Overexpression of ATG5 Gene Makes Granulocyte-Like HL-60 Susceptible to Release Reactive Oxygen Species

 ,
,
Abstract
:1. Introduction
2. Results
2.1. ATG5 Overexpression in HL-60 Cells Does Not Result in the Upregulation of Autophagy
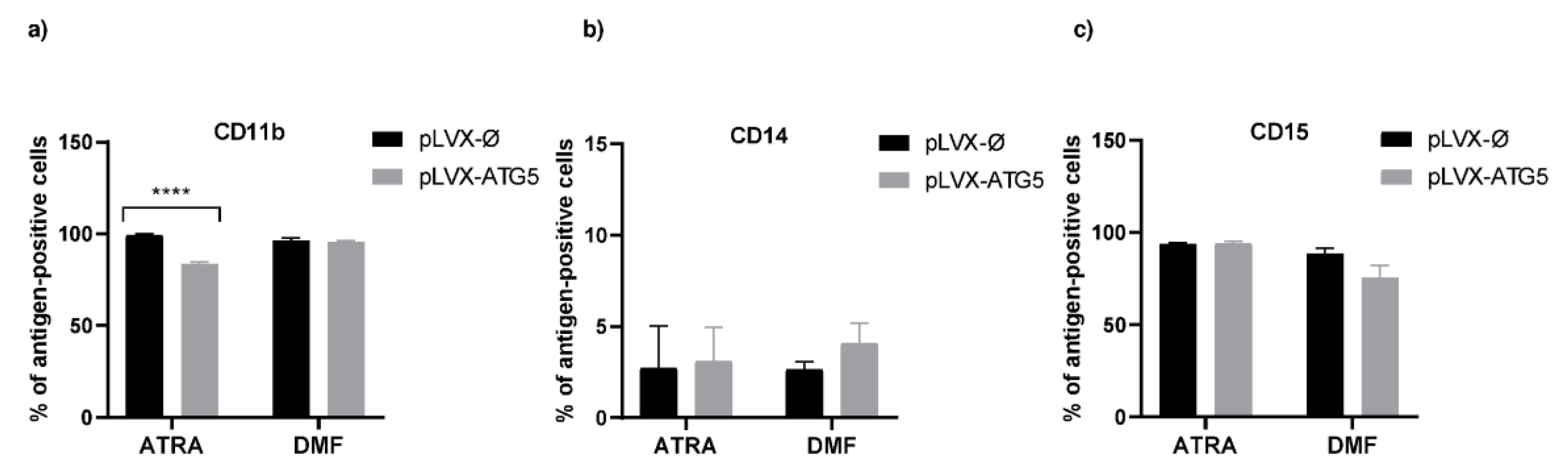
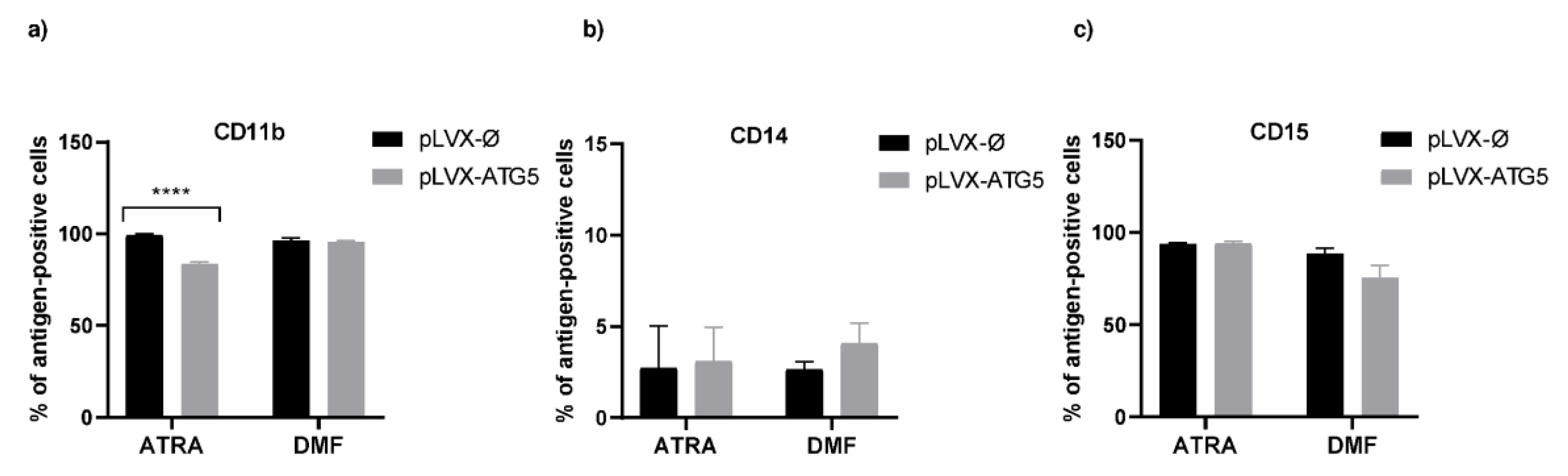
2.2. Increased Expression of ATG5 Affects Differentiation of HL-60 Cells with ATRA
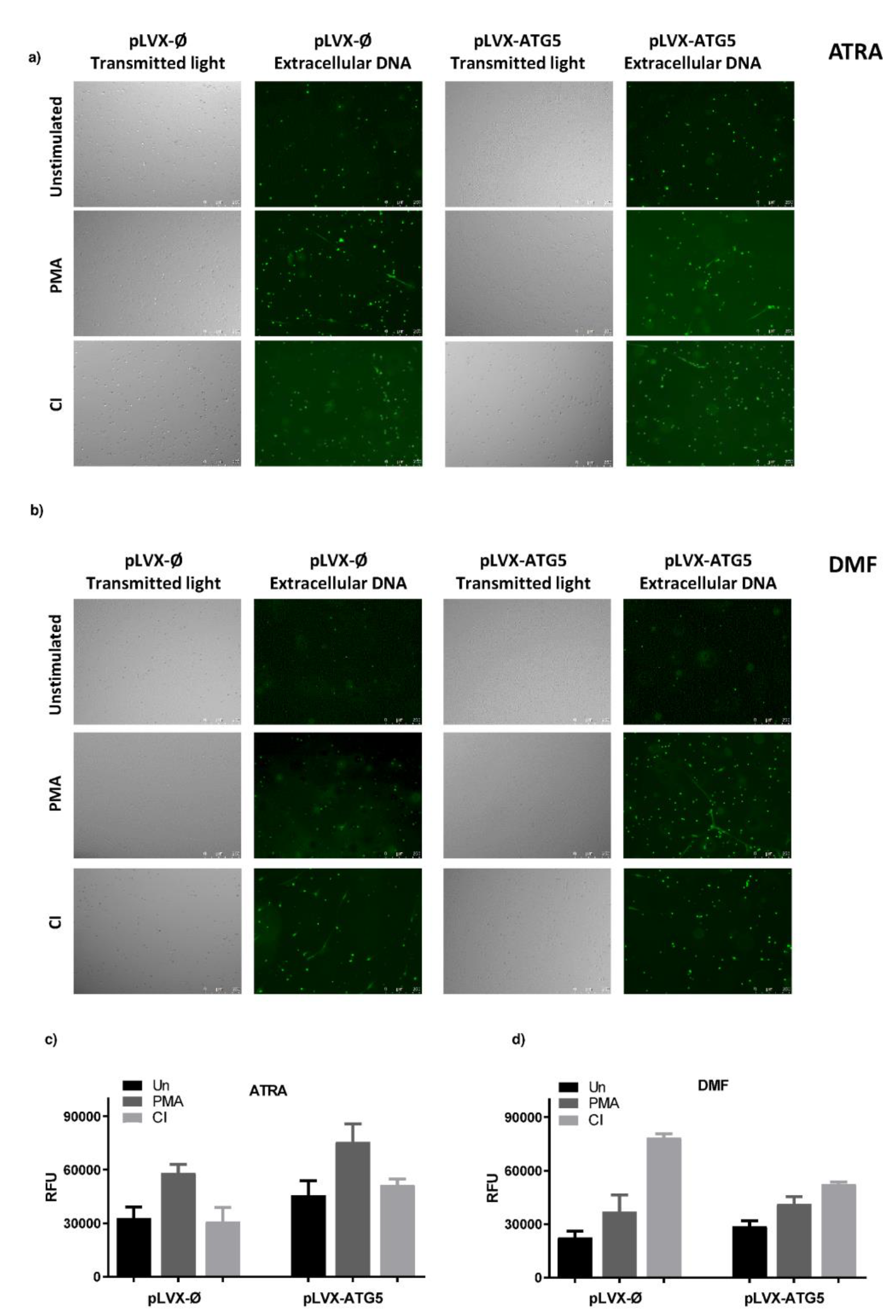
2.3. Increased Expression of ATG5 in Granulocyte-Like Cells Does Not Affect Ability to Form NETs
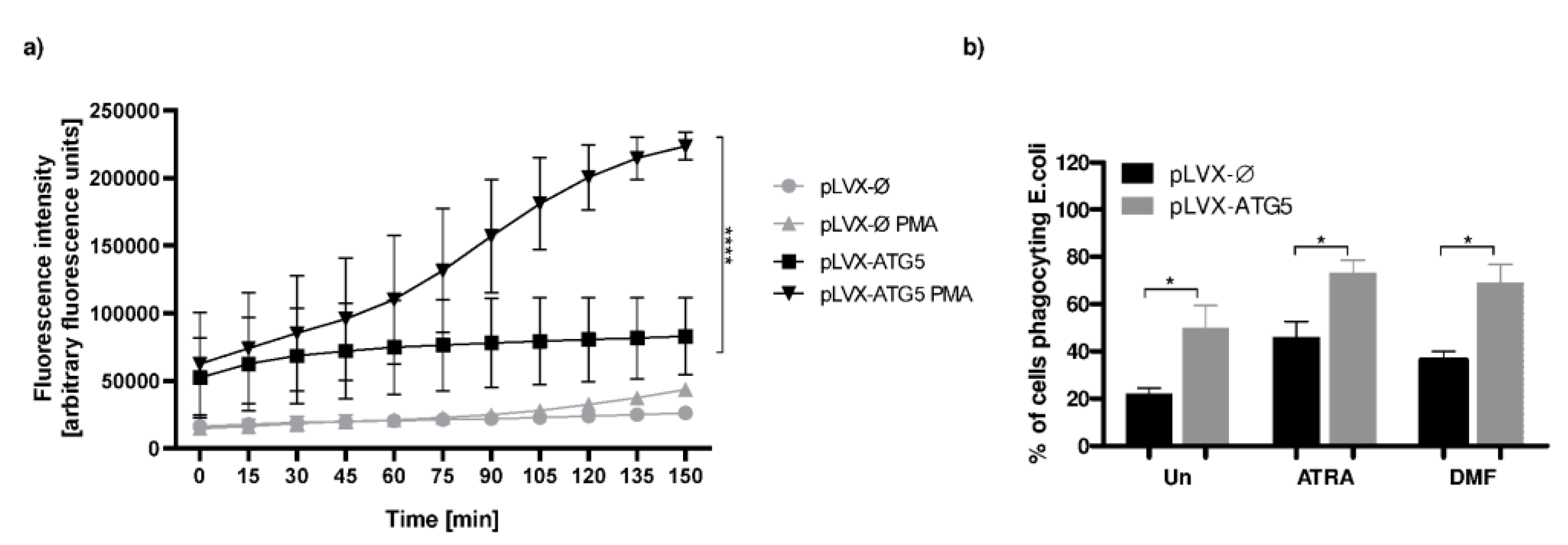
2.4. ROS Production Is Augmented in ATRA-dHL-60-ATG5 Cells
2.5. Phagocytosis Level Tends to Be Elevated in dHL-60 Cells Overexpressing ATG5
2.6. Increased Expression of ATG5 Does Not Influence the Proliferative Potential of HL-60 Cells
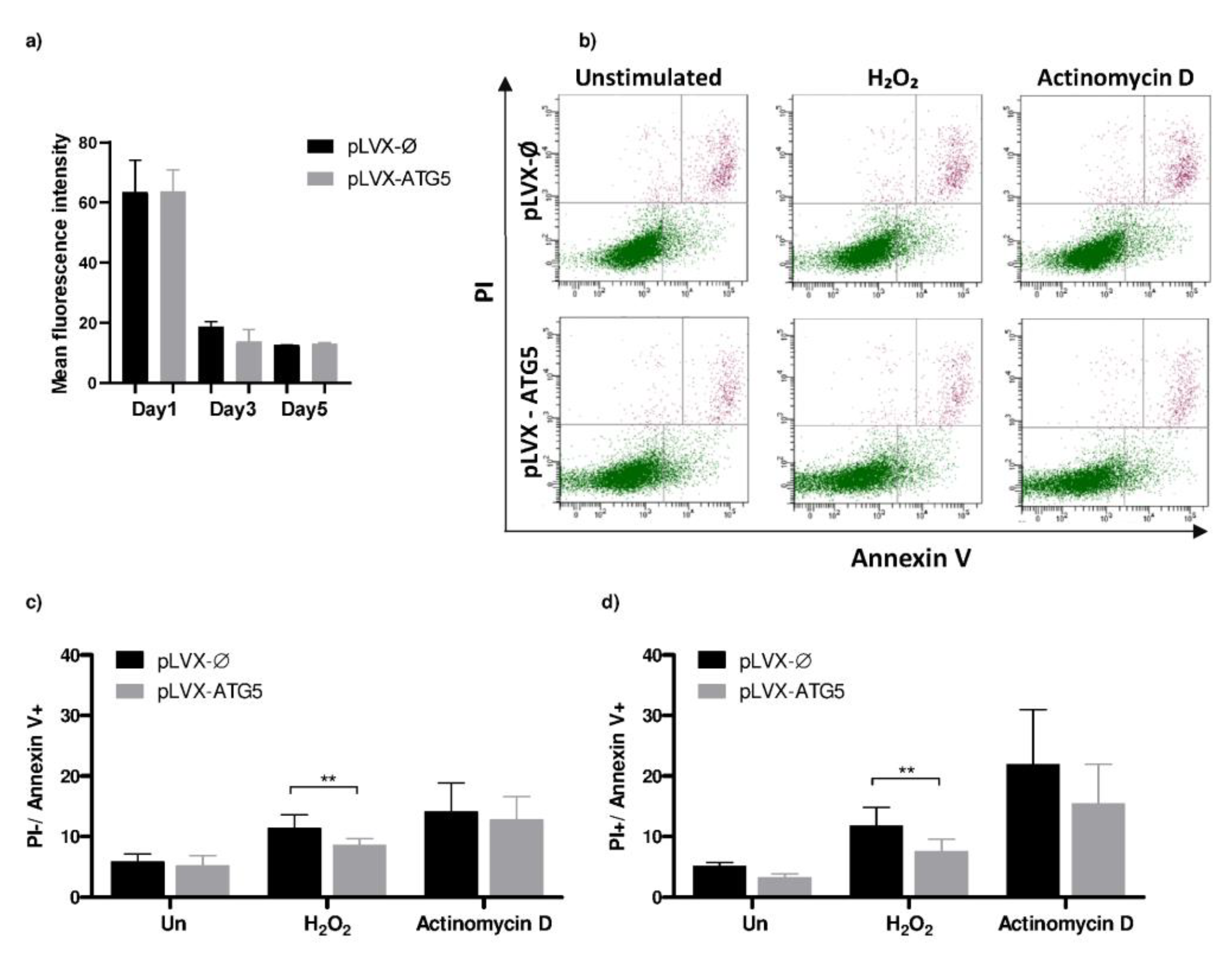
2.7. HL-60-ATG5 Are More Resistant to Chemical Apoptosis Inducers
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture and Differentiation
4.3. Plasmid Construction
4.4. Generation of HL-60 Cells Line Stably Overexpressing ATG5 Gene
4.5. qPCR
4.6. Western Blot
4.7. NETs Quantification
4.8. NETs Visualization
4.9. Oxidative Burst Measurement
4.10. Phagocytosis Assay
4.11. Proliferation Assay
4.12. Phosphatidylserine Externalization
4.13. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Kumar, V.; Sharma, A. Neutrophils: Cinderella of innate immune system. Int. Immunopharmacol. 2010, 10, 1325–1334. [Google Scholar] [CrossRef]
- Skendros, P.; Mitroulis, I.; Ritis, K. Autophagy in Neutrophils: From Granulopoiesis to Neutrophil Extracellular Traps. Front. Cell Dev. Biol. 2018, 6, 109. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Sun, B. Autophagy-mediated regulation of neutrophils and clinical applications. Burns Trauma 2020, 8, tkz001. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Fux, B.; Goodwin, M.; Dunay, I.R.; Strong, D.; Miller, B.C.; Cadwell, K.; Delgado, M.A.; Ponpuak, M.; Green, K.G.; et al. Autophagosome-independent essential function for the autophagy protein Atg5 in cellular immunity to intracellular pathogens. Cell Host Microbe 2008, 4, 458–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimmey, J.M.; Huynh, J.P.; Weiss, L.A.; Park, S.; Kambal, A.; Debnath, J.; Virgin, H.W.; Stallings, C.L. Unique role for ATG5 in neutrophil-mediated immunopathology during M. tuberculosis infection. Nature 2015, 528, 565–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remijsen, Q.; Vanden Berghe, T.; Wirawan, E.; Asselbergh, B.; Parthoens, E.; De Rycke, R.; Noppen, S.; Delforge, M.; Willems, J.; Vandenabeele, P. Neutrophil extracellular trap cell death requires both autophagy and superoxide generation. Cell Res. 2011, 21, 290–304. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, M.; Waguri, S.; Ueno, T.; Iwata, J.; Murata, S.; Tanida, I.; Ezaki, J.; Mizushima, N.; Ohsumi, Y.; Uchiyama, Y.; et al. Impairment of starvation-induced and constitutive autophagy in Atg7-deficient mice. J. Cell Biol. 2005, 169, 425–434. [Google Scholar] [CrossRef]
- Pierdominici, M.; Vomero, M.; Barbati, C.; Colasanti, T.; Maselli, A.; Vacirca, D.; Giovannetti, A.; Malorni, W.; Ortona, E. Role of autophagy in immunity and autoimmunity, with a special focus on systemic lupus erythematosus. FASEB J. 2012, 26, 1400–1412. [Google Scholar] [CrossRef]
- Galluzzi, L.; Green, D.R. Autophagy-Independent Functions of the Autophagy Machinery. Cell 2019, 177, 1682–1699. [Google Scholar] [CrossRef]
- Pua, H.H.; Dzhagalov, I.; Chuck, M.; Mizushima, N.; He, Y.W. A critical role for the autophagy gene Atg5 in T cell survival and proliferation. J. Exp. Med. 2007, 204, 25–31. [Google Scholar] [CrossRef]
- Ye, X.; Zhou, X.J.; Zhang, H. Exploring the Role of Autophagy-Related Gene 5 (ATG5) Yields Important Insights Into Autophagy in Autoimmune/Autoinflammatory Diseases. Front. Immunol. 2018, 9, 2334. [Google Scholar] [CrossRef] [PubMed]
- Manda-Handzlik, A.; Bystrzycka, W.; Wachowska, M.; Sieczkowska, S.; Stelmaszczyk-Emmel, A.; Demkow, U.; Ciepiela, O. The influence of agents differentiating HL-60 cells toward granulocyte-like cells on their ability to release neutrophil extracellular traps. Immunol. Cell Biol. 2018, 96, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Sham, R.L.; Phatak, P.D.; Belanger, K.A.; Packman, C.H. Functional properties of HL60 cells matured with all-trans-retinoic acid and DMSO: Differences in response to interleukin-8 and fMLP. Leuk. Res. 1995, 19, 1–6. [Google Scholar] [CrossRef]
- Weng, Y.M.; Ke, C.R.; Kong, J.Z.; Chen, H.; Hong, J.J.; Zhou, D.S. The significant role of ATG5 in the maintenance of normal functions of Mc3T3-E1 osteoblast. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Bardoel, B.W.; Kenny, E.F.; Sollberger, G.; Zychlinsky, A. The balancing act of neutrophils. Cell Host Microbe 2014, 15, 526–536. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Debnath, J. Autophagy and tumorigenesis. FEBS Lett. 2010, 584, 1427–1435. [Google Scholar] [CrossRef] [Green Version]
- Simon, H.U.; Yousefi, S.; Schmid, I.; Friis, R. ATG5 can regulate p53 expression and activation. Cell Death Dis. 2014, 5, e1339. [Google Scholar] [CrossRef] [Green Version]
- Maskey, D.; Yousefi, S.; Schmid, I.; Zlobec, I.; Perren, A.; Friis, R.; Simon, H.U. ATG5 is induced by DNA-damaging agents and promotes mitotic catastrophe independent of autophagy. Nat. Commun. 2013, 4, 2130. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Xie, W.; Yin, D.; Luo, R.; Liu, M.; Guo, F. ATG5 and ATG7 induced autophagy interplays with UPR via PERK signaling. Cell Commun. Signal. 2019, 17, 42. [Google Scholar] [CrossRef] [Green Version]
- Remijsen, Q.; Kuijpers, T.W.; Wirawan, E.; Lippens, S.; Vandenabeele, P.; Vanden Berghe, T. Dying for a cause: NETosis, mechanisms behind an antimicrobial cell death modality. Cell Death Differ. 2011, 18, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Orfali, N.; O’Donovan, T.R.; Nyhan, M.J.; Britschgi, A.; Tschan, M.P.; Cahill, M.R.; Mongan, N.P.; Gudas, L.J.; McKenna, S.L. Induction of autophagy is a key component of all-trans-retinoic acid-induced differentiation in leukemia cells and a potential target for pharmacologic modulation. Exp. Hematol. 2015, 43, 781–793.e2. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Cao, L.; Kang, R.; Yang, M.; Liu, L.; Zhao, Y.; Yu, Y.; Xie, M.; Yin, X.; Livesey, K.M.; et al. Autophagy regulates myeloid cell differentiation by p62/SQSTM1-mediated degradation of PML-RARalpha oncoprotein. Autophagy 2011, 7, 401–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britschgi, A.; Yousefi, S.; Laedrach, J.; Shan, D.; Simon, H.-U.; Tobler, A.; Fey, M.F.; Tschan, M.P. Blocking the Autophagy Gene 5 (ATG5) Impairs ATRA-Induced Myeloid Differentiation, and ATG5 Is Downregulated in AML. Blood 2008, 112, 309. [Google Scholar] [CrossRef]
- Xu, F.; Zhang, C.; Zou, Z.; Fan, E.K.Y.; Chen, L.; Li, Y.; Billiar, T.R.; Wilson, M.A.; Shi, X.; Fan, J. Aging-related Atg5 defect impairs neutrophil extracellular traps formation. Immunology 2017, 151, 417–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germic, N.; Stojkov, D.; Oberson, K.; Yousefi, S.; Simon, H.U. Neither eosinophils nor neutrophils require ATG5-dependent autophagy for extracellular DNA trap formation. Immunology 2017, 152, 517–525. [Google Scholar] [CrossRef]
- Tal, M.C.; Sasai, M.; Lee, H.K.; Yordy, B.; Shadel, G.S.; Iwasaki, A. Absence of autophagy results in reactive oxygen species-dependent amplification of RLR signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 2770–2775. [Google Scholar] [CrossRef] [Green Version]
- Pyo, J.O.; Yoo, S.M.; Ahn, H.H.; Nah, J.; Hong, S.H.; Kam, T.I.; Jung, S.; Jung, Y.K. Overexpression of Atg5 in mice activates autophagy and extends lifespan. Nat. Commun. 2013, 4, 2300. [Google Scholar] [CrossRef] [Green Version]
- Ullah, I.; Ritchie, N.D.; Evans, T.J. The interrelationship between phagocytosis, autophagy and formation of neutrophil extracellular traps following infection of human neutrophils by Streptococcus pneumoniae. Innate Immun. 2017, 23, 413–423. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Sequence (5′–3′) |
|---|
| For ATG5NotI: GCGCGGCCGCGCCACCATGACAGA |
| Rev ATG5BamHI: GCGGATCCTCAATCTGTTGC |
| Primer Sequences (5′–3′) | ||
|---|---|---|
| Gene Name | Forward | Reverse |
| ATG5 | GGACGAAACAGCTTCTGAAT | GATGGGATTGCAAAATGACA |
| ACTB | AAATCTGGCACCACACCTTC | GGGGTGTTGAAGGTCTCAAA |
| HPRT | GACCAGTCAACAGGGGACAT | AACACTTCGTGGGGTCCTTTTC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mroczek, A.; Cieloch, A.; Manda-Handzlik, A.; Kuźmicka, W.; Muchowicz, A.; Wachowska, M. Overexpression of ATG5 Gene Makes Granulocyte-Like HL-60 Susceptible to Release Reactive Oxygen Species. Int. J. Mol. Sci. 2020, 21, 5194. https://doi.org/10.3390/ijms21155194
Mroczek A, Cieloch A, Manda-Handzlik A, Kuźmicka W, Muchowicz A, Wachowska M. Overexpression of ATG5 Gene Makes Granulocyte-Like HL-60 Susceptible to Release Reactive Oxygen Species. International Journal of Molecular Sciences. 2020; 21(15):5194. https://doi.org/10.3390/ijms21155194
Chicago/Turabian StyleMroczek, Agnieszka, Adrianna Cieloch, Aneta Manda-Handzlik, Weronika Kuźmicka, Angelika Muchowicz, and Małgorzata Wachowska. 2020. "Overexpression of ATG5 Gene Makes Granulocyte-Like HL-60 Susceptible to Release Reactive Oxygen Species" International Journal of Molecular Sciences 21, no. 15: 5194. https://doi.org/10.3390/ijms21155194
APA StyleMroczek, A., Cieloch, A., Manda-Handzlik, A., Kuźmicka, W., Muchowicz, A., & Wachowska, M. (2020). Overexpression of ATG5 Gene Makes Granulocyte-Like HL-60 Susceptible to Release Reactive Oxygen Species. International Journal of Molecular Sciences, 21(15), 5194. https://doi.org/10.3390/ijms21155194