Adenovirus-Mediated FasL Minigene Transfer Endows Transduced Cells with Killer Potential


 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
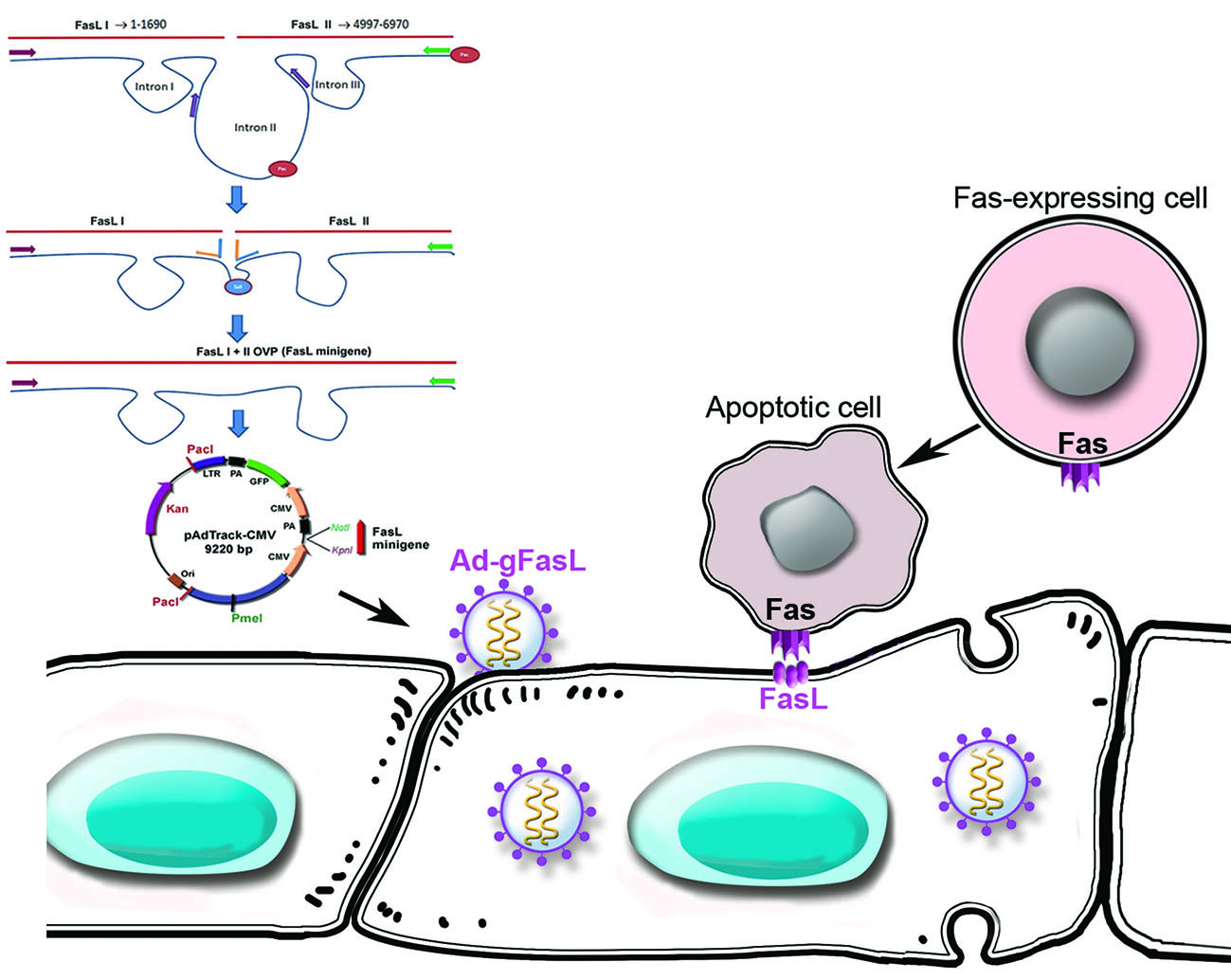
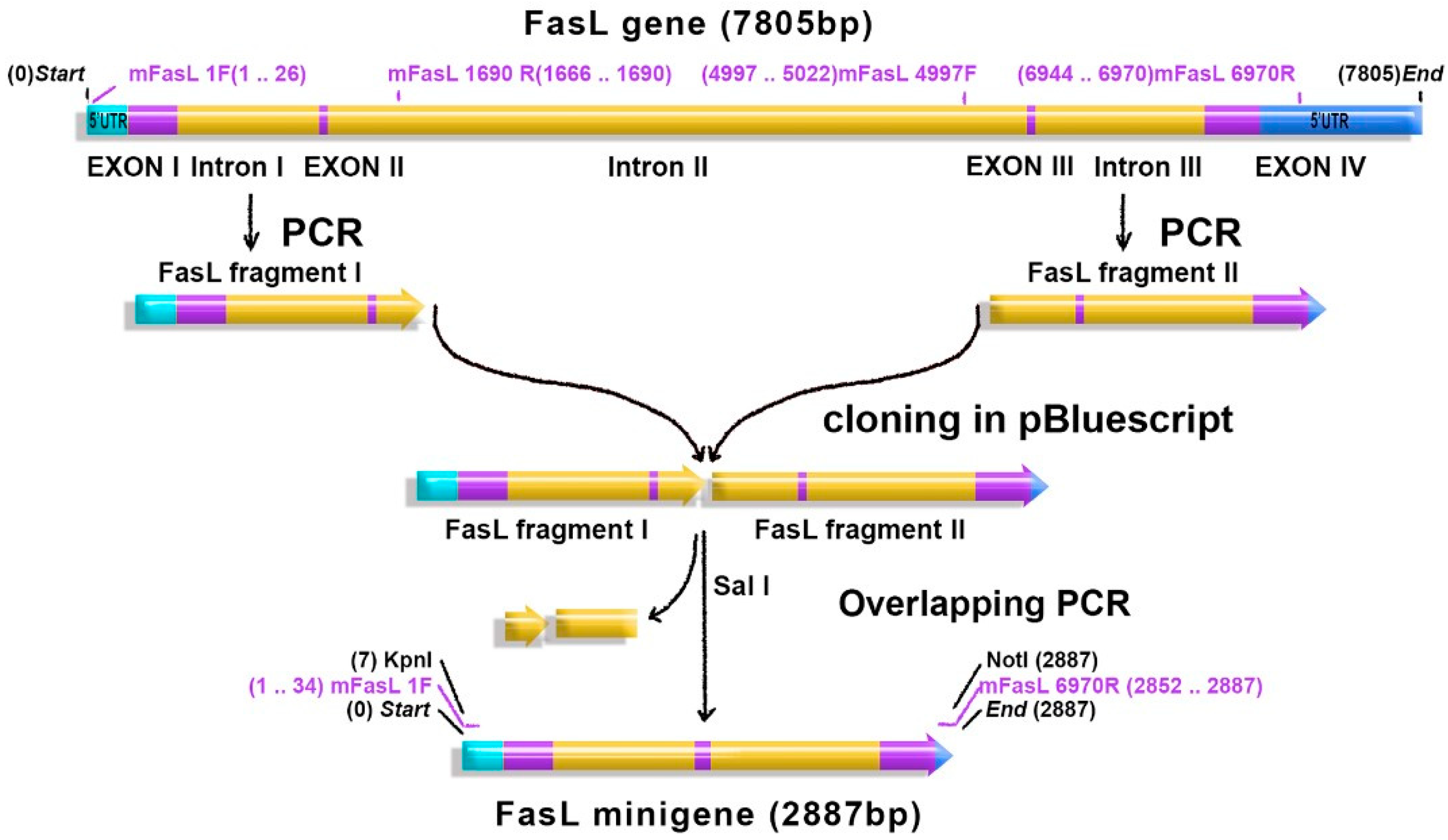
2.1. FasL Minigene and FasL cDNA Carrying Adenoviruses
2.2. FasL Expression in Transduced Cells
2.3. Functional FasL Expression in Transduced Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. DNA Isolation, Cloning, and Sequencing
4.3. FasL Adenovirus Packaging and Purification
4.4. Titration of Adenoviral Vectors
4.5. Viral Transduction for FasL Expression
4.6. RT-PCR
4.7. Western Blot
4.8. Flow Cytometric Analysis of FasL Expressed on the Cell Surface
4.9. Apoptosis Assay in Mixed Cultures
5. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FasL | First apoptosis signal ligand |
| GFP | Green Fluorescent Protein |
| FADD | Fas-associated death domain |
| BAEC | Bovine aortic endothelial cells |
References
- Lowin, B.; Hahne, M.; Mattmann, C.; Tschopp, J. Cytolytic T-cell cytotoxicity is mediated through perforin and Fas lytic pathways. Nature 1994, 370, 650–652. [Google Scholar] [CrossRef]
- Kagi, D.; Vignaux, F.; Ledermann, B.; Burki, K.; Depraetere, V.; Nagata, S.; Hengartner, H.; Golstein, P. Fas and perforin pathways as major mechanisms of T cell-mediated cytotoxicity. Science 1994, 265, 528–530. [Google Scholar] [CrossRef]
- Schneider, P.; Bodmer, J.L.; Holler, N.; Mattmann, C.; Scuderi, P.; Terskikh, A.; Peitsch, M.C.; Tschopp, J. Characterization of Fas (Apo-1, CD95)-Fas ligand interaction. J. Biol. Chem. 1997, 272, 18827–18833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; Itai, T.; Adachi, M.; Nagata, S. Downregulation of Fas ligand by shedding. Nat. Med. 1998, 4, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Alderson, M.R.; Tough, T.W.; Davis-Smith, T.; Braddy, S.; Falk, B.; Schooley, K.A.; Goodwin, R.G.; Smith, C.A.; Ramsdell, F.; Lynch, D.H. Fas ligand mediates activation-induced cell death in human T lymphocytes. J. Exp. Med. 1995, 181, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffith, T.S.; Brunner, T.; Fletcher, S.M.; Green, D.R.; Ferguson, T.A. Fas ligand-induced apoptosis as a mechanism of immune privilege. Science 1995, 270, 1189–1192. [Google Scholar] [CrossRef]
- Kaminitz, A.; Yolcu, E.S.; Stein, J.; Yaniv, I.; Shirwan, H.; Askenasy, N. Killer Treg restore immune homeostasis and suppress autoimmune diabetes in prediabetic NOD mice. J. Autoimmun. 2011, 37, 39–47. [Google Scholar] [CrossRef]
- Lau, H.T.; Yu, M.; Fontana, A.; Stoeckert, C.J., Jr. Prevention of islet allograft rejection with engineered myoblasts expressing FasL in mice. Science 1996, 273, 109–112. [Google Scholar] [CrossRef]
- Yolcu, E.S.; Zhao, H.; Bandura-Morgan, L.; Lacelle, C.; Woodward, K.B.; Askenasy, N.; Shirwan, H. Pancreatic islets engineered with SA-FasL protein establish robust localized tolerance by inducing regulatory T cells in mice. J. Immunol. 2011, 187, 5901–5909. [Google Scholar] [CrossRef] [Green Version]
- Zhan, H.G.; Mountz, J.D.; Fleck, M.; Zhou, T.; Hsu, H.C. Specific deletion of autoreactive T cells by adenovirus-transfected, Fas ligand-producing antigen-presenting cells. Immunol. Res. 2002, 26, 235–246. [Google Scholar] [CrossRef]
- Yolcu, E.S.; Askenasy, N.; Singh, N.P.; Cherradi, S.E.; Shirwan, H. Cell membrane modification for rapid display of proteins as a novel means of immunomodulation: FasL-decorated cells prevent islet graft rejection. Immunity 2002, 17, 795–808. [Google Scholar] [CrossRef] [Green Version]
- Kaminitz, A.; Yolcu, E.S.; Mizrahi, K.; Shirwan, H.; Askenasy, N. Killer Treg cells ameliorate inflammatory insulitis in non-obese diabetic mice through local and systemic immunomodulation. Int. Immunol. 2013, 25, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Kaminitz, A.; Askenasy, N.; Yolcu, E.S. Immunomodulation with regulatory T cells and Fas-ligand ameliorate established inflammatory colitis. Gut 2013, 62, 1228–1230. [Google Scholar] [CrossRef] [PubMed]
- Yolcu, E.S.; Kaminitz, A.; Mizrahi, K.; Ash, S.; Yaniv, I.; Stein, J.; Shirwan, H.; Askenasy, N. Immunomodulation with donor regulatory T cells armed with Fas-ligand alleviates graft-versus-host disease. Exp. Hematol. 2013, 41, 903–911. [Google Scholar] [CrossRef]
- Franke, D.D.; Yolcu, E.S.; Alard, P.; Kosiewicz, M.M.; Shirwan, H. A novel multimeric form of FasL modulates the ability of diabetogenic T cells to mediate type 1 diabetes in an adoptive transfer model. Mol. Immunol. 2007, 44, 2884–2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianco, S.R.; Sun, J.; Fosmire, S.P.; Hance, K.; Padilla, M.L.; Ritt, M.G.; Getzy, D.M.; Duke, R.C.; Withrow, S.J.; Lana, S.; et al. Enhancing antimelanoma immune responses through apoptosis. Cancer Gene Ther. 2003, 10, 726–736. [Google Scholar] [CrossRef]
- Modiano, J.F.; Bellgrau, D. Fas ligand based immunotherapy: A potent and effective neoadjuvant with checkpoint inhibitor properties, or a systemically toxic promoter of tumor growth? Discov. Med. 2016, 21, 109–116. [Google Scholar]
- Luo, X.; Miller, S.D.; Shea, L.D. Immune Tolerance for Autoimmune Disease and Cell Transplantation. Annu. Rev. Biomed. Eng. 2016, 18, 181–205. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Chen, M.; Pettersson, U. A new look at adenovirus splicing. Virology 2014, 456–457, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Biasiotto, R.; Akusjärvi, G. Regulation of human adenovirus alternative RNA splicing by the adenoviral L4-33K and L4-22K proteins. Int. J. Mol. Sci. 2015, 16, 2893–2912. [Google Scholar] [CrossRef] [Green Version]
- Kayagaki, N.; Yamaguchi, N.; Nagao, F.; Matsuo, S.; Maeda, H.; Okumura, K.; Yagita, H. Polymorphism of murine Fas ligand that affects the biological activity. Proc. Natl. Acad. Sci. USA 1997, 94, 3914–3919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, T.C.; Zhou, S.; da Costa, L.T.; Yu, J.; Kinzler, K.W.; Vogelstein, B. A simplified system for generating recombinant adenoviruses. Proc. Natl. Acad. Sci. USA 1998, 95, 2509–2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Dijk, K.W.; Kypreos, K.E.; Fallaux, F.J.; Hageman, J. Adenovirus-mediated gene transfer. Methods Mol. Biol. 2011, 693, 321–343. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Smith, S.K.; Siegel, F.; Shi, Z.; Van Kampen, K.R.; Elmets, C.A.; Tang, D.C. AdEasy system made easier by selecting the viral backbone plasmid preceding homologous recombination. Biotechniques 2001, 31, 260–262. [Google Scholar] [CrossRef] [Green Version]
- Larregina, A.T.; Morelli, A.E.; Dewey, R.A.; Castro, M.G.; Fontana, A.; Lowenstein, P.R. FasL induces Fas/Apo1-mediated apoptosis in human embryonic kidney 293 cells routinely used to generate E1-deleted adenoviral vectors. Gene Ther. 1998, 5, 563–568. [Google Scholar] [CrossRef]
- Liu, J.Y.; Wang, S.M.; Chen, I.C.; Yu, C.K.; Liu, C.C. Hepatic damage caused by coxsackievirus B3 is dependent on age-related tissue tropisms associated with the coxsackievirus-adenovirus receptor. Pathog. Dis. 2013, 68, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Zannis, V.I.; Chroni, A.; Kypreos, K.E.; Kan, H.Y.; Cesar, T.B.; Zanni, E.E.; Kardassis, D. Probing the pathways of chylomicron and HDL metabolism using adenovirus-mediated gene transfer. Curr.Opin. Lipidol. 2004, 15, 151–166. [Google Scholar] [CrossRef]
- Kypreos, K.E.; Zannis, V.I. Pathway of biogenesis of apolipoprotein E-containing HDL in vivo with the participation of ABCA1 and LCAT. Biochem. J. 2007, 403, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Woodward, K.B.; Zhao, H.; Shrestha, P.; Batra, L.; Tan, M.; Grimany-Nuno, O.; Bandura-Morgan, L.; Askenasy, N.; Shirwan, H.; Yolcu, E.S. Pancreatic islets engineered with a FasL protein induce systemic tolerance at the induction phase that evolves into long-term graft-localized immune privilege. Am. J. Transplant. 2020, 20, 1285–1295. [Google Scholar] [CrossRef]
- Desbarats, J.; Newell, M.K. Fas engagement accelerates liver regeneration after partial hepatectomy. Nat. Med. 2000, 6, 920–923. [Google Scholar] [CrossRef]
- Fechner, H.; Haack, A.; Wang, H.; Wang, X.; Eizema, K.; Pauschinger, M.; Schoemaker, R.; Veghel, R.; Houtsmuller, A.; Schultheiss, H.P.; et al. Expression of coxsackie adenovirus receptor and alphav-integrin does not correlate with adenovector targeting in vivo indicating anatomical vector barriers. Gene Ther. 1999, 6, 1520–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangari, D.S.; Shukla, S.; Mittal, S.K. Comparative transduction efficiencies of human and nonhuman adenoviral vectors in human, murine, bovine, and porcine cells in culture. Biochem. Biophys. Res. Commun. 2005, 327, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Richardson, B.C.; Lalwani, N.D.; Johnson, K.J.; Marks, R.M. Fas ligation triggers apoptosis in macrophages but not endothelial cells. Eur. J. Immunol. 1994, 24, 2640–2645. [Google Scholar] [CrossRef] [PubMed]
- Gafencu, A.; Stanescu, M.; Toderici, A.M.; Heltianu, C.; Simionescu, M. Protein and fatty acid composition of caveolae from apical plasmalemma of aortic endothelial cells. Cell Tissue Res. 1998, 293, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Valiente, M.; Obenauf, A.C.; Jin, X.; Chen, Q.; Zhang, X.H.; Lee, D.J.; Chaft, J.E.; Kris, M.G.; Huse, J.T.; Brogi, E.; et al. Serpins promote cancer cell survival and vascular co-option in brain metastasis. Cell 2014, 156, 1002–1016. [Google Scholar] [CrossRef] [Green Version]
- Trusca, V.G.; Fuior, E.V.; Kardassis, D.; Simionescu, M.; Gafencu, A.V. The Opposite Effect of c-Jun Transcription Factor on Apolipoprotein E Gene Regulation in Hepatocytes and Macrophages. Int. J. Mol. Sci. 2019, 20, 1471. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dumitrescu, M.; Trusca, V.G.; Savu, L.; Stancu, I.G.; Ratiu, A.C.; Simionescu, M.; Gafencu, A.V. Adenovirus-Mediated FasL Minigene Transfer Endows Transduced Cells with Killer Potential. Int. J. Mol. Sci. 2020, 21, 6011. https://doi.org/10.3390/ijms21176011
Dumitrescu M, Trusca VG, Savu L, Stancu IG, Ratiu AC, Simionescu M, Gafencu AV. Adenovirus-Mediated FasL Minigene Transfer Endows Transduced Cells with Killer Potential. International Journal of Molecular Sciences. 2020; 21(17):6011. https://doi.org/10.3390/ijms21176011
Chicago/Turabian StyleDumitrescu, Madalina, Violeta Georgeta Trusca, Lorand Savu, Ioana Georgeta Stancu, Attila Cristian Ratiu, Maya Simionescu, and Anca Violeta Gafencu. 2020. "Adenovirus-Mediated FasL Minigene Transfer Endows Transduced Cells with Killer Potential" International Journal of Molecular Sciences 21, no. 17: 6011. https://doi.org/10.3390/ijms21176011
APA StyleDumitrescu, M., Trusca, V. G., Savu, L., Stancu, I. G., Ratiu, A. C., Simionescu, M., & Gafencu, A. V. (2020). Adenovirus-Mediated FasL Minigene Transfer Endows Transduced Cells with Killer Potential. International Journal of Molecular Sciences, 21(17), 6011. https://doi.org/10.3390/ijms21176011