Quipazine Elicits Swallowing in the Arterially Perfused Rat Preparation: A Role for Medullary Raphe Nuclei?



Abstract
:1. Introduction
2. Results
2.1. Systemic QPZ Effects on Cardiorespiratory Parameters
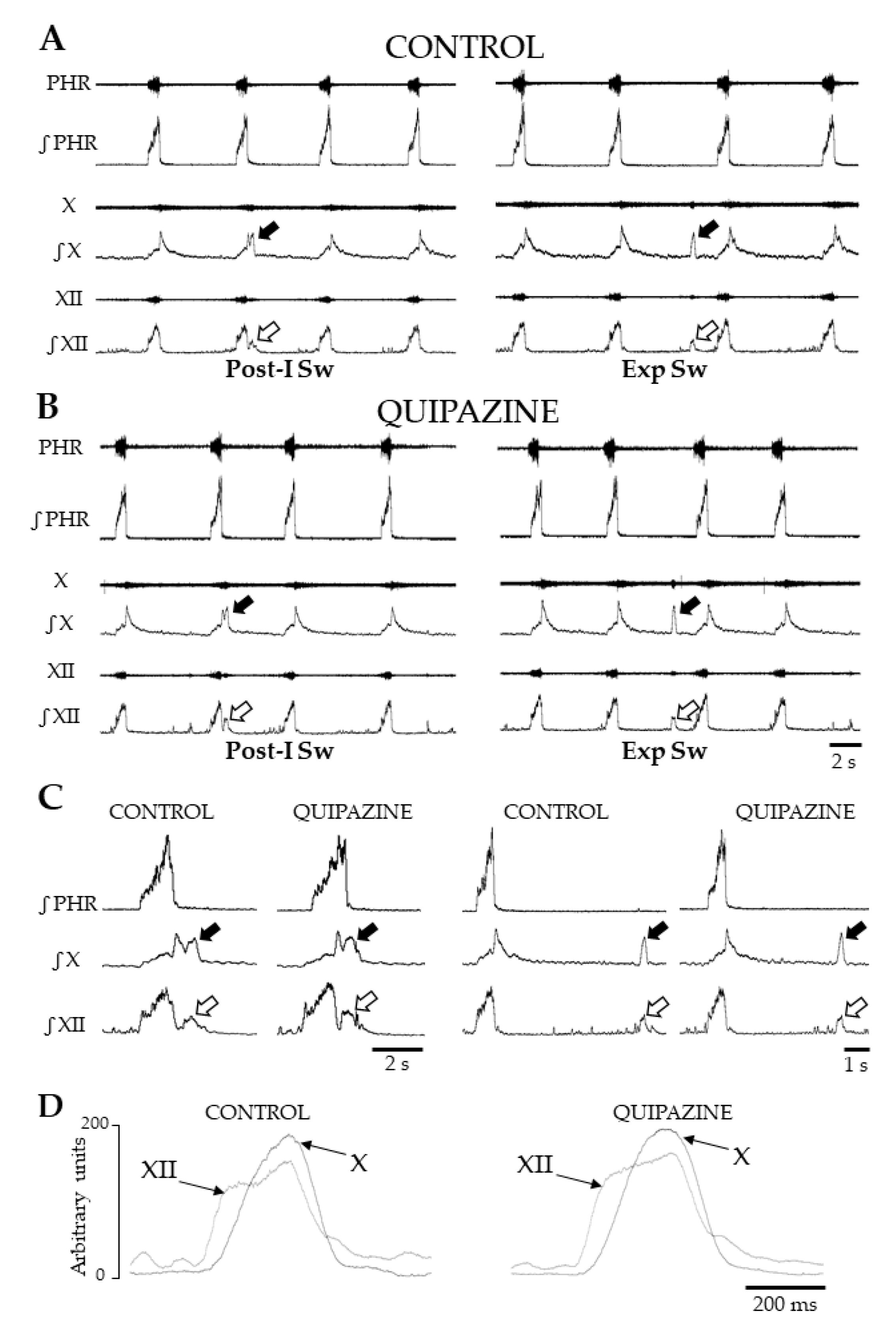
2.2. Characterization of Swallows
2.3. Systemic QPZ Increased Swallow Rate
2.4. Analyses of Swallow-Breathing Coordination and Effect of Swallowing on Central Respiration
2.5. Effects of Central Drug Injections on Swallowing
3. Discussion
3.1. Distribution of Serotonergic Receptors in Brainstem Respiratory and Swallowing Networks
3.2. Specificity and Functional Effects of Quipazine
3.3. Effects of QPZ on Swallowing
3.4. Swallow-Breathing Coordination In Situ
3.5. Is the NTS Involved in Serotonergic Modulation of Swallowing?
3.6. Medullary Raphe Nuclei May Modulate Swallowing
3.7. Functional Relevance of Findings
4. Materials and Methods
4.1. Working Heart-Brainstem or In Situ Preparation
4.2. Recording and Analyses of Cardiorespiratory and Swallowing Parameters
4.3. Pharmacological Induction of Swallowing
4.4. Experimental Protocol and Parameter Analyses
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Altman, K.W. Understanding dysphagia: A rapidly emerging problem. Otolaryngol. Clin. N. Am. 2013, 46, xiii–xvi. [Google Scholar] [CrossRef]
- Costa, M.M.B. Neural Control of Swallowing. Arq. Gastroenterol. 2018, 55 (Suppl. 1), 61–75. [Google Scholar] [CrossRef]
- Miller, N. Swallowing in Parkinson’s disease: Clinical issues and management. Neurodegener. Dis. Manag. 2017, 7, 205–217. [Google Scholar] [CrossRef]
- Suntrup, S.; Teismann, I.; Bejer, J.; Suttrup, I.; Winkels, M.; Mehler, D.; Pantev, C.; Dziewas, R.; Warnecke, T. Evidence for adaptive cortical changes in swallowing in Parkinson’s disease. Brain J. Neurol. 2013, 136, 726–738. [Google Scholar] [CrossRef]
- Bieger, D. Role of bulbar serotonergic neurotransmission in the initiation of swallowing in the rat. Neuropharmacology 1981, 20, 1073–1083. [Google Scholar] [CrossRef]
- Jean, A. Brain stem control of swallowing: Neuronal network and cellular mechanisms. Physiol. Rev. 2001, 81, 929–969. [Google Scholar] [CrossRef] [Green Version]
- Gestreau, C.; Dutschmann, M.; Obled, S.; Bianchi, A.L. Activation of XII motoneurons and premotor neurons during various oropharyngeal behaviors. Respir. Physiol. Neurobiol. 2005, 147, 159–176. [Google Scholar] [CrossRef]
- Horton, K.K.; Segers, L.S.; Nuding, S.C.; O’Connor, R.; Alencar, P.A.; Davenport, P.W.; Bolser, D.C.; Pitts, T.; Lindsey, B.G.; Morris, K.F.; et al. Central Respiration and Mechanical Ventilation in the Gating of Swallow with Breathing. Front. Physiol. 2018, 9, 785. [Google Scholar] [CrossRef] [PubMed]
- Dick, T.E.; Oku, Y.; Romaniuk, J.R.; Cherniack, N.S. Interaction between central pattern generators for breathing and swallowing in the cat. J. Physiol. 1993, 465, 715–730. [Google Scholar] [CrossRef] [Green Version]
- Gestreau, C.; Milano, S.; Bianchi, A.L.; Grelot, L. Activity of dorsal respiratory group inspiratory neurons during laryngeal-induced fictive coughing and swallowing in decerebrate cats. Exp. Brain Res. 1996, 108, 247–256. [Google Scholar] [CrossRef]
- Roda, F.; Pio, J.; Bianchi, A.L.; Gestreau, C. Effects of anesthetics on hypoglossal nerve discharge and c-Fos expression in brainstem hypoglossal premotor neurons. J. Comp. Neurol. 2004, 468, 571–586. [Google Scholar] [CrossRef] [PubMed]
- Bautista, T.G.; Dutschmann, M. Ponto-medullary nuclei involved in the generation of sequential pharyngeal swallowing and concomitant protective laryngeal adduction in situ. J. Physiol. 2014, 592, 2605–2623. [Google Scholar] [CrossRef] [PubMed]
- Fuse, S.; Sugiyama, Y.; Hashimoto, K.; Umezaki, T.; Oku, Y.; Dutschmann, M.; Hirano, S. Laryngeal afferent modulation of swallowing interneurons in the dorsal medulla in perfused rats. Laryngoscope 2019. [Google Scholar] [CrossRef]
- Hashimoto, K.; Sugiyama, Y.; Fuse, S.; Umezaki, T.; Oku, Y.; Dutschmann, M.; Hirano, S. Activity of swallowing-related neurons in the medulla in the perfused brainstem preparation in rats. Laryngoscope 2019, 129, E72–E79. [Google Scholar] [CrossRef]
- Paton, J.F.; Li, Y.W.; Kasparov, S. Reflex response and convergence of pharyngoesophageal and peripheral chemoreceptors in the nucleus of the solitary tract. Neuroscience 1999, 93, 143–154. [Google Scholar] [CrossRef]
- Bautista, T.G.; Fong, A.Y.; Dutschmann, M. Spontaneous swallowing occurs during autoresuscitation in the in situ brainstem preparation of rat. Respir. Physiol. Neurobiol. 2014, 202, 35–43. [Google Scholar] [CrossRef]
- Roda, F.; Gestreau, C.; Bianchi, A.L. Discharge patterns of hypoglossal motoneurons during fictive breathing, coughing, and swallowing. J. Neurophysiol. 2002, 87, 1703–1711. [Google Scholar] [CrossRef] [Green Version]
- Kessler, J.P.; Jean, A. Evidence that activation of N-methyl-D-aspartate (NMDA) and non-NMDA receptors within the nucleus tractus solitarii triggers swallowing. Eur. J. Pharm. 1991, 201, 59–67. [Google Scholar] [CrossRef]
- Hashim, M.A.; Bieger, D. Excitatory action of 5-HT on deglutitive substrates in the rat solitary complex. Brain Res. Bull. 1987, 18, 355–363. [Google Scholar] [CrossRef]
- Haney, M.M.; Sinnott, J.; Osman, K.L.; Deninger, I.; Andel, E.; Caywood, V.; Mok, A.; Ballenger, B.; Cummings, K.; Thombs, L.; et al. Mice Lacking Brain-Derived Serotonin Have Altered Swallowing Function. Otolaryngol. Head Neck Surg. 2019, 161, 468–471. [Google Scholar] [CrossRef] [PubMed]
- Kessler, J.P.; Jean, A. Inhibition of the swallowing reflex by local application of serotonergic agents into the nucleus of the solitary tract. Eur. J. Pharm. 1985, 118, 77–85. [Google Scholar] [CrossRef]
- Kessler, J.P.; Jean, A. Inhibitory influence of monoamines and brainstem monoaminergic regions on the medullary swallowing reflex. Neurosci. Lett. 1986, 65, 41–46. [Google Scholar] [CrossRef]
- St-John, W.M.; Paton, J.F. Characterizations of eupnea, apneusis and gasping in a perfused rat preparation. Respir. Physiol. 2000, 123, 201–213. [Google Scholar] [CrossRef]
- Benarroch, E.E. Medullary serotonergic system: Organization, effects, and clinical correlations. Neurology 2014, 83, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Do, H.J.; Seo, H.G.; Lee, H.H.; Oh, B.M.; Kim, Y.; Kim, A.; Kim, H.J.; Jeon, B.; Han, T.R. tProgression of Oropharyngeal Dysphagia in Patients with Multiple System Arophy. Dysphagia 2020, 35, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Simons, J.A. Swallowing Dysfunctions in Parkinson’s Disease. Int. Rev. Neurobiol. 2017, 134, 1207–1238. [Google Scholar] [CrossRef]
- Saito, Y.; Ezure, K.; Tanaka, I. Swallowing-related activities of respiratory and non-respiratory neurons in the nucleus of solitary tract in the rat. J. Physiol. 2002, 540, 1047–1060. [Google Scholar] [CrossRef]
- Paterson, D.S.; Darnall, R. 5-HT2A receptors are concentrated in regions of the human infant medulla involved in respiratory and autonomic control. Auton. Neurosci. Basic Clin. 2009, 147, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Descarries, L.; Riad, M.; Parent, M. Ultrastructure of the serotonin innervation in the mammalian central nervous system. Handb. Behav. Neurobiol. Serotonin 2010, 21, 65–101. [Google Scholar]
- Holtman, J.R., Jr. Immunohistochemical localization of serotonin- and substance P-containing fibers around respiratory muscle motoneurons in the nucleus ambiguus of the cat. Neuroscience 1988, 26, 169–178. [Google Scholar] [CrossRef]
- Holtman, J.R., Jr.; Vascik, D.S.; Maley, B.E. Ultrastructural evidence for serotonin-immunoreactive terminals contacting phrenic motoneurons in the cat. Exp. Neurol. 1990, 109, 269–272. [Google Scholar] [CrossRef]
- Hodges, M.R.; Richerson, G.B. Contributions of 5-HT neurons to respiratory control: Neuromodulatory and trophic effects. Respir. Physiol. Neurobiol. 2008, 164, 222–232. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, B.L.; Azmitia, E.C. Structure and function of the brain serotonin system. Physiol. Rev. 1992, 72, 165–229. [Google Scholar] [CrossRef] [Green Version]
- Voss, M.D.; De Castro, D.; Lipski, J.; Pilowsky, P.M.; Jiang, C. Serotonin immunoreactive boutons form close appositions with respiratory neurons of the dorsal respiratory group in the cat. J. Comp. Neurol. 1990, 295, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Okabe, S.; Mackiewicz, M.; Kubin, L. Serotonin receptor mRNA expression in the hypoglossal motor nucleus. Respir. Physiol. 1997, 110, 151–160. [Google Scholar] [CrossRef]
- Austgen, J.R.; Dantzler, H.A.; Barger, B.K.; Kline, D.D. 5-hydroxytryptamine 2C receptors tonically augment synaptic currents in the nucleus tractus solitarii. J. Neurophysiol. 2012, 108, 2292–2305. [Google Scholar] [CrossRef] [Green Version]
- Nosjean, A.; Callera, J.C.; Bonagamba, L.; Machado, B.; Hamon, M.; Laguzzi, R. Serotonin (3) receptor stimulation in the nucleus tractus solitarii activates non-catecholaminergic neurons in the rat ventrolateral medulla. Neuroscience 2002, 112, 935–949. [Google Scholar] [CrossRef]
- Edwards, E.; Paton, J.F. 5-HT (4) receptors in nucleus tractus solitarii attenuate cardiopulmonary reflex in anesthetized rats. Am. J. Physiol. 1999, 277, H1914–H1923. [Google Scholar] [CrossRef]
- Matott, M.P.; Kline, D.D. Activation of 5-hyrdoxytryptamine 7 receptors within the rat nucleus tractus solitarii modulates synaptic properties. Brain Res. 2016, 1635, 12–26. [Google Scholar] [CrossRef] [Green Version]
- Cornea-Hebert, V.; Riad, M.; Wu, C.; Singh, S.K.; Descarries, L. Cellular and subcellular distribution of the serotonin 5-HT2A receptor in the central nervous system of adult rat. J. Comp. Neurol. 1999, 409, 187–209. [Google Scholar] [CrossRef]
- Doi, A.; Ramirez, J.M. State-dependent interactions between excitatory neuromodulators in the neuronal control of breathing. J. Neurosci. 2010, 30, 8251–8262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzke, T.; Niebert, M.; Koch, U.R.; Caley, A.; Vogelgesang, S.; Hulsmann, S.; Ponimaskin, E.; Muller, U.; Smart, T.G.; Harvey, R.J.; et al. Serotonin receptor 1A-modulated phosphorylation of glycine receptor alpha3 controls breathing in mice. J. Clin. Investig. 2010, 120, 4118–4128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niebert, M.; Vogelgesang, S.; Koch, U.R.; Bischoff, A.M.; Kron, M.; Bock, N.; Manzke, T. Expression and function of serotonin 2A and 2B receptors in the mammalian respiratory network. PLoS ONE 2011, 6, e21395. [Google Scholar] [CrossRef]
- Gobbi, G.; Cassano, T.; Radja, F.; Morgese, M.G.; Cuomo, V.; Santarelli, L.; Hen, R.; Blier, P. Neurokinin 1 receptor antagonism requires norepinephrine to increase serotonin function. Eur. Neuropsychopharmacol. 2007, 17, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wong-Riley, M.T. Postnatal changes in the expressions of serotonin 1A, 1B, and 2A receptors in ten brain stem nuclei of the rat: Implication for a sensitive period. Neuroscience 2010, 165, 61–78. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ramage, A.G. The role of central 5-HT (1A) receptors in the control of B-fibre cardiac and bronchoconstrictor vagal preganglionic neurones in anaesthetized cats. J. Physiol. 2001, 536, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Dergacheva, O.; Griffioen, K.J.; Wang, X.; Kamendi, H.; Gorini, C.; Mendelowitz, D. 5-HT (2) receptor subtypes mediate different long-term changes in GABAergic activity to parasympathetic cardiac vagal neurons in the nucleus ambiguus. Neuroscience 2007, 149, 696–705. [Google Scholar] [CrossRef] [Green Version]
- Mantilla, C.B.; Bailey, J.P.; Zhan, W.Z.; Sieck, G.C. Phrenic motoneuron expression of serotonergic and glutamatergic receptors following upper cervical spinal cord injury. Exp. Neurol. 2012, 234, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.L.; Fuller, R.W. Antagonism of vascular serotonin receptors by m-chlorophenylpiperazine and m-trifluoromethylphenylpiperazine. Life Sci. 1983, 32, 711–718. [Google Scholar] [CrossRef]
- Glennon, R.A.; Young, R.; Rosecrans, J.A. Antagonism of the effects of the hallucinogen DOM and the purported 5-HT agonist quipazine by 5-HT2 antagonists. Eur. J. Pharm. 1983, 91, 189–196. [Google Scholar] [CrossRef]
- Gorea, E.; Adrien, J. Serotonergic regulation of noradrenergic coerulean neurons: Electrophysiological evidence for the involvement of 5-HT2 receptors. Eur. J. Pharm. 1988, 154, 285–291. [Google Scholar] [CrossRef]
- Goodwin, G.M.; Green, A.R. A behavioural and biochemical study in mice and rats of putative selective agonists and antagonists for 5-HT1 and 5-HT2 receptors. Br. J. Pharmacol. 1985, 84, 743–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peroutka, S.J. Pharmacological differentiation and characterization of 5-HT1A, 5-HT1B, and 5-HT1C binding sites in rat frontal cortex. J. Neurochem. 1986, 47, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Sills, M.A.; Wolfe, B.B.; Frazer, A. Determination of selective and nonselective compounds for the 5-HT 1A and 5-HT 1B receptor subtypes in rat frontal cortex. J. Pharmacol. Exp. Ther. 1984, 231, 480–487. [Google Scholar] [PubMed]
- Ireland, S.J.; Tyers, M.B. Pharmacological characterization of 5-hydroxytryptamine-induced depolarization of the rat isolated vagus nerve. Br. J. Pharmacol. 1987, 90, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Round, A.; Wallis, D.I. Further studies on the blockade of 5-HT depolarizations of rabbit vagal afferent and sympathetic ganglion cells by MDL 72222 and other antagonists. Neuropharmacology 1987, 26, 39–48. [Google Scholar] [CrossRef]
- Courtine, G.; Gerasimenko, Y.; van den Brand, R.; Yew, A.; Musienko, P.; Zhong, H.; Song, B.; Ao, Y.; Ichiyama, R.M.; Lavrov, I.; et al. Transformation of nonfunctional spinal circuits into functional states after the loss of brain input. Nat. Neurosci. 2009, 12, 1333–1342. [Google Scholar] [CrossRef]
- Swann, H.E.; Kauer, S.D.; Allmond, J.T.; Brumley, M.R. Stimulation of 5-HT2A receptors recovers sensory responsiveness in acute spinal neonatal rats. Behav. Neurosci. 2017, 131, 92–98. [Google Scholar] [CrossRef]
- Oueghlani, Z.; Juvin, L.; Lambert, F.M.; Cardoit, L.; Courtand, G.; Masmejean, F.; Cazalets, J.R.; Barriere, G. Serotonergic modulation of sacral dorsal root stimulation-induced locomotor output in newborn rat. Neuropharmacology 2019. [Google Scholar] [CrossRef]
- Chopek, J.W.; MacDonell, C.W.; Power, K.E.; Gardiner, K.; Gardiner, P.F. Removal of supraspinal input reveals a difference in the flexor and extensor monosynaptic reflex response to quipazine independent of motoneuron excitation. J. Neurophysiol. 2013, 109, 2056–2063. [Google Scholar] [CrossRef]
- Slawinska, U.; Miazga, K.; Jordan, L.M. 5-HT (2) and 5-HT (7) receptor agonists facilitate plantar stepping in chronic spinal rats through actions on different populations of spinal neurons. Front. Neural Circuits 2014, 8, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker-Herman, T.L.; Fuller, D.D.; Bavis, R.W.; Zabka, A.G.; Golder, F.J.; Doperalski, N.J.; Johnson, R.A.; Watters, J.J.; Mitchell, G.S. BDNF is necessary and sufficient for spinal respiratory plasticity following intermittent hypoxia. Nat. Neurosci. 2004, 7, 48–55. [Google Scholar] [CrossRef]
- Bou-Flores, C.; Lajard, A.M.; Monteau, R.; De Maeyer, E.; Seif, I.; Lanoir, J.; Hilaire, G. Abnormal phrenic motoneuron activity and morphology in neonatal monoamine oxidase A-deficient transgenic mice: Possible role of a serotonin excess. J. Neurosci. 2000, 20, 4646–4656. [Google Scholar] [CrossRef] [Green Version]
- Cayetanot, F.; Gros, F.; Larnicol, N. Postnatal changes in the respiratory response of the conscious rat to serotonin 2A/2C receptor activation are reflected in the developmental pattern of fos expression in the brainstem. Brain Res. 2002, 942, 51–57. [Google Scholar] [CrossRef]
- Kinkead, R.; Belzile, O.; Gulemetova, R. Serotonergic modulation of respiratory motor output during tadpole development. J. Appl. Physiol. 2002, 93, 936–946. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.M.; Tryba, A.K.; Pena, F. Pacemaker neurons and neuronal networks: An integrative view. Curr. Opin. Neurobiol. 2004, 14, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.M.; Wang, J.; Ni, C.R.; Yu, J.G.; Wang, W.Z.; Su, D.F. Ketanserin-induced baroreflex enhancement in spontaneously hypertensive rats depends on central 5-HT(2A) receptors. Clin. Exp. Pharmacol. Physiol. 2007, 34, 702–707. [Google Scholar] [CrossRef]
- Villalon, C.M.; Centurion, D. Cardiovascular responses produced by 5-hydroxytriptamine: A pharmacological update on the receptors/mechanisms involved and therapeutic implications. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2007, 376, 45–63. [Google Scholar] [CrossRef]
- Zhan, G.; Shaheen, F.; Mackiewicz, M.; Fenik, P.; Veasey, S.C. Single cell laser dissection with molecular beacon polymerase chain reaction identifies 2A as the predominant serotonin receptor subtype in hypoglossal motoneurons. Neuroscience 2002, 113, 145–154. [Google Scholar] [CrossRef]
- Bobker, D.H. A slow excitatory postsynaptic potential mediated by 5-HT2 receptors in nucleus prepositus hypoglossi. J. Neurosci. 1994, 14, 2428–2434. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, A.D.; Feldman, J.L. Modulation of respiratory activity of neonatal rat phrenic motoneurones by serotonin. J. Physiol. 1993, 461, 213–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, D.; Monteau, R.; Hilaire, G. Compared effects of serotonin on cervical and hypoglossal inspiratory activities: An in vitro study in the newborn rat. J. Physiol. 1992, 451, 605–629. [Google Scholar] [CrossRef] [Green Version]
- Lalley, P.M.; Bischoff, A.M.; Schwarzacher, S.W.; Richter, D.W. 5-HT2 receptor-controlled modulation of medullary respiratory neurones in the cat. J. Physiol. 1995, 487 Pt 3, 653–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickering, A.E.; Spanswick, D.; Logan, S.D. 5-Hydoxytryptamine evokes depolarizations and membrane potential oscillations in rat sympathetic preganglionic neurones. J. Physiol. 1994, 480 Pt 1, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Ogasa, T.; Ray, A.D.; Michlin, C.P.; Farkas, G.A.; Grant, B.J.; Magalang, U.J. Systemic administration of serotonin 2A/2C agonist improves upper airway stability in Zucker rats. Am. J. Respir. Crit. Care Med. 2004, 170, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Bieger, D. Rhombencephalic pathways and neurotransmitters controlling deglutition. Am. J. Med. 2001, 111 (Suppl. 8A), 85–89. [Google Scholar] [CrossRef]
- Ertekin, C.; Aydogdu, I. Neurophysiology of swallowing. Clin. Neurophysiol. 2003, 114, 2226–2244. [Google Scholar] [CrossRef]
- Jean, A. Brainstem organization of the swallowing network. Brain Behav. Evol. 1984, 25, 109–116. [Google Scholar] [CrossRef]
- Poliacek, I.; Jakus, J.; Simera, M.; Veternik, M.; Plevkova, J. Control of coughing by medullary raphe. Prog. Brain Res. 2014, 212, 277–295. [Google Scholar] [CrossRef]
- Sotelo, C.; Cholley, B.; El Mestikawy, S.; Gozlan, H.; Hamon, M. Direct Immunohistochemical Evidence of the Existence of 5-HT1A Autoreceptors on Serotoninergic Neurons in the Midbrain Raphe Nuclei. Eur. J. Neurosci. 1990, 2, 1144–1154. [Google Scholar] [CrossRef]
- Bayliss, D.A.; Li, Y.W.; Talley, E.M. Effects of serotonin on caudal raphe neurons: Inhibition of N- and P/Q-type calcium channels and the afterhyperpolarization. J. Neurophysiol. 1997, 77, 1362–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayliss, D.A.; Li, Y.W.; Talley, E.M. Effects of serotonin on caudal raphe neurons: Activation of an inwardly rectifying potassium conductance. J. Neurophysiol. 1997, 77, 1349–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, J.; Bland, B.H.; Antle, M.C. Nonserotonergic projection neurons in the midbrain raphe nuclei contain the vesicular glutamate transporter VGLUT3. Synapse 2009, 63, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Fountain, M.D.; Schaaf, C.P. Prader-Willi Syndrome and Schaaf-Yang Syndrome: Neurodevelopmental Diseases Intersecting at the MAGEL2 Gene. Diseases 2016, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanella, S.; Barthelemy, M.; Muscatelli, F.; Hilaire, G. Necdin gene, respiratory disturbances and Prader-Willi syndrome. Adv. Exp. Med. Biol. 2008, 605, 159–164. [Google Scholar] [CrossRef]
- Zanella, S.; Tauber, M.; Muscatelli, F. Breathing deficits of the Prader-Willi syndrome. Respir. Physiol. Neurobiol. 2009, 168, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Matarazzo, V.; Caccialupi, L.; Schaller, F.; Shvarev, Y.; Kourdougli, N.; Bertoni, A.; Menuet, C.; Voituron, N.; Deneris, E.; Gaspar, P.; et al. Necdin shapes serotonergic development and SERT activity modulating breathing in a mouse model for Prader-Willi syndrome. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Paton, J.F. A working heart-brainstem preparation of the mouse. J. Neurosci. Methods 1996, 65, 63–68. [Google Scholar] [CrossRef]
- Wang, S.; Benamer, N.; Zanella, S.; Kumar, N.N.; Shi, Y.; Bevengut, M.; Penton, D.; Guyenet, P.G.; Lesage, F.; Gestreau, C.; et al. TASK-2 channels contribute to pH sensitivity of retrotrapezoid nucleus chemoreceptor neurons. J. Neurosci. 2013, 33, 16033–16044. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hypoglossal Nerve | Vagus Nerve | ||||
|---|---|---|---|---|---|
| Control | QPZ | Control | QPZ | ||
| Normalized Amplitude (%) | 100 | 101 ± 19 | 100 | 97 ± 9 | ns |
| Normalized area under the curve (%) | 100 | 107 ± 19 | 100 | 98 ± 13 | ns |
| Duration (s) | 0.56 ± 0.12 | 0.51 ± 0.14 | 0.43 ± 0.08 | 0.43 ± 0.06 | ns |
| Obex (mm) | |||||
|---|---|---|---|---|---|
| Depth (mm) | +1 | +1.5 | +2 | +2.5 | +3 |
| 1.25 | (6) | 3 (11) | (11) | (1) | (2) |
| 1.75 | (7) | 9 (5) | 5 (9) | (1) | (2) |
| 2.25 | (7) | 8 (6) | 6 (6) | (1) | (1) |
| 2.75 | --- | --- | 1 (8) | (1) | (1) |
| Successful Sites (mm) | Swallow Rate (sw/min) | ||||
|---|---|---|---|---|---|
| From Obex | Depth | Control | QPZ 0–1 min | QPZ 0–5 min | Latency |
| +1.5 | −1.75 | 0.8 | 2.9 | 2.0 | 7.8 |
| +1.5 | −2.25 | 0.8 | 3.0 | 2.1 | 6.0 |
| +2 | −2.25 | 1.4 | 2.3 | 2.0 | 13.4 |
| Mean ± SD | 1.0 ± 0.3 | 2.7 ± 0.4 | 2.2 ± 0.1 | 9.1 ± 3.9 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergé-Laval, V.; Gestreau, C. Quipazine Elicits Swallowing in the Arterially Perfused Rat Preparation: A Role for Medullary Raphe Nuclei? Int. J. Mol. Sci. 2020, 21, 5120. https://doi.org/10.3390/ijms21145120
Bergé-Laval V, Gestreau C. Quipazine Elicits Swallowing in the Arterially Perfused Rat Preparation: A Role for Medullary Raphe Nuclei? International Journal of Molecular Sciences. 2020; 21(14):5120. https://doi.org/10.3390/ijms21145120
Chicago/Turabian StyleBergé-Laval, Victor, and Christian Gestreau. 2020. "Quipazine Elicits Swallowing in the Arterially Perfused Rat Preparation: A Role for Medullary Raphe Nuclei?" International Journal of Molecular Sciences 21, no. 14: 5120. https://doi.org/10.3390/ijms21145120
APA StyleBergé-Laval, V., & Gestreau, C. (2020). Quipazine Elicits Swallowing in the Arterially Perfused Rat Preparation: A Role for Medullary Raphe Nuclei? International Journal of Molecular Sciences, 21(14), 5120. https://doi.org/10.3390/ijms21145120