The Role of the Bacterial Muramyl Dipeptide in the Regulation of GLP-1 and Glycemia

Abstract
:1. Introduction
2. Results
2.1. L18-MDP Stimulates GLP-1 In Vitro
2.2. NOD2 is Expressed in L-cells
2.3. L18-MDP Causes a Slight Increase in p38 MAPK Phosphorylation
2.4. The In Vivo Effects of MDP on GLP-1 Secretion in Chow-Fed Mice
2.5. The In Vivo Effects of MDP on Glucose Tolerance in Obesogenic-Fed Mice
2.6. The Effects of MDP on Fasting GLP-1 Levels
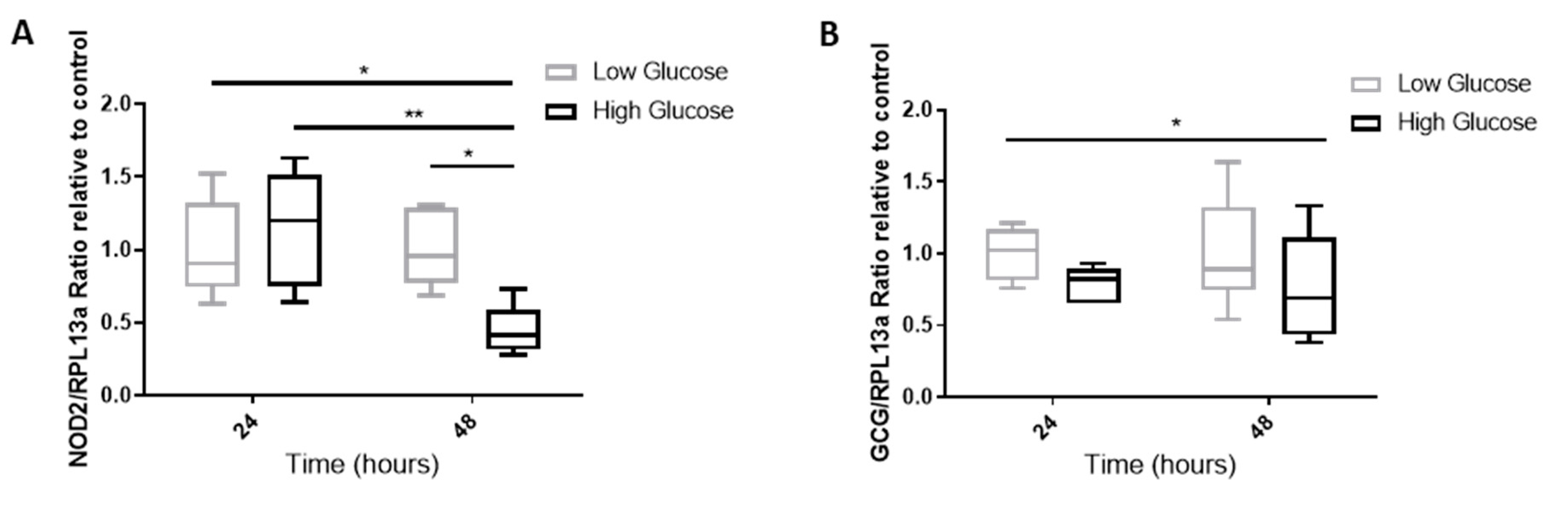
2.7. The Effect of A Hyperglycemic Environment on NOD2 and GCG mRNA Expression
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. In Vivo MDP Experiments
4.2.1. 45% High Fat Diet and Study Design
4.2.2. Western Diet and Study Design
4.3. Oral Glucose Tolerance Test (OGTT)
4.4. Evaluation of GLP-1 and Insulin Hormone Levels
4.5. In Vitro GLP-1 Secretion Experiments
4.6. Gene Expression Analyses
4.7. Fluorescent Immunohistochemistry
4.8. Western Blot
4.9. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AUC | Area under the curve |
| DPP4 | Dipeptidyl peptidase 4 |
| ELISA | Enzyme-linked immunosorbent assay |
| GCG | Proglucagon |
| GLP-1 | Glucagon-like peptide-1 |
| GLP-1R | Glucagon-like peptide-1 receptor |
| HFD | High fat diet |
| IP | Intraperitoneal |
| L18-MDP | 6-O-stearoyl-N-acetyl-muramyl-l-alanine-d-isoglutamine |
| MAPK | Mitogen-activated protein kinase |
| MDP | Muramyl dipeptide |
| mRNA | Messenger ribonucleic acid |
| NAG | N-acetylglucosamine |
| NAM | N-acetylmuramic acid |
| NF-κB | Nuclear factor kappa B |
| NLR | Nucleotide oligomerization domain-like receptor |
| NOD | Nucleotide oligomerization domain |
| OGTT | Oral glucose tolerance test |
| PepT1 | Peptide transporter 1 |
| T2D | Type 2 Diabetes |
| WD | Western diet |
References
- Herrmann, C.; Göke, R.; Richter, G.; Fehmann, H.-C.; Arnold, R.; Göke, B. Glucagon-Like Peptide-1 and Glucose-Dependent Insulin-Releasing Polypeptide Plasma Levels in Response to Nutrients. Digestion 1995, 56, 117–126. [Google Scholar] [CrossRef]
- Spreckley, E.; Murphy, K.G. The L-Cell in Nutritional Sensing and the Regulation of Appetite. Front. Nutr. 2015, 2, 23. [Google Scholar] [CrossRef] [Green Version]
- Baggio, L.L.; Drucker, D.J. Biology of Incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef]
- Rondas, D.; D’Hertog, W.; Overbergh, L.; Mathieu, C. Glucagon-like peptide-1: Modulator of β-cell dysfunction and death. Diabetes Obes. Metab. 2013, 15, 185–192. [Google Scholar] [CrossRef]
- Drucker, D.J. Mechanisms of Action and Therapeutic Application of Glucagon-like Peptide-1. Cell Metabolism. 2018, 27, 740–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eissele, R.; Göke, R.; Willemer, S.; Harthus, H.-P.; Vermeer, H.; Arnold, R.; Göke, B. Glucagon-like peptide-1 cells in the gastrointestinal tract and pancreas of rat, pig and man. Eur. J. Clin. Investig. 1992, 22, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Brial, F.; Le Lay, A.; Dumas, M.-E.; Gauguier, D. Implication of gut microbiota metabolites in cardiovascular and metabolic diseases. Cell Mol. Life Sci. 2018, 75, 3977–3990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Thingholm, L.B.; Rühlemann, M.C.; Koch, M.; Fuqua, B.; Laucke, G.; Boehm, R.; Bang, C.; Franzosa, E.A.; Hübenthal, M.; Rahnavard, A.; et al. Obese Individuals with and without Type 2 Diabetes Show Different Gut Microbial Functional Capacity and Composition. Cell Host Microbe 2019, 26, 252–264.e10. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.M.; Sun, E.W.; Rogers, G.B.; Keating, D.J. The Influence of the Gut Microbiome on Host Metabolism Through the Regulation of Gut Hormone Release. Front. Physiol. 2019, 10, 428. [Google Scholar] [CrossRef] [PubMed]
- Delzenne, N.M.; Neyrinck, A.M.; Bäckhed, F.; Cani, P.D. Targeting gut microbiota in obesity: Effects of prebiotics and probiotics. Nat. Rev. Endocrinol. 2011, 7, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-Chain Fatty Acids Stimulate Glucagon-Like Peptide-1 Secretion via the G-Protein–Coupled Receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chimerel, C.; Emery, E.; Summers, D.K.; Keyser, U.; Gribble, F.M.; Reimann, F. Bacterial Metabolite Indole Modulates Incretin Secretion from Intestinal Enteroendocrine L Cells. Cell Rep. 2014, 9, 1202–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichette, J.; Fynn-Sackey, N.; Gagnon, J. Hydrogen Sulfide and Sulfate Prebiotic Stimulates the Secretion of GLP-1 and Improves Glycemia in Male Mice. Endocrinology 2017, 158, 3416–3425. [Google Scholar] [CrossRef]
- Morimoto, K.; Watanabe, M.; Sugizaki, T.; Irie, J.; Itoh, H. Intestinal Bile Acid Composition Modulates Prohormone Convertase 1/3 (PC1/3) Expression and Consequent GLP-1 Production in Male Mice. Endocrinology 2016, 157, 1071–1081. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Hul, M.V.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Cani, P.D.; Delzenne, N.M.; Amar, J.; Burcelin, R. Role of gut microflora in the development of obesity and insulin resistance following high-fat diet feeding. Pathol. Biol. 2008, 56, 305–309. [Google Scholar] [CrossRef]
- Schleifer, K.H.; Kandler, O. Peptidoglycan types of bacterial cell walls and their taxonomic implications. Bacteriol. Rev. 1972, 36, 407–477. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.S.; Chamaillard, M.; Ogura, Y.; Henegariu, O.; Inohara, N.; Nuñez, G.; Flavell, R.A. Nod2-Dependent Regulation of Innate and Adaptive Immunity in the Intestinal Tract. Science 2005, 307, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Negroni, A.; Pierdomenico, M.; Cucchiara, S.; Stronati, L. NOD2 and Inflammation: Current Insights. Available online: https://www.dovepress.com/nod2-and-inflammation-current-insights-peer-reviewed-fulltext-article-JIR (accessed on 31 March 2019).
- Moltzau Anderson, J.; Lipinski, S.; Sommer, F.; Pan, W.-H.; Boulard, O.; Rehman, A.; Falk-Paulsen, M.; Stengel, S.T.; Aden, K.; Häsler, R.; et al. NOD2 Influences Trajectories of Intestinal Microbiota Recovery After Antibiotic Perturbation. Cell. Mol. Gastroenterol. Hepatol. 2020, 10, 365–389. [Google Scholar] [CrossRef]
- Cavallari, J.F.; Fullerton, M.D.; Duggan, B.M.; Foley, K.P.; Denou, E.; Smith, B.K.; Desjardins, E.M.; Henriksbo, B.D.; Kim, K.J.; Tuinema, B.R.; et al. Muramyl Dipeptide-Based Postbiotics Mitigate Obesity-Induced Insulin Resistance via IRF4. Cell Metab. 2017, 25, 1063–1074.e3. [Google Scholar] [CrossRef]
- Salem, M.; Seidelin, J.B.; Rogler, G.; Nielsen, O.H. Muramyl dipeptide responsive pathways in Crohn’s disease: From NOD2 and beyond. Cell. Mol. Life Sci. 2013, 70, 3391–3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasmuth, S.; Jalilvand, T.V.; Laffer, B.; Busch, M.; Bauer, D.; Langmann, T.; Thanos, S.; Kasper, M.; Heiligenhaus, A. Phenotypic Differences in Primary Murine Microglia Treated with NOD1, NOD2, and NOD1/2 Agonists. Available online: https://www.meta.org/papers/phenotypic-differences-in-primary-murine/31907866 (accessed on 16 July 2020).
- Sidhu, S.S.; Thompson, D.G.; Warhurst, G.; Case, R.M.; Benson, R.S.P. Fatty acid-induced cholecystokinin secretion and changes in intracellular Ca2+ in two enteroendocrine cell lines, STC-1 and GLUTag. J. Physiol. 2000, 528, 165–176. [Google Scholar] [CrossRef]
- Hirasawa, A.; Tsumaya, K.; Awaji, T.; Katsuma, S.; Adachi, T.; Yamada, M.; Sugimoto, Y.; Miyazaki, S.; Tsujimoto, G. Free fatty acids regulate gut incretin glucagon-like peptide-1 secretion through GPR120. Nat. Med. 2005, 11, 90–94. [Google Scholar] [CrossRef]
- Iakoubov, R.; Izzo, A.; Yeung, A.; Whiteside, C.I.; Brubaker, P.L. Protein Kinase Cζ Is Required for Oleic Acid-Induced Secretion of Glucagon-Like Peptide-1 by Intestinal Endocrine L Cells. Endocrinology 2007, 148, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Vavricka, S.R.; Musch, M.W.; Chang, J.E.; Nakagawa, Y.; Phanvijhitsiri, K.; Waypa, T.S.; Merlin, D.; Schneewind, O.; Chang, E.B. hPepT1 Transports Muramyl Dipeptide, Activating NF-κB and Stimulating IL-8 Secretion in Human Colonic Caco2/bbe Cells. Gastroenterology 2004, 127, 9. [Google Scholar] [CrossRef]
- Ismair, M.G.; Vavricka, S.R.; Kullak-Ublick, G.A.; Fried, M.; Mengin-Lecreulx, D.; Girardin, S.E. hPepT1 selectively transports muramyl dipeptide but not Nod1-activating muramyl peptides. Can. J. Physiol. Pharmacol. 2006, 84, 1313–1319. [Google Scholar] [CrossRef]
- Wells, J.M.; Rossi, O.; Meijerink, M.; van Baarlen, P. Epithelial crosstalk at the microbiota-mucosal interface. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4607–4614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lala, S.; Ogura, Y.; Osborne, C.; Hor, S.Y.; Bromfield, A.; Davies, S.; Ogunbiyi, O.; Nuñez, G.; Keshav, S. Crohn’s disease and the NOD2 gene: A role for paneth cells. Gastroenterology 2003, 125, 47–57. [Google Scholar] [CrossRef]
- Windheim, M.; Lang, C.; Peggie, M.; Plater, L.A.; Cohen, P. Molecular mechanisms involved in the regulation of cytokine production by muramyl dipeptide. Biochem. J. 2007, 404, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Lim, G.E.; Brubaker, P.L. Glucagon-Like Peptide 1 Secretion by the L-Cell: The View from Within. Diabetes 2006, 55, S70–S77. [Google Scholar] [CrossRef] [Green Version]
- Hedl, M.; Abraham, C. Nod2-Induced Autocrine Interleukin-1 Alters Signaling by ERK and p38 to Differentially Regulate Secretion of Inflammatory Cytokines. Gastroenterology 2012, 143, 1530–1543. [Google Scholar] [CrossRef] [Green Version]
- Parant, M.; Parant, F.; Chedid, L.; Yapo, A.; Petit, J.F.; Lederer, E. Fate of the synthetic immunoadjuvant, muramyl dipeptide (14C-labelled) in the mouse. Int. J. Immunopharmacol. 1979, 1, 35–41. [Google Scholar] [CrossRef]
- Grasset, E.; Puel, A.; Charpentier, J.; Collet, X.; Christensen, J.E.; Tercé, F.; Burcelin, R. A Specific Gut Microbiota Dysbiosis of Type 2 Diabetic Mice Induces GLP-1 Resistance through an Enteric NO-Dependent and Gut-Brain Axis Mechanism. Cell Metab. 2017, 25, 1075–1090.e5. [Google Scholar] [CrossRef] [Green Version]
- Denou, E.; Lolmède, K.; Garidou, L.; Pomie, C.; Chabo, C.; Lau, T.C.; Fullerton, M.D.; Nigro, G.; Zakaroff-Girard, A.; Luche, E.; et al. Defective NOD2 peptidoglycan sensing promotes diet-induced inflammation, dysbiosis, and insulin resistance. EMBO Mol. Med. 2015, 7, 259–274. [Google Scholar] [CrossRef]
- Repetto, G.; del Peso, A.; Zurita, J. Neutral red uptake assay for the estimation of cell ciability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Gagnon, J.; Mayne, J.; Mbikay, M.; Woulfe, J.; Chrétien, M. Expression of PCSK1 (PC1/3), PCSK2 (PC2) and PCSK3 (furin) in mouse small intestine. Regul. Pept. 2009, 152, 54–60. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward 5′-3′ | Reverse 5′-3′ |
|---|---|---|
| NOD2 | 5′-GTCCAACAATGGCATCACCT-3′ | 5′-TGTGTTCCCTCGAAGCCAAA-3′ |
| GCG | 5′-TTGAGAGGCATGCTGAAGGG-3′ | 5′-TCTTCTGGGAAGTCTCGCCT-3′ |
| RPL13a | 5′-GAAGCAGATCTTGAGGTTACGGA-3′ | 5′-AGGCATGAGGCAAACAGTCT-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, L.; Alshehri, A.; Robichaud, B.; Cudmore, A.; Gagnon, J. The Role of the Bacterial Muramyl Dipeptide in the Regulation of GLP-1 and Glycemia. Int. J. Mol. Sci. 2020, 21, 5252. https://doi.org/10.3390/ijms21155252
Williams L, Alshehri A, Robichaud B, Cudmore A, Gagnon J. The Role of the Bacterial Muramyl Dipeptide in the Regulation of GLP-1 and Glycemia. International Journal of Molecular Sciences. 2020; 21(15):5252. https://doi.org/10.3390/ijms21155252
Chicago/Turabian StyleWilliams, Laura, Amal Alshehri, Bianca Robichaud, Alison Cudmore, and Jeffrey Gagnon. 2020. "The Role of the Bacterial Muramyl Dipeptide in the Regulation of GLP-1 and Glycemia" International Journal of Molecular Sciences 21, no. 15: 5252. https://doi.org/10.3390/ijms21155252