4.8 (3.5) | 5.0 (3.7) |
| Completeness (%) | 95.0 (75.9) | 98.9 (96.5) | 90.2 (87.0) | 97.3 (91.5) | 96.5 (92.6) |
| Mean I/sigma (I) | 14.5 (2.3) | 28.0 (4.4) | 16.9 (3.8) | 11.3 (2.3) | 9.6(2.2) |
| Rmerge (%) | 12.2 (39.7) | 8.6 (31.0) | 12.4 (41.4) | 13.9 (38.2) | 16.5 (43.7) |
| Refinement | |||||
| Reflections used in refinement | 214406 (16993) | 387739 (37693) | 50455 (4819) | 109291 (10188) | 79574 (7627) |
| Reflections used for Rfree | 10759 (868) | 19516 (1870) | 2481 (270) | 5527 (483) | 3906 (344) |
| Rwork | 0.22 (0.35) | 0.20 (0.21) | 0.19 (0.25) | 0.24 (0.28) | 0.21 (0.27) |
| Rfree | 0.26 (0.36) | 0.23 (0.24) | 0.26 (0.35) | 0.29 (0.33) | 0.26 (0.32) |
| Number of non-hydrogen atoms | 23700 | 24548 | 11853 | 11315 | 11670 |
| macromolecules | 22738 | 23404 | 11540 | 10640 | 11349c |
| ligands | 8 | 8 | 4 | 8 | 4 |
| solvent | 954 | 1136 | 309 | 667 | 317 |
| Protein residues | 2887 | 2976 | 1460 | 1347 | 1444 |
| RMS (bonds) | 0.014 | 0.015 | 0.016 | 0.022 | 0.015 |
| RMS (angles) | 1.84 | 1.92 | 1.97 | 2.09 | 2.00 |
| Ramachandran favored (%) | 95.5 | 96.7 | 93.1 | 92.7 | 93.8 |
| Ramachandran allowed (%) | 4.1 | 2.7 | 6.0 | 5.6 | 5.2 |
| Ramachandran outliers (%) | 0.4 | 0.6 | 0.9 | 1.7 | 1.0 |
| Rotamer outliers (%) | 4.2 | 3.0 | 5.3 | 5.0 | 4.0 |
| Clash score | 7.3 | 4.3 | 6.7 | 9.0 | 5.9 |
| Average B-factor | 28.1 | 14.3 | 31.3 | 26.9 | 20.4 |
| macromolecules | 28.1 | 14.2 | 31.48 | 26.7 | 20.5 |
| ligands | 21.4 | 9.4 | 26.21 | 19.8 | 18.0 |
| solvent | 27.9 | 17.6 | 24.74 | 30.6 | 16.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.S.; Kim, T.Y.; Park, H.; Lee, J.H.; Nguyen, D.Q.; Hong, M.-K.; Lee, S.H.; Kang, L.-W. Structural Study of Metal Binding and Coordination in Ancient Metallo-?-Lactamase PNGM-1 Variants. Int. J. Mol. Sci. 2020, 21, 4926. https://doi.org/10.3390/ijms21144926
Park YS, Kim TY, Park H, Lee JH, Nguyen DQ, Hong M-K, Lee SH, Kang L-W. Structural Study of Metal Binding and Coordination in Ancient Metallo-?-Lactamase PNGM-1 Variants. International Journal of Molecular Sciences. 2020; 21(14):4926. https://doi.org/10.3390/ijms21144926
Chicago/Turabian StylePark, Yoon Sik, Tae Yeong Kim, Hyunjae Park, Jung Hun Lee, Diem Quynh Nguyen, Myoung-Ki Hong, Sang Hee Lee, and Lin-Woo Kang. 2020. "Structural Study of Metal Binding and Coordination in Ancient Metallo-?-Lactamase PNGM-1 Variants" International Journal of Molecular Sciences 21, no. 14: 4926. https://doi.org/10.3390/ijms21144926
APA StylePark, Y. S., Kim, T. Y., Park, H., Lee, J. H., Nguyen, D. Q., Hong, M.-K., Lee, S. H., & Kang, L.-W. (2020). Structural Study of Metal Binding and Coordination in Ancient Metallo-?-Lactamase PNGM-1 Variants. International Journal of Molecular Sciences, 21(14), 4926. https://doi.org/10.3390/ijms21144926