Sialylated Immunoglobulins for the Treatment of Immuno-Inflammatory Diseases

 , , , , and
, , , , and 
{kind=link}
Abstract
1. Introduction
2. Glycosylation of Immunoglobulins: The Effects in Various Diseases
3. Non-Enzymatic Glycosylation and Advanced Glycation End Products
4. Glycosylation Disorders in Rheumatological Diseases
5. Systemic Inflammation and Premature Atherosclerosis
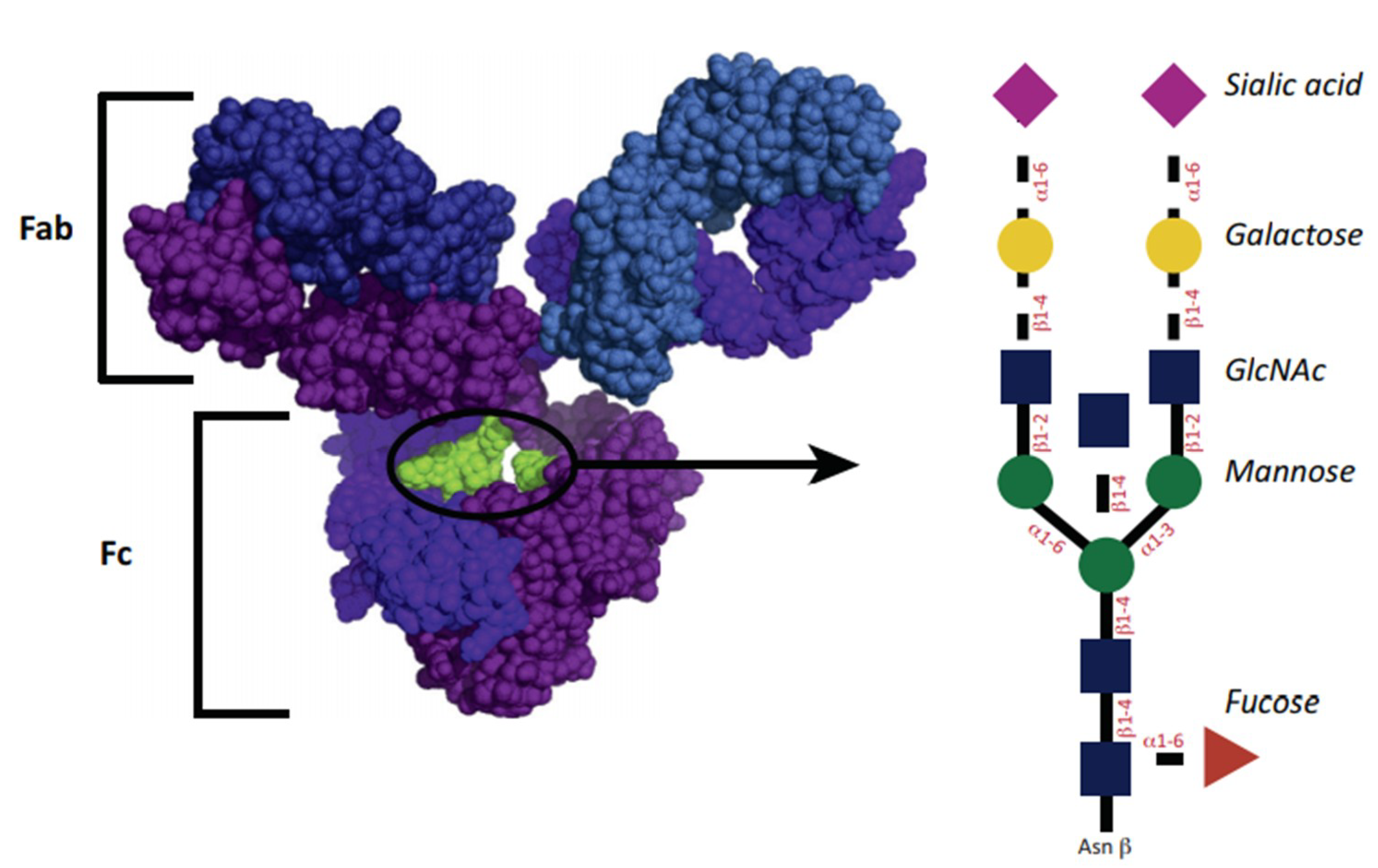
6. Immunoglobulin Fab and Fc Fragment Glycosylation
7. Various Uses of Modified Immunoglobulins in the Treatment of Certain Diseases
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| aAbs | Autoantibodies |
| ACPA | Anti-citrullinated protein antibody |
| ADCC | Antibody-dependent cell cytotoxicity |
| ADCP | Antibody-dependent cell phagocytosis |
| AGEs | Advanced glycation end products |
| CDC | Complement-dependent cytotoxicity |
| CVD | Cardiovascular disease |
| CEL | Nε-(carboxyethyl) lysine |
| CDRs | Complementarity-determining regions |
| CIA | Collagen-induced arthritis |
| CRP | C-reactive protein |
| CVC | Cardiovascular complications |
| CVD | Cardiovascular disease |
| DC | Dendritic cells |
| EBA | Epidermolysis of bullosa acquisita |
| ECs | Endothelial cells |
| Ig | Immunoglobulin |
| IVIg | Intravenous immunoglobulin |
| IL | Interleukin |
| ICs | Immune complexes |
| ITP | Thrombocytopenic purpura |
| HIV | Human immunodeficiency virus |
| LPS | Lipopolysaccharide |
| MBL | Mannose-binding lectin |
| MG | Methylglyoxal |
| PHA | Phytohemagglutinin |
| pDCs | Plasmacytoid dendritic cells |
| RA | Rheumatoid arthritis |
| sVCAM-1 | Soluble vascular cellular activation molecule 1 |
| SpA | Spondyloarthropathy |
| TG | Triglyceride |
| TLA | Three letter acronymn |
| TNF | Tumor necrosis factor |
| VLDL | Very low-density lipoprotein |
References
- Quast, I.; Peschke, B.; Lünemann, J.D. Regulation of antibody effector functions through IgG Fc N-glycosylation. Cell Mol. Life Sci. 2017, 74, 837–847. [Google Scholar] [CrossRef]
- Russell, A.; Adua, E.; Ugrina, I.; Laws, S.; Wang, W. Unravelling Immunoglobulin G Fc NGlycosylation: A Dynamic Marker Potentiating Predictive, Preventive and Personalised Medicine. Int. J. Mol. Sci. 2018, 19, 390. [Google Scholar] [CrossRef]
- Nimmerjahn, F.; Ravetch, J.V. Fc-gamma receptors as regulators of immune responses. Nat. Rev. Immunol. 2008, 8, 34–47. [Google Scholar] [CrossRef]
- Cooper, M.D.; Alder, M.N. The evolution of adaptive immune systems. Cell 2006, 124, 815–822. [Google Scholar] [CrossRef]
- Schroeder, H.W., Jr.; Cavacini, L. Structure and function of immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, 41–52. [Google Scholar] [CrossRef]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG subclasses and allotypes: From structure to effector functions. Front. Immunol. 2014, 5, 520. [Google Scholar] [CrossRef]
- Deisenhofer, J. Crystallographic refinement and atomic models of a human Fc fragment and its complex with fragment B of protein A from Staphylococcus aureus at 2.9- and 2.8-A resolution. Biochemistry 1981, 20, 2361–2370. [Google Scholar] [CrossRef]
- Kaneko, Y.; Nimmerjahn, F.; Ravetch, J.V. Anti-inflammatory activity of immunoglobulin G resulting from Fc sialylation. Science 2006, 313, 670–673. [Google Scholar] [CrossRef]
- Jefferis, R. Glycosylation of recombinant antibody therapeutics. Biotechnol. Prog. 2005, 21, 11–16. [Google Scholar] [CrossRef]
- Jefferis, R. Glycosylation of antibody therapeutics: Optimisation for purpose. Methods Mol. Biol. 2009, 483, 223–238. [Google Scholar]
- Jefferis, R. Glycosylation as a strategy to improve antibody-based therapeutics. Nat. Rev. Drug Discov. 2009, 8, 226–234. [Google Scholar] [CrossRef]
- Ohmi, Y.; Ise, W.; Harazono, A.; Takakura, D.; Fukuyama, H.; Baba, Y.; Narazaki, M.; Shoda, H.; Takahashi, N.; Ohkawa, Y.; et al. Sialylation converts arthritogenic IgG into inhibitors of collagen-induced arthritis. Nat. Commun. 2016, 7, 11205. [Google Scholar] [CrossRef]
- Schwab, I.; Biburger, M.; Krönke, G.; Schett, G.; Nimmerjahn, F. IVIg-mediated amelioration of ITP in mice is dependent on sialic acid and SIGNR1. Eur. J. Immunol. 2012, 42, 826–830. [Google Scholar] [CrossRef]
- Washburn, N.; Schwab, I.; Ortiz, D.; Bhatnagar, N.; Lansing, J.C.; Medeiros, A.; Tyler, S.; Mekala, D.; Cochran, E.; Sarvaiya, H.; et al. Controlled tetra-Fc sialylation of IVIg results in a drug candidate with consistent enhanced anti-inflammatory activity. Proc. Natl. Acad. Sci. USA 2015, 112, 1297–1306. [Google Scholar] [CrossRef]
- Anthony, R.M.; Kobayashi, T.; Wermeling, F.; Ravetch, J.V. Intravenous gammaglobulin suppresses inflammation through a novel T(H)2 pathway. Nature 2011, 475, 110–113. [Google Scholar] [CrossRef]
- Tackenberg, B.; Jelcic, I.; Baerenwaldt, A.; Oertel, W.H.; Sommer, N.; Nimmerjahn, F.; Lünemann, J.D. Impaired inhibitory Fcgamma receptor IIB expression on B cells in chronic inflammatory demyelinating polyneuropathy. Proc. Natl. Acad. Sci. USA 2009, 106, 4788–4792. [Google Scholar] [CrossRef]
- Gudelj, I.; Lauc, G.; Pezer, M. Immunoglobulin G glycosylation in aging and diseases. Cell Immunol. 2018, 333, 65–79. [Google Scholar] [CrossRef]
- Market, E.; Papavasiliou, F.N. V(D)J recombination and the evolution of the adaptive immune system. PLoS Biol. 2003, 1, E16. [Google Scholar] [CrossRef]
- Bournazos, S.; Ravetch, J.V. Fcgamma receptor function and the design of vaccination strategies. Immunity 2017, 47, 224–233. [Google Scholar] [CrossRef]
- Rajewsky, K.; Forster, I.; Cumano, A. Evolutionary and somatic selection of the antibody repertoire in the mouse. Science 1987, 238, 1088–1094. [Google Scholar] [CrossRef]
- Maizels, N. Somatic hypermutation: How many mechanisms diversify V region sequences? Cell 1995, 83, 9–12. [Google Scholar] [CrossRef]
- Nimmerjahn, F.; Ravetch, J.V. Divergent immunoglobulin g subclass activity through selective Fc receptor binding. Science 2005, 310, 1510–1512. [Google Scholar] [CrossRef]
- Mahan, A.E.; Jennewein, M.F.; Suscovich, T.; Dionne, K.; Tedesco, J.; Chung, A.W.; Streeck, H.; Pau, M.; Schuitemaker, H.; Francis, D.; et al. Antigen-Specific Antibody Glycosylation Is Regulated via Vaccination. PLoS Pathog. 2016, 12, e1005456. [Google Scholar] [CrossRef]
- Dwek, R.A. Biological importance of glycosylation. Dev. Biol. Stand. 1998, 96, 43–47. [Google Scholar]
- Nesspor, T.C.; Raju, T.S.; Chin, C.N.; Vafa, O.; Brezski, R.J. Avidity confers FcγR binding and immune effector function to aglycosylated immunoglobulin G1. J. Mol. Recognit. 2012, 25, 147–154. [Google Scholar] [CrossRef]
- Arnold, J.N.; Wormald, M.R.; Sim, R.B.; Rudd, P.M.; Dwek, R.A. The impact of glycosylation on the biological function and structure of human immunoglobulins. Annu. Rev. Immunol. 2007, 25, 21–50. [Google Scholar] [CrossRef]
- Jennewein, M.F.; Alter, G. The immunoregulatory roles of antibody glycosylation. Trends Immunol. 2017, 38, 358–372. [Google Scholar] [CrossRef]
- Alter, G.; Ottenhoff, T.H.M.; Joosten, S.A. Antibody glycosylation in inflammation, disease and vaccination. Semin. Immunol. 2018, 39, 102–110. [Google Scholar] [CrossRef]
- Goulabchand, R.; Vincent, T.; Batteux, F.; Eliaou, J.F.; Guilpain, P. Impact of autoantibody glycosylation in autoimmune diseases. Autoimmun. Rev. 2014, 13, 742–750. [Google Scholar] [CrossRef]
- Schwab, I.; Nimmerjahn, F. Intravenous immunoglobulin therapy: How does IgG modulate the immune system? Nat. Rev. Immunol. 2013, 13, 176–189. [Google Scholar] [CrossRef]
- Yamada, K.; Ito, K.; Furukawa, J.-I.; Nakata, J.; Alvarez, M.; Verbeek, J.S.; Shinohara, Y.; Izui, S. Galactosylation of IgG1 modulates FcgammaRIIB-mediated inhibition of murine autoimmune hemolytic anemia. J. Autoimmun. 2013, 47, 104–110. [Google Scholar] [CrossRef]
- Van de Geijn, F.E.; Wuhrer, M.; Selman, M.H.; Willemsen, S.P.; de Man, Y.A.; Deelder, A.M.; Hazes, J.M. Radboud Jem DolhainImmunoglobulin G galactosylation and sialylation are associated with pregnancy-induced improvement of rheumatoid arthritis and the postpartum flare: Results from a large prospective cohort study. Arthritis Res. Ther. 2009, 11, R193. [Google Scholar] [CrossRef]
- Seeling, M.; Bruckner, C.; Nimmerjahn, F. Differential antibody glycosylation in autoimmunity: Sweet biomarker or modulator of disease activity? Nat. Rev. Rheumatol. 2017, 13, 621–630. [Google Scholar] [CrossRef]
- Gornik, O.; Lauc, G. Glycosylation of serum proteins in inflammatory diseases. Dis. Markers 2008, 25, 26778. [Google Scholar] [CrossRef]
- Van Zeben, D.; Rook, G.A.; Hazes, J.M.; Zwinderman, A.H.; Zhang, Y.; Ghelani, S.; Rademacher, T.W.; Breedveld, F.C. Early agalactosylation of IgG is associated with a more progressive disease course in patients with rheumatoid arthritis: Results of a follow-up study. Br. J. Rheumatol. 1994, 33, 36–43. [Google Scholar] [CrossRef]
- Rademacher, T.W.; Williams, P.; Dwek, R.A. Agalactosyl glycoforms of IgG autoantibodies are pathogenic. Proc. Natl. Acad. Sci. USA 1994, 91, 6123–6127. [Google Scholar] [CrossRef]
- Matsumoto, A.; Shikata, K.; Takeuchi, F.; Kojima, N.; Mizuochi, T. Autoantibody activity of IgG rheumatoid factor increases with decreasing levels of galactosylation and sialylation. J. Biochem. 2000, 128, 621–628. [Google Scholar] [CrossRef]
- Fokkink, W.J.; Selman, M.H.; Dortland, J.R.; Durmus, B.; Kuitwaard, K.; Huizinga, R.; van Rijs, W.; Tio-Gillen, A.P.; van Doorn, P.A.; Deelder, A.M.; et al. IgG Fc N-glycosylation in Guillain-Barre syndrome treated with immunoglobulins. J. Proteome Res. 2014, 13, 1722–1730. [Google Scholar] [CrossRef]
- Kemna, M.J.; Plomp, R.; van Paassen, P.; Koeleman, C.A.M.; Jansen, B.C.; Damoiseaux, J.; Cohen Tervaert, J.W.; Wuhrer, M. Galactosylation and sialylation levels of IgG predict relapse in patients with PR3-ANCA associated vasculitis. EBioMedicine 2017, 17, 108–118. [Google Scholar] [CrossRef]
- Quast, I.; Keller, C.W.; Maurer, M.A.; Giddens, J.P.; Tackenberg, B.; Wang, L.X.; Munz, C.; Nimmerjahn, F.; Dalakas, M.C.; Lunemann, J.D. Sialylation of IgG Fc domain impairs complement-dependent cytotoxicity. J. Clin. Investig. 2015, 125, 4160–4170. [Google Scholar] [CrossRef]
- Ogata, S.; Shimizu, C.; Franco, A.; Touma, R.; Kanegaye, J.T.; Choudhury, B.P.; Naidu, N.N.; Kanda, Y.; Hoang, L.T.; Hibberd, M.L.; et al. Treatment response in Kawasaki disease is associated with sialylation levels of endogenous but not therapeutic intravenous immunoglobulin G. PLoS ONE 2013, 8, e81448. [Google Scholar] [CrossRef]
- Strassheim, D.; Karoor, V.; Stenmark, K.; Verin, A.; Gerasimovskaya, E. A current view of G protein-coupled receptor-mediated signaling in pulmonary hypertension: Finding opportunities for therapeutic intervention. Vessel Plus 2018, 2, 21. [Google Scholar] [CrossRef]
- Anthony, R.M.; Nimmerjahn, F.; Ashline, D.J.; Reinhold, V.N.; Paulson, J.C.; Ravetch, J.V. Recapitulation of IVIG anti-inflammatory activity with a recombinant IgG Fc. Science 2008, 320, 373–376. [Google Scholar] [CrossRef]
- Mehta, A.S.; Long, R.E.; Comunale, M.A.; Wang, M.; Rodemich, L.; Krakover, J.; Philip, R.; Marrero, J.A.; Dwek, R.A.; Block, T.M. Increased levels of galactose-deficient anti-Gal immunoglobulin G in the sera of hepatitis C virus-infected individuals with fibrosis and cirrhosis. J. Virol. 2008, 82, 1259–1270. [Google Scholar] [CrossRef]
- Ackerman, M.E.; Crispin, M.; Baruah, K.; Yu, X.; Boesch, A.W.; Harvey, D.J.; Dugast, A.S.; Heizen, E.L.; Ercan, A.; Choi, I.; et al. Natural variation in Fc glycosylation of HIV-specific antibodies impacts antiviral activity. J. Clin. Investig. 2013, 123, 2183–2192. [Google Scholar] [CrossRef]
- Selman, M.H.; de Jong, S.E.; Soonawala, D.; Kroon, F.P.; Adegnika, A.A.; Deelder, A.M.; Hokke, C.H.; Yazdanbakhsh, M.; Wuhrer, M. Changes in antigen-specific IgG1 Fc N-glycosylation upon influenza and tetanus vaccination. Mol. Cell. Proteom. 2012, 11, M111.014563. [Google Scholar] [CrossRef]
- Vestrheim, A.C.; Moen, A.; Egge-Jacobsen, W.; Reubsaet, L.; Halvorsen, T.G.; Bratlie, D.B.; Paulsen, B.S.; Michaelsen, T.E. A pilot study showing differences in glycosylation patterns of IgG subclasses induced by pneumococcal, meningococcal and two types of influenza vaccines. Immun. Inflamm. Dis. 2014, 2, 76–91. [Google Scholar] [CrossRef]
- Calvo, M.J.; Martínez, M.S.; Torres, W.; Chávez-Castillo, M.; Luzardo, E.; Villasmil, N.; Salazar, J.; Velasco, M.; Bermúdez, V. Omega-3 polyunsaturated fatty acids and cardiovascular health: A molecular view into structure and function. Vessel Plus 2017, 1, 116–128. [Google Scholar] [CrossRef][Green Version]
- Nishiura, T.; Fujii, S.; Kanayama, Y.; Nishikawa, A.; Tomiyama, Y.; Iida, M.; Karasuno, T.; Nakao, H.; Yonezawa, T.; Taniguchi, N.; et al. Carbohydrate analysis of immunoglobulin G myeloma proteins by lectin and high performance liquid chromatography: Role of glycosyltransferases in the structures. Cancer Res. 1990, 50, 5345–5350. [Google Scholar]
- Aurer, I.; Lauc, G.; Dumic, J.; Rendic, D.; Matisic, D.; Milos, M.; Heffer-Lauc, M.; Flogel, M.; Labar, B. Aberrant glycosylation of Igg heavy chain in multiple myeloma. Coll. Antropol. 2007, 31, 247–251. [Google Scholar]
- Fleming, S.C.; Smith, S.; Knowles, D.; Skillen, A.; Self, C.H. Increased sialylation of oligosaccharides on IgG paraproteins—A potential new tumour marker in multiple myeloma. J. Clin. Pathol. 1998, 51, 825–830. [Google Scholar] [CrossRef]
- Mimura, Y.; Ashton, P.R.; Takahashi, N.; Harvey, D.J.; Jefferis, R. Contrasting glycosylation profiles between Fab and Fc of a human IgG protein studied by electrospray ionization mass spectrometry. J. Immunol. Methods 2007, 326, 116–126. [Google Scholar] [CrossRef]
- Mittermayr, S.; Le, G.N.; Clarke, C.; Millan Martin, S.; Larkin, A.M.; O’Gorman, P.; Bones, J. Polyclonal Immunoglobulin G N-Glycosylation in the pathogenesis of plasma cell disorders. J. Proteome Res. 2017, 16, 748–762. [Google Scholar] [CrossRef]
- Bones, J.; Mittermayr, S.; O’Donoghue, N.; Guttman, A.; Rudd, P.M. Ultra performance liquid chromatographic profiling of serum N-glycans for fast and efficient identification of cancer associated alterations in glycosylation. Anal. Chem. 2010, 82, 10208–10215. [Google Scholar] [CrossRef]
- Arnold, J.N.; Saldova, R.; Galligan, M.C.; Murphy, T.B.; Mimura-Kimura, Y.; Telford, J.E.; Godwin, A.K.; Rudd, P.M. Novel glycan biomarkers for the detection of lung cancer. J. Proteome Res. 2011, 10, 1755–1764. [Google Scholar] [CrossRef]
- Alley, W.R., Jr.; Vasseur, J.A.; Goetz, J.A.; Svoboda, M.; Mann, B.F.; Matei, D.E.; Menning, N.; Hussein, A.; Mechref, Y.; Novotny, M.V. N-linked glycan structures and their expressions change in the blood sera of ovarian cancer patients. J. Proteome Res. 2012, 11, 2282–2300. [Google Scholar] [CrossRef]
- Bones, J.; Byrne, J.C.; O’Donoghue, N.; McManus, C.; Scaife, C.; Boissin, H.; Nastase, A.; Rudd, P.M. Glycomic and glycoproteomic analysis of serum from patients with stomach cancer reveals potential markers arising from host defense response mechanisms. J. Proteome Res. 2011, 10, 1246–1265. [Google Scholar] [CrossRef]
- Padarti, A.; Zhang, J. Recent advances in cerebral cavernous malformation research. Vessel Plus 2018, 2, 29. [Google Scholar] [CrossRef]
- Menni, C.; Gudelj, I.; Macdonald-Dunlop, E.; Mangino, M.; Zierer, J.; Bešić, E.; Joshi, P.K.; Trbojević-Akmačić, I.; Chowienczyk, P.; Spector, T.; et al. Glycosylation Profile of Immunoglobulin G Is Cross-Sectionally Associated With Cardiovascular Disease Risk Score and Subclinical Atherosclerosis in Two Independent Cohorts. Circ. Res. 2018, 122, 1555–1564. [Google Scholar] [CrossRef]
- Ji, J.D.; Woo, J.H.; Choi, S.J.; Lee, Y.H.; Song, G.G. Advanced glycation end-products (AGEs): A novel therapeutic target for osteoporosis in patients with rheumatoid arthritis. Med. Hypotheses 2009, 73, 201–202. [Google Scholar] [CrossRef]
- De Groot, L.; Jager, N.A.; Westra, J.; Smit, A.J.; Kallenberg, C.G.M.; Posthumus, M.D.; Bijl, M. Does reduction of disease activity improve early markers of cardiovascular disease in newly diagnosed rheumatoid arthritis patients? Rheumatology 2015, 54, 1257–1261. [Google Scholar] [CrossRef][Green Version]
- Kerkeni, M.; Weiss, I.S.; Jaisson, S.; Dandana, A.; Addad, F.; Gillery, P.; Hammami, M. Increased serum concentrations of pentosidine are related to presence and severity of coronary artery disease. Thromb. Res. 2014, 134, 633–638. [Google Scholar] [CrossRef]
- Haddad, M.; Knani, I.; Bouzidi, H.; Berriche, O.; Hammami, M.; Kerkeni, M. Plasma levels of pentosidine, carboxymethyl lysine, soluble receptor for advanced glycation end products, and metabolic syndrome: The metformin effect. Dis. Markers 2016, 2016, 8. [Google Scholar] [CrossRef]
- Takahashi, T.; Katsuta, S.; Tamura, Y.; Nagase, N.; Suzuki, K.; Nomura, M.; Tomatsu, S.; Miyamoto, K.; Kobayashi, S. Bone-targeting endogenous secretory receptor for advanced glycation end products rescues rheumatoid arthritis. Mol. Med. 2013, 19, 183–194. [Google Scholar] [CrossRef]
- Chavakis, T.; Bierhaus, A.; Al-Fakhri, N.; Schneider, D.; Witte, S.; Linn, T.; Nagashima, M.; Morser, J.; Arnold, B.; Preissner, K.T.; et al. The pattern recognition receptor (RAGE) is a counterreceptor for leukocyte integrins: A novel pathway for infammatory cell recruitment. J. Exp. Med. 2003, 198, 1507–1515. [Google Scholar] [CrossRef]
- Knani, I.; Bouzidi, H.; Zrour, S.; Bergaoui, N.; Hammami, M.; Kerkeni, M. Increased serum concentrations of Nξcarboxymethyllysine are related to presence and severity of rheumatoid arthritis. Ann. Clin. Biochem. 2018, 55, 430–436. [Google Scholar] [CrossRef]
- Pullerits, R.; Bokarewa, M.; Dahlberg, L.; Tarkowski, A. Decreased levels of soluble receptor for advanced glycation end products in patients with rheumatoid arthritis indicating deficient inflammatory control. Arthritis Res. Ther. 2005, 7, R817–R824. [Google Scholar] [CrossRef]
- Knani, I.; Bouzidi, H.; Zrou, S.; Bergaoui, N.; Hammami, M.; Kerkeni, M. Methylglyoxal: A Relevant Marker of Disease Activity in Patients with Rheumatoid Arthritis. Dis. Markers 2018, 2018, 8735926. [Google Scholar] [CrossRef]
- Ma, C.Y.; Ma, J.L.; Jiao, Y.L.; Li, J.-F.; Wang, L.-C.; Yang, Q.-R.; You, L.; Cui, B.; Chen, Z.-J.; Zhao, Y.-R. The plasma level of soluble receptor for advanced glycation end products is decreased in patients with systemic lupus erythematosus. Scand. J. Immunol. 2012, 75, 614–622. [Google Scholar] [CrossRef]
- Manganelli, V.; Truglia, S.; Capozzi, A.; Alessandri, C.; Riitano, G.; Spinelli, F.R.; Ceccarelli, F.; Mancuso, S.; Garofalo, T.; Longo, A.; et al. Alarmin HMGB1 and Soluble RAGE as New Tools to Evaluate the Risk Stratification in Patients With the Antiphospholipid Syndrome. Front. Immunol. 2019, 10, 460. [Google Scholar] [CrossRef]
- Stewart, C.; Cha, S.; Caudle, R.M.; Berg, K.; Katz, J. Decreased levels of soluble receptor for advanced glycation end products in patients with primary Sjogren’s syndrome. Rheumatol. Int. 2008, 28, 771–776. [Google Scholar] [CrossRef]
- Sternberg, Z.; Weinstock-Guttman, B.; Hojnacki, D.; Zamboni, P.; Zivadinov, R.; Chadha, K.; Lieberman, A.; Kazim, L.; Drake, A.; Rocco, P.; et al. Soluble receptor for advanced glycation end products in multiple sclerosis: A potential marker of disease severity. Mult. Scler. 2008, 14, 759–763. [Google Scholar] [CrossRef]
- Jafari-Nakhjavani, M.R.; Jafarpour, M.; Ghorbanihaghjo, A.; Azar, S.A.; Mahdavi, A.M. Relationship between serum-soluble receptor for advanced glycation end products (sRAGE) and disease activity in rheumatoid arthritis patients. Mod. Rheumatol. 2019, 29, 943–948. [Google Scholar] [CrossRef]
- Sobenin, I.A.; Tertov, V.V.; Orekhov, A.N.; Smirnov, V.N. Synergetic effect of desialylated and glycated low density lipoproteins on cholesterol accumulation in cultured smooth muscle intimal cells. Atherosclerosis 1991, 89, 151–154. [Google Scholar] [CrossRef]
- Sobenin, I.A.; Tertov, V.V.; Koschinsky, T.; Buenting, C.E.; Slavina, E.S.; Dedov, I.I.; Orekhov, A.N. Modified low density lipoprotein from diabetic patients causes cholesterol accumulation in human intimal aortic cells. Atherosclerosis 1993, 100, 41–54. [Google Scholar] [CrossRef]
- Sobenin, I.A.; Tertov, V.V.; Orekhov, A.N. Atherogenic modified LDL in diabetes. Diabetes 1996, 45, 35–39. [Google Scholar] [CrossRef]
- Parekh, R.B.; Dwek, R.A.; Sutton, B.J.; Fernandes, D.L.; Leung, A.; Stanworth, D.; Rademacher, T.W.; Mizuochi, T.; Taniguchi, T.; Matsuta, K. Association of rheumatoid arthritis and primary osteoarthritis with changes in the glycosylation pattern of total serum IgG. Nature 1985, 316, 452–457. [Google Scholar] [CrossRef]
- Bodman, K.B.; Sumar, N.; Mackenzie, L.E.; Isenberg, D.A.; Hay, F.C.; Roitt, I.M.; Lydyard, P.M. Lymphocytes from patients with rheumatoid arthritis produce agalactosylated IgG in vitro. Clin. Exp. Immunol. 1992, 88, 420–423. [Google Scholar] [CrossRef]
- Ercan, A.; Cui, J.; Chatterton, D.E.; Deane, K.D.; Hazen, M.M.; Brintnell, W.; O’Donnell, C.I.; Derber, L.A.; Weinblatt, M.E.; Shadick, N.A.; et al. Aberrant IgG galactosylation precedes disease onset, correlates with disease activity, and is prevalent in autoantibodies in rheumatoid arthritis. Arthritis Rheum. 2010, 62, 2239–2248. [Google Scholar] [CrossRef]
- Gudelj, I.; Salo, P.P.; Trbojevic-Akmacic, I.; Albers, M.; Primorac, D.; Perola, M.; Lauc, G. Low galactosylation of IgG associates with higher risk for future diagnosis of rheumatoid arthritis during 10years of follow-up. Biochim. Biophys. Acta 2018, 1864, 2034–2039. [Google Scholar] [CrossRef]
- Martínez, M.S.; García, A.; Luzardo, E.; Chávez-Castillo, M.; Olivar, L.C.; Salazar, J.; Velasco, M.; Rojas Quintero, J.J.; Bermúdez, V. Energetic metabolism in cardiomyocytes: Molecular basis of heart ischemia and arrhythmogenesis. Vessel Plus 2017, 1, 130–141. [Google Scholar] [CrossRef]
- Troelsen, L.N.; Jacobsen, S.; Abrahams, J.L.; Royle, L.; Rudd, P.M.; Narvestad, E.; Heegaard, N.H.; Garred, P. IgG glycosylation changes and MBL2 polymorphisms: Associations with markers of systemic inflammation and joint destruction in rheumatoid arthritis. J. Rheumatol. 2012, 39, 463–469. [Google Scholar] [CrossRef]
- Miranda, S.; Canellada, A.; Gentile, T.; Margni, R. Interleukin-6 and dexamethasone modulate in vitro asymmetric antibody synthesis and UDP-Glc glycoprotein glycosyltransferase activity. J. Reprod. Immunol. 2005, 66, 141–150. [Google Scholar] [CrossRef]
- Scherer, H.U.; van der Woude, D.; Ioan-Facsinay, A.; el Bannoudi, H.; Trouw, L.A.; Wang, J.; Häupl, T.; Burmester, G.-R.; Deelder, A.M.; Huizinga, T.W.J.; et al. Glycan profiling of anti-citrullinated protein antibodies isolated from human serum and synovial fluid. Arthritis Rheum. 2010, 62, 1620–1629. [Google Scholar] [CrossRef]
- Lauc, G.; Huffman, J.E.; Pučić, M.; Zgaga, L.; Adamczyk, B.; Mužinić, A.; Novokmet, M.; Polašek, O.; Gornik, O.; Krištić, J.; et al. Loci associated with N-glycosylation of human immunoglobulin G show pleiotropy with autoimmune diseases and haematological cancers. PLoS Genet. 2013, 9, e1003225. [Google Scholar] [CrossRef]
- Arend, W.P.; Firestein, G.S. Pre-rheumatoid arthritis: Predisposition and transition to clinical synovitis. Nat. Rev. Rheumatol. 2012, 8, 573–586. [Google Scholar] [CrossRef]
- Gornik, I.; Maravic, G.; Dumic, J.; Flogel, M.; Lauc, G. Fucosylation of IgG heavy chains is increased in rheumatoid arthritis. Clin. Biochem. 1999, 32, 605–608. [Google Scholar] [CrossRef]
- Rombouts, Y.; Ewing, E.; van de Stadt, L.A. Anti-citrullinated protein antibodies acquire a pro-inflammatory Fc glycosylation phenotype prior to the onset of rheumatoid arthritis. Ann. Rheum. Dis. 2015, 74, 234–241. [Google Scholar] [CrossRef]
- Engelmann, R.; Brandt, J.; Eggert, M.; Karberg, K.; Krause, A.; Neeck, G.; Mueller-Hilke, B. IgG1 and IgG4 are the predominant subclasses among auto-antibodies against two citrullinated antigens in RA. Rheumatology 2008, 47, 1489–1492. [Google Scholar] [CrossRef]
- Anthony, R.M.; Wermeling, F.; Ravetch, J.V. Novel roles for the IgG Fc glycan. Ann. N. Y. Acad. Sci. 2012, 1253, 170–180. [Google Scholar] [CrossRef]
- Harre, U.; Lang, S.C.; Pfeifle, R.; Rombouts, Y.; Frühbeißer, S.; Amara, K.; Bang, H.; Lux, A.; Koeleman, C.A.; Baum, W.; et al. Glycosylation of immunoglobulin G determines osteoclast differentiation and bone loss. Nat. Commun. 2015, 6, 6651. [Google Scholar] [CrossRef]
- Harre, U.; Georgess, D.; Bang, H.; Bozec, A.; Axmann, R.; Ossipova, E.; Jakobsson, P.-J.; Baum, W.; Nimmerjahn, F.; Szarka, E.; et al. Induction of osteoclastogenesis and bone loss by human autoantibodies against citrullinated vimentin. J. Clin. Investig. 2012, 122, 1791–1802. [Google Scholar] [CrossRef]
- Go, M.F.; Schrohenloher, R.E.; Tomana, M. Deficient galactosylation of serum IgG in inflammatory bowel disease: Correlation with disease activity. J. Clin. Gastroenterol. 1994, 18, 86–87. [Google Scholar] [CrossRef]
- Trbojevic Akmacic, I.; Ventham, N.T.; Theodoratou, E.; Vuckovic, F.; Kennedy, N.A.; Kristic, J.; Nimmo, E.R.; Kalla, R.; Drummond, H.; Stambuk, J.; et al. Inflammatory bowel disease associates with proinflammatory potential of the immunoglobulin G glycome. Inflamm. Bowel Dis. 2015, 21, 1237–1247. [Google Scholar] [CrossRef]
- Espy, C.; Morelle, W.; Kavian, N.; Grange, P.; Goulvestre, C.; Viallon, V.; Chereau, C.; Pagnoux, C.; Michalski, J.C.; Guillevin, L.; et al. Sialylation levels of anti-proteinase 3 antibodies are associated with the activity of granulomatosis with polyangiitis (Wegener’s). Arthritis Rheum. 2011, 63, 2105–2115. [Google Scholar] [CrossRef]
- Gowdar, S.; Syal, S.; Chhabra, L. Probable protective role of diabetes mellitus in takotsubo cardiomyopathy: A review. Vessel Plus 2017, 1, 129–136. [Google Scholar] [CrossRef]
- Mihai, S.; Nimmerjahn, F. The role of Fc receptors and complement in autoimmunity. Autoimmun. Rev. 2013, 12, 657–660. [Google Scholar] [CrossRef]
- Engdahl, C.; Bondt, A.; Harre, U.; Raufer, J.; Pfeifle, R.; Camponeschi, A.; Wuhrer, M.; Seeling, M.; Martensson, I.L.; Nimmerjahn, F.; et al. Estrogen induces St6gal1 expression and increases IgG sialylation in mice and patients with rheumatoid arthritis: A potential explanation for the increased risk of rheumatoid arthritis in postmenopausal women. Arthritis Res. Ther. 2018, 20, 84. [Google Scholar] [CrossRef]
- Chan, A.C.; Carter, P.J. Therapeutic antibodies for autoimmunity and inflammation. Nat. Rev. Immunol. 2010, 10, 301–316. [Google Scholar] [CrossRef]
- Akinkuolie, A.O.; Buring, J.E.; Ridker, P.M.; Mora, S. A novel protein glycan biomarker and future cardiovascular disease events. J. Am. Heart Assoc. 2014, 3, e001221. [Google Scholar] [CrossRef]
- Libby, P. Role of inflammation in atherosclerosis associated with rheumatoid arthritis. Am. J. Med. 2008, 121, S2131. [Google Scholar] [CrossRef] [PubMed]
- Lander, H.M.; Tauras, J.M.; Ogiste, J.S.; Hori, O.; Moss, R.A.; Schmidt, A.M. Activation of the receptor for advanced glycation end products triggers a p21(ras)-dependent mitogen-activated protein kinase pathway regulated by oxidant stress. J. Biol. Chem. 1997, 272, 178104. [Google Scholar] [CrossRef] [PubMed]
- Wahl, A.; van den Akker, E.; Klaric, L.; Štambuk, J.; Benedetti, E.; Plomp, R.; Razdorov, G.; Trbojević-Akmačić, I.; Deelen, J.; van Heemst, D.; et al. GenomeWide Association Study on Immunoglobulin G Glycosylation Patterns. Front. Immunol. 2018, 9, 277. [Google Scholar] [CrossRef] [PubMed]
- Van de Bovenkamp, F.S.; Hafkenscheid, L.; Rispens, T.; Rombouts, Y. The emerging importance of IgG fab glycosylation in immunity. J. Immunol. 2016, 196, 1435–1441. [Google Scholar] [CrossRef]
- Rombouts, Y.; Willemze, A.; Van Beers, J.J.B.C.; Shi, J.; Kerkman, P.F.; Van Toorn, L.; Janssen, G.M.C.; Zaldumbide, A.; Hoeben, R.C.; Pruijn, G.J.M.; et al. Extensive glycosylation of ACPAIgG variable domains modulates binding to citrulli nated antigens in rheumatoid arthritis. Ann. Rheum. 2016, 75, 578585. [Google Scholar] [CrossRef]
- Vletter, E.M.; Koning, M.T.; Scherer, H.U.; Veelken, H.; Toes, R.E.M. A Comparison of Immunoglobulin Variable Region N-Linked Glycosylation in Healthy Donors, Autoimmune Disease and Lymphoma. Front. Immunol. 2020, 11, 241. [Google Scholar] [CrossRef]
- Käsermann, F.; Boerema, D.J.; Ruegsegger, M.; Hofmann, A.; Wymann, S.; Zuercher, A.W.; Miescher, S. Analysis and functional consequences of increased Fab-sialylation of intravenous immunoglobulin (IVIG) after lectin fractionation. PLoS ONE 2012, 7, e37243. [Google Scholar] [CrossRef]
- Wiedeman, A.E.; Santer, D.M.; Yan, W.; Miescher, S.; Käsermann, F.; Elkon, K.B. Contrasting mechanisms of interferon-alpha inhibition by intravenous immunoglobulin after induction by immune complexes versus Toll-like receptor agonists. Arthritis Rheum. 2013, 65, 2713–2723. [Google Scholar]
- Takai, T. Roles of Fc receptors in autoimmunity. Nat. Rev. Immunol. 2002, 2, 580–592. [Google Scholar] [CrossRef]
- Pincetic, A.; Bournazos, S.; DiLillo, D.; Maamary, J.; Wang, T.T.; Dahan, R.; Fiebiger, B.-M.; Ravetch, J.V. Type I and type II Fc receptors regulate innate and adaptive immunity. Nat. Immunol. 2014, 15, 707–716. [Google Scholar] [CrossRef]
- Schwedler, C.; Blanchard, V. Measurement of Neutral and Sialylated IgG N-Glycome at Asn-297 by CE-LIF to Assess Hypogalactosylation in Rheumatoid Arthritis. Methods Mol. Biol. 2019, 1972, 77–93. [Google Scholar] [PubMed]
- Seite, J.F.; Cornec, D.; Renaudineau, Y.; Youinou, P.; Mageed, R.A.; Hillion, S. IVIg modulates BCR signaling through CD22 and promotes apoptosis in mature human B lymphocytes. Blood 2010, 116, 1698–1704. [Google Scholar] [CrossRef] [PubMed]
- Sondermann, P.; Pincetic, A.; Maamary, J.; Lammens, K.; Ravetch, J.V. General mechanism for modulating immunoglobulin effector function. Proc. Natl. Acad. Sci. USA 2013, 110, 9868–9872. [Google Scholar] [CrossRef] [PubMed]
- Massoud, A.H.; Yona, M.; Xue, D.; Chouiali, F.; Alturaihi, H.; Ablona, A.; Mourad, W.; Piccirillo, C.A.; Mazer, B.D. Dendritic cell immunoreceptor: A novel receptor for intravenous immunoglobulin mediates induction of regulatory T cells. J. Allergy Clin. Immunol. 2014, 133, 853–863. [Google Scholar] [CrossRef]
- Haga, C.L.; Ehrhardt Götz, R.A.; Boohaker, R.J.; Davis, R.S.; Cooper, M.D. Fc receptor-like 5 inhibits B cell activation via SHP-1 tyrosine phosphatase recruitment. Proc. Natl. Acad. Sci. USA 2007, 104, 9770–9775. [Google Scholar] [CrossRef]
- Maverakis, E.; Kim, K.; Shimoda, M.; Gershwin, M.E.; Patel, F.; Wilken, R.; Raychaudhuri, S.; Ruhaak, L.R.; Lebrilla, C. Glycans in the immune system and The Altered Glycan Theory of Autoimmunity: A critical review. J. Autoimmun. 2015, 57, 1–13. [Google Scholar] [CrossRef]
- Ehlers, M. Glycosylation-dependent effector function of IgG antibodies. Z. Rheumatol. 2012, 71, 798–800. [Google Scholar] [CrossRef]
- Debre, M.; Bonnet, M.C.; Fridman, W.H.; Carosella, E.; Philippe, N.; Reinert, P.; Vilmer, E.; Kaplan, C.; Teillaud, J.L.; Griscelli, C. Infusion of Fc gamma fragments for treatment of children with acute immune thrombocytopenic purpura. Lancet 1993, 342, 945–949. [Google Scholar] [CrossRef]
- Nandakumar, K.S. Targeting IgG in Arthritis: Disease Pathways and Therapeutic Avenues. Int. J. Mol. Sci. 2018, 19, 677. [Google Scholar] [CrossRef]
- Gómez-Guerrero, C.; Duque, N.; Casado, M.T.; Pastor, C.; Blanco, J.; Mampaso, F.; Vivanco, F.; Egido, J. Administration of IgG Fc fragments prevents glomerular injury in experimental immune complex nephritis. J. Immunol. 2000, 164, 2092–2101. [Google Scholar] [CrossRef]
- Yau, L.F.; Liu, J.; Jiang, M.; Bai, G.; Wang, J.-R.; Jiang, Z.-H. An integrated approach for comprehensive profiling and quantitation of IgG-Fc glycopeptides with application to rheumatoid arthritis. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1122–1123, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Anthony, R.M.; Wermeling, F.; Karlsson, M.C.; Ravetch, J.V. Identification of a receptor required for the anti-inflammatory activity of IVIG. Proc. Natl. Acad. Sci. USA 2008, 105, 19571–19578. [Google Scholar] [CrossRef] [PubMed]
- Durandy, A.; Kaveri, S.V.; Kuijpers, T.W.; Basta, M.; Miescher, S.; Ravetch, J.V.; Rieben, R. Intravenous immunoglobulins—Understanding properties and mechanisms. Clin. Exp. Immunol. 2009, 158, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Pagan, J.D.; Kitaoka, M.; Anthony, R.M. Engineered sialylation of pathogenic antibodies in vivo attenuates autoimmune disease. Cell 2017, 72, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Oefner, C.M.; Winkler, A.; Hess, C.; Lorenz, A.K.; Holecska, V.; Huxdorf, M.; Schommartz, T.; Petzold, D.; Bitterling, J.; Schoen, A.-L.; et al. Tolerance induction with T cell-dependent protein antigens induces regulatory sialylated IgGs. J. Allergy Clin. Immunol. 2012, 129, 1647–1655. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, Y.C.; Rahmöller, J.; Mertes, M.; Eiglmeier, S.; Lorenz, F.K.M.; Stoehr, A.D.; Braumann, D.; Lorenz, A.K.; Winkler, A.; Lilienthal, G.-M.; et al. Sialylated Autoantigen-Reactive IgG Antibodies Attenuate Disease Development in Autoimmune Mouse Models of Lupus Nephritis and Rheumatoid Arthritis. Front. Immunol. 2018, 9, 1183. [Google Scholar] [CrossRef]
- Guo, N.; Liu, Y.; Masuda, Y.; Kawagoe, M.; Ueno, Y.; Kameda, T.; Sugiyama, T. Repeated immunization induces the increase in fucose content on antigen-specific IgG N-linked oligosaccharides. Clin. Biochem. 2005, 38, 149–153. [Google Scholar] [CrossRef]
- Van Beneden, K.; Coppieters, K.; Laroy, W.; De Keyser, F.; Hoffman, I.E.; Van den Bosch, F.; Vander Cruyssen, B.; Drennan, M.; Jacques, P.; Rottiers, P.; et al. Reversible changes in serum immunoglobulin galactosylation during the immune response and treatment of inflammatory autoimmune arthritis. Ann. Rheum. Dis. 2009, 68, 13605. [Google Scholar] [CrossRef]
- Croce, A.; Firuzi, O.; Altieri, F.; Eufemi, M.; Agostino, R.; Priori, R.; Bombardieri, M.; Alessandri, C.; Valesini, G.; Sasocorresponding, L. Effect of infliximab on the glycosylation of IgG of patients with rheumatoid arthritis. J. Clin. Lab. Anal. 2007, 21, 30314. [Google Scholar] [CrossRef]
- Pasek, M.; Duk, M.; Podbielska, M.; Sokolik, R.; Szechiński, J.; Lisowska, E.; Krotkiewski, H. Galactosylation of IgG from rheumatoid arthritis (RA) patients—Changes during therapy. Glycoconj. J. 2006, 23, 46371. [Google Scholar] [CrossRef]
- Ercan, A.; Cui, J.; Hazen, M.M.; Batliwalla, F.; Royle, L.; Rudd, P.M.; Coblyn, J.S.; Shadick, N.; Weinblatt, M.E.; Gregersen, P.; et al. Hypogalactosylation of serum N-glycans fails to predict clinical response to methotrexate and TNF inhibition in rheumatoid arthritis. Arthritis Res. Ther. 2012, 14, R43. [Google Scholar] [CrossRef] [PubMed]
- Collins, E.S.; Galligan, M.C.; Saldova, R.; Adamczyk, B.; Abrahams, J.L.; Campbell, M.P.; Ng, C.-T.; Veale, D.J.; Murphy, T.B.; Rudd, P.M.; et al. Glycosylation status of serum in inflammatory arthritis in response to anti-TNF treatment. Rheumatology 2013, 52, 1572–1582. [Google Scholar] [CrossRef] [PubMed]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markina, Y.V.; Gerasimova, E.V.; Markin, A.M.; Glanz, V.Y.; Wu, W.-K.; Sobenin, I.A.; Orekhov, A.N. Sialylated Immunoglobulins for the Treatment of Immuno-Inflammatory Diseases. Int. J. Mol. Sci. 2020, 21, 5472. https://doi.org/10.3390/ijms21155472
Markina YV, Gerasimova EV, Markin AM, Glanz VY, Wu W-K, Sobenin IA, Orekhov AN. Sialylated Immunoglobulins for the Treatment of Immuno-Inflammatory Diseases. International Journal of Molecular Sciences. 2020; 21(15):5472. https://doi.org/10.3390/ijms21155472
Chicago/Turabian StyleMarkina, Yuliya V., Elena V. Gerasimova, Alexander M. Markin, Victor Y. Glanz, Wei-Kai Wu, Igor A. Sobenin, and Alexander N. Orekhov. 2020. "Sialylated Immunoglobulins for the Treatment of Immuno-Inflammatory Diseases" International Journal of Molecular Sciences 21, no. 15: 5472. https://doi.org/10.3390/ijms21155472
APA StyleMarkina, Y. V., Gerasimova, E. V., Markin, A. M., Glanz, V. Y., Wu, W.-K., Sobenin, I. A., & Orekhov, A. N. (2020). Sialylated Immunoglobulins for the Treatment of Immuno-Inflammatory Diseases. International Journal of Molecular Sciences, 21(15), 5472. https://doi.org/10.3390/ijms21155472