Leucocyte-Rich Platelet-Rich Plasma Enhances Fibroblast and Extracellular Matrix Activity: Implications in Wound Healing

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Haematological Values: Whole Blood, LR-PRP and LP-PPP
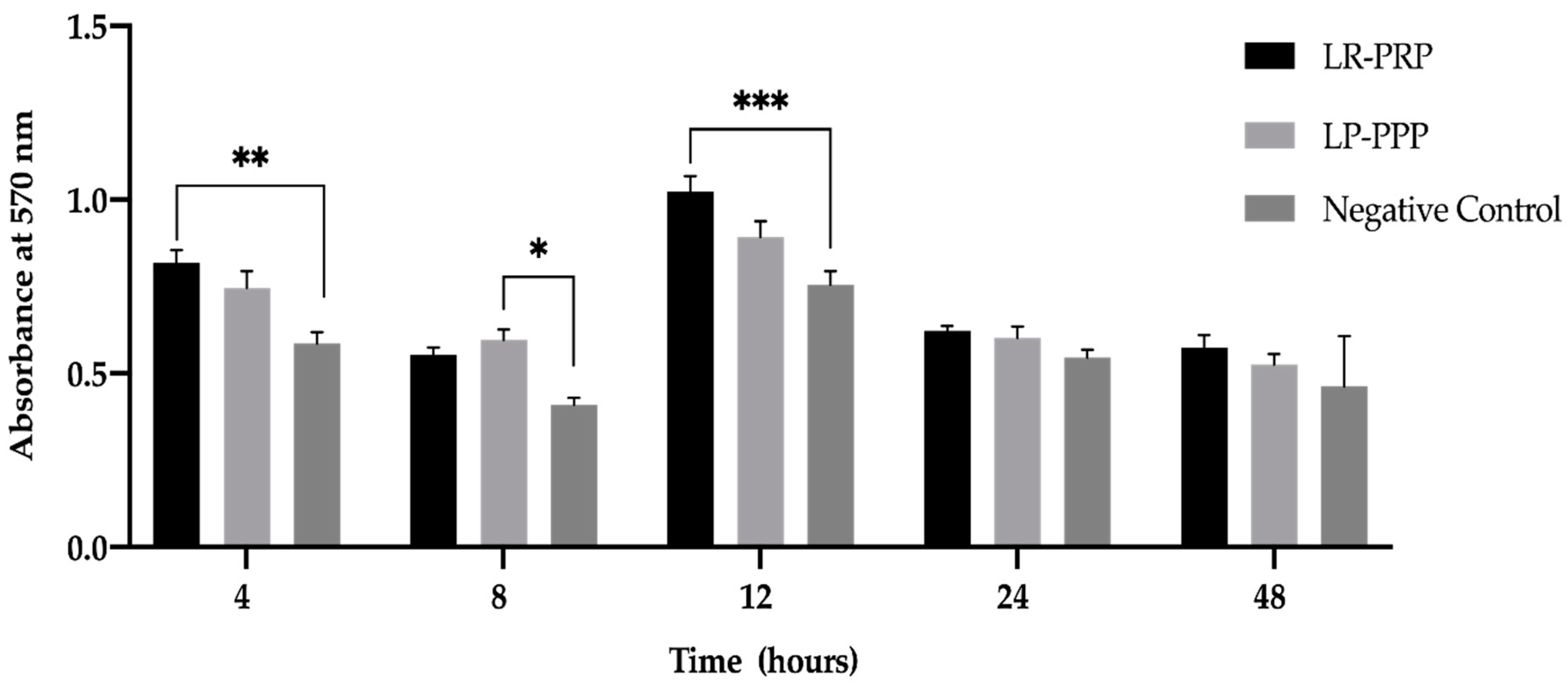
2.2. LR-PRP and LP-PPP Enhance Proliferation of Fibroblast Cells
2.3. LR-PRP and LP-PPP Enhance Fibroblast Migration in Wound Scratch Assay
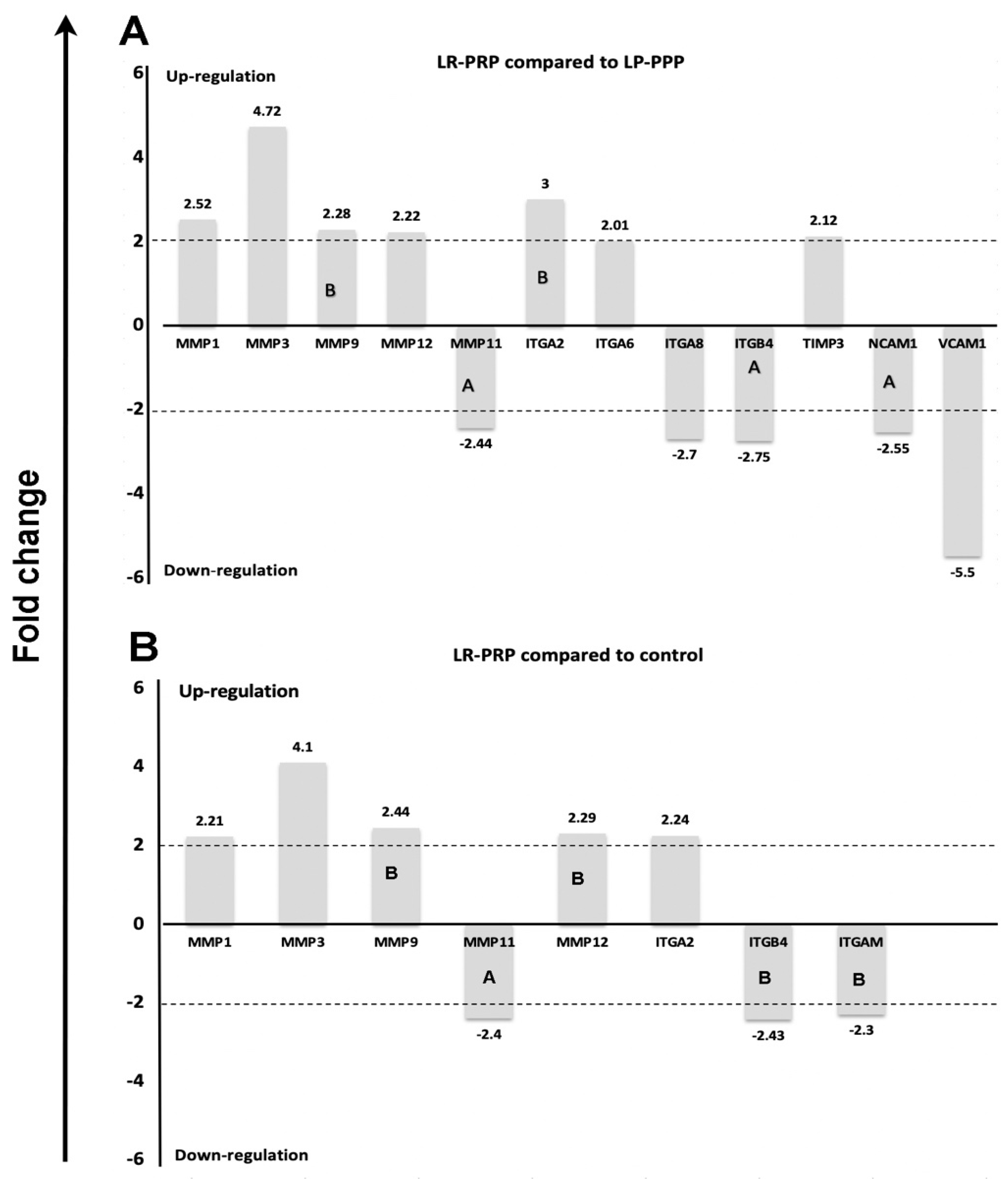
2.4. Expression of mRNA Genes of Human ECM and Adhesion Molecules
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. Analysis of Leucocyte-Rich, Platelet-Rich Plasma and Leucocyte-Poor, Platelet-Poor Plasma
4.3. Cell Culture
4.4. Fibroblast Proliferation Method Using MTT
4.5. In Vitro Scratch Wound Assay
4.6. The Human ECM and Adhesion Molecules RT2 Profiler PCR Gene Array
4.7. Data Analysis Using Prism Programs and Qiagen Software Programs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Availability of Data and Materials
Abbreviations
| ECM | Extracellular matrix |
| ITG | Integrin |
| ITGA | Integrin alpha |
| ITGB4 | Integrin beta 4 |
| ITGAM | Integrin alpha M chain |
| LP-PPP | Leucocyte-poor, platelet poor plasma |
| LR-PRP | Leucocyte-rich, platelet rich plasma |
| MMP | Matrix metalloproteinase |
| MMP1 | Matrix metallopeptidase 1 (interstitial collagenase) |
| MMP3 | Matrix metallopeptidase (stromelysin 1, progelatinase) |
| MMP9 | Matrix metallopeptidase (gelatinase, B.; 92kDa gelatinase, 92kDa type IV collagenase) |
| MMP11 | Matrix metallopeptidase (stromelysin 3) |
| MMP12 | Matrix metallopeptidase (macrophage elastase, metalloelastase) |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide, a tetrazole |
| NCAM1 | Neural cell adhesion molecule |
| PRP | Platelet rich plasma |
| RBC | Red blood cell |
| TIMP3 | Tissue inhibitors of metalloproteinases |
| TGF-β | Transforming growth factor beta |
| VCAM1 | Vascular cell adhesion molecule 1 |
| WBC | White blood cell |
References
- Roffi, A.; Filardo, G.; Assirelli, E.; Cavallo, C.; Cenacchi, A.; Facchini, A.; Grigolo, B.; Kon, E.; Mariani, E.; Pratelli, L.; et al. Does platelet-rich plasma freeze-thawing influence growth factor release and their effects on chondrocytes and synoviocytes? Biomed. Res. Int. 2014, 2014, 692913. [Google Scholar] [CrossRef] [PubMed]
- Giusti, I.; D’Ascenzo, S.; Mancò, A.; Di Stefano, G.; Di Francesco, M.; Rughetti, A.; Dal Mas, A.; Properzi, G.; Calvisi, V.; Dolo, V. Platelet concentration in platelet-rich plasma affects tenocyte behavior in vitro. Biomed. Res. Int. 2014, 2014, 630870. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.; Qi, X.; Zhang, Y.; Sheng, J.; Xu, Z.; Tao, S.; Xie, X.; Li, X.; Zhang, C. Advantages of pure platelet-rich plasma compared with leukocyte- and platelet-rich plasma in promoting repair of bone defects. J. Transl Med. 2016, 14, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalon, J.; Velier, M.; Francois, P.; Graiet, H.; Veran, J.; Sabatier, F. Comment on “responders to platelet-rich plasma in osteoarthritis: A technical analysis”. Biomed. Res. Int. 2017, 2017, 8620257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrangelo, A.N.; Vavken, P.; Fleming, B.C.; Harrison, S.L.; Murray, M.M. Reduced platelet concentration does not harm prp effectiveness for acl repair in a porcine in vivo model. J. Orthop. Res. 2011, 29, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.J.; Xu, H.T.; Sheng, J.G.; An, Z.Q.; Guo, S.C.; Xie, X.T.; Zhang, C.Q. Advantages of pure platelet-rich plasma compared with leukocyte- and platelet-rich plasma in treating rabbit knee osteoarthritis. Med. Sci. Monit. 2016, 22, 1280–1290. [Google Scholar] [CrossRef] [Green Version]
- Wirohadidjojo, Y.W.; Budiyanto, A.; Soebono, H. Platelet-rich fibrin lysate can ameliorate dysfunction of chronically uva-irradiated human dermal fibroblasts. Yonsei Med. J. 2016, 57, 1282–1285. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Zapata, M.J.; Martí-Carvajal, A.J.; Solà, I.; Expósito, J.A.; Bolíbar, I.; Rodríguez, L.; Garcia, J.; Zaror, C. Autologous platelet-rich plasma for treating chronic wounds. Cochrane Database Syst. Rev. 2016, 5, CD006899. [Google Scholar] [CrossRef]
- Mazzucco, L.; Balbo, V.; Guaschino, R. “Reasonable compromise” to define the quality standards of platelet concentrate for non-transfusion use (cpunt). Transfus. Apher. Sci. 2012, 47, 207–211. [Google Scholar] [CrossRef]
- Bielecki, T.; Dohan Ehrenfest, D.M.; Everts, P.A.; Wiczkowski, A. The role of leukocytes from l-prp/l-prf in wound healing and immune defense: New perspectives. Curr. Pharm. Biotechnol. 2012, 13, 1153–1162. [Google Scholar] [CrossRef]
- Riboh, J.C.; Saltzman, B.M.; Yanke, A.B.; Fortier, L.; Cole, B.J. Effect of leukocyte concentration on the efficacy of platelet-rich plasma in the treatment of knee osteoarthritis. Am. J. Sports Med. 2016, 44, 792–800. [Google Scholar] [CrossRef] [PubMed]
- McCarrel, T.M.; Minas, T.; Fortier, L.A. Optimization of leukocyte concentration in platelet-rich plasma for the treatment of tendinopathy. J. Bone Joint Surg. Am. 2012, 94, e143. [Google Scholar] [CrossRef]
- Kuffler, D.P. Variables affecting the potential efficacy of prp in providing chronic pain relief. J. Pain Res. 2019, 12, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khorshidi, H.; Raoofi, S.; Bagheri, R.; Banihashemi, H. Comparison of the mechanical properties of early leukocyte- and platelet-rich fibrin versus prgf/endoret membranes. Int. J. Dent. 2016, 1849207. [Google Scholar] [CrossRef] [Green Version]
- Anitua, E.; Zalduendo, M.; Troya, M.; Padilla, S.; Orive, G. Leukocyte inclusion within a platelet rich plasma-derived fibrin scaffold stimulates a more pro-inflammatory environment and alters fibrin properties. PLoS ONE 2015, 10, 1932–6203. [Google Scholar] [CrossRef]
- Everts, P.A.M.; Knape, J.T.A.; Weibrich, G.; Schönberger, J.P.; Hoffmann, J.; Overdevest, E.P.; Box, H.A.; van Zundert, A. Platelet rich plasma and platelet gel, a review. J. Extra Corpor. Technol. 2006, 38, 174–187. [Google Scholar]
- Ozer, K.; Kankaya, Y.; Colak, O. An important and overlooked parameter in platelet rich plasma preparation: The mean platelet volume. J. Cosmet. Dermatol. 2018. [Google Scholar] [CrossRef]
- Magalon, J.; Chateau, A.L.; Bertrand, B.; Louis, M.L.; Silvestre, A.; Giraudo, L.; Veran, J.; Sabatier, F. Depa classification: A proposal for standardising prp use and a retrospective application of available devices. BMJ Open Sport Exerc Med. 2016, 2, e000060. [Google Scholar] [CrossRef] [Green Version]
- Everts, P.A.; van Zundert, A.; Schonberger, J.P.; Devilee, R.J.; Knape, J.T. What do we use: Platelet-rich plasma or platelet-leukocyte gel? J. Biomed. Mater. Res. A 2008, 85, 1135–1136. [Google Scholar] [CrossRef]
- Aghababazadeh, M.; Kerachian, M.A. Cell fasting: Cellular response and application of serum starvation. J. Nutr. Fasting Health 2014, 2, 147–150. [Google Scholar]
- Radiono, S.; Yohanes Widodo, W.; Budiyanto, A. The effect of prf on serum starved human dermal fibroblast. J. Med. Sci. 2016, 48, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Herrick, S.E.; Ireland, G.W.; Simon, D.; McCollum, C.N.; Ferguson, M.W. Venous ulcer fibroblasts compared with normal fibroblasts show differences in collagen but not fibronectin production under both normal and hypoxic conditions. J. Investig. Dermatol. 1996, 106, 187–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renjith, P.N.; Jasmin, J.; Harikrishnan, V.S.; Krishnan, V.K.; Lissy, K. Contribution of fibroblasts to the mechanical stability of in vitro engineered dermal-like tissue through extracellular matrix deposition. BioResearch Open Access 2014, 3, 217–225. [Google Scholar]
- Thangapazham, R.L.; Darling, T.N.; Meyerle, J. Alteration of skin properties with autologous dermal fibroblasts. Int. J. Mol. Sci. 2014, 15, 8407–8427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jesús, A.M.; Sander, E.A. Observing and quantifying fibroblast-mediated fibrin gel compaction. J. Vis. Exp. 2014, 50918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Z.; Balimunkwe, R.M.; Quan, T. Age-related reduction of dermal fibroblast size upregulates multiple matrix metalloproteinases as observed in aged human skin in vivo. Br. J. Derm. 2017, 177, 1337–1348. [Google Scholar] [CrossRef]
- Shin, M.K.; Lee, J.W.; Kim, Y.I.; Kim, Y.-O.; Seok, H.; Kim, N.I. The effects of platelet-rich clot releasate on the expression of mmp-1 and type I collagen in human adult dermal fibroblasts: Prp is a stronger mmp-1 stimulator. Mol. Biol. Rep. 2014, 41, 3–8. [Google Scholar] [CrossRef]
- Karamanos, N.K.; Theocharis, A.D.; Neill, T.; Iozzo, R.V. Matrix modeling and remodeling: A biological interplay regulating tissue homeostasis and diseases. Matrix Biol. 2019, 75–76, 1–11. [Google Scholar] [CrossRef]
- Caley, M.P.; Martins, V.L.C.; O’Toole, E.A. Metalloproteinases and wound healing. Adv. Wound Care 2015, 4, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.; Frampton, G.; Nelson, P.N. Cell adhesion molecules. Sticky moments in the clinic. BMJ 1999, 319, 332–334. [Google Scholar] [CrossRef]
- Schnittert, J.; Bansal, R.; Storm, G.; Prakash, J. Integrins in wound healing, fibrosis and tumor stroma: High potential targets for therapeutics and drug delivery. Adv. Drug Deliv. Rev. 2018, 129, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Vranka, J.A.; Bradley, J.M.; Yang, Y.-F.; Keller, K.E.; Acott, T.S. Mapping molecular differences and extracellular matrix gene expression in segmental outflow pathways of the human ocular trabecular meshwork. PLoS ONE 2015, 10, e0122483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamanos, N.K. Extracellular Matrix: Pathobiology and Signaling; Walter de Gruyter GmbH & Co.: Berlin, Germany, 2012. [Google Scholar]
- Adams, J.C.; Watt, F.M. Regulation of development and differentiation by the extracellular matrix. Development 1993, 117, 1183–1198. [Google Scholar] [PubMed]
- Martinez, F.O.; Gordon, S. The m1 and m2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokarram, N.; Merchant, A.; Mukhatyar, V.; Patel, G.; Bellamkonda, R.V. Effect of modulating macrophage phenotype on peripheral nerve repair. Biomaterials 2012, 33, 8793–8801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiller, K.L.; Koh, T.J. Macrophage-based therapeutic strategies in regenerative medicine. Adv. Drug Deliv. Rev. 2017, 122, 74–83. [Google Scholar] [CrossRef]
- Ogle, M.E.; Segar, C.E.; Sridhar, S.; Botchwey, E.A. Monocytes and macrophages in tissue repair: Implications for immunoregenerative biomaterial design. Exp. Biol. Med. (Maywoodn. J.) 2016, 241, 1084–1097. [Google Scholar] [CrossRef]
- Giacco, F.; Perruolo, G.; D’Agostino, E.; Fratellanza, G.; Perna, E.; Misso, S.; Saldalamacchia, G.; Oriente, F.; Fiory, F.; Miele, C.; et al. Thrombin-activated platelets induce proliferation of human skin fibroblasts by stimulating autocrine production of insulin-like growth factor-1. FASEB J. 2006, 20, 2402–2404. [Google Scholar] [CrossRef]
- Everts, P.A.; Hoffmann, J.; Weibrich, G.; Mahoney, C.B.; Schonberger, J.P.; van Zundert, A.; Knape, J.T. Differences in platelet growth factor release and leucocyte kinetics during autologous platelet gel formation. Transfus Med. 2006, 16, 363–368. [Google Scholar] [CrossRef]
- Mizoguchi, T.; Ueno, K.; Takeuchi, Y.; Samura, M.; Suzuki, R.; Morikage, N.; Hamano, K.; Murata, T.; Hosoyama, T. Treatment of cutaneous ulcers with multilayered mixed sheets of autologous fibroblasts and peripheral blood mononuclear cells. Cell. Physiol. Biochem. 2018, 47, 201–211. [Google Scholar] [CrossRef]
- Devereaux, J.; Nurgali, K.; Kiatos, D.; Sakkal, S.; Apostolopoulos, V. Effects of platelet-rich plasma and platelet-poor plasma on human dermal fibroblasts. Maturitas 2018, 117, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Magalon, J.; Bausset, O.; Serratrice, N.; Giraudo, L.; Aboudou, H.; Veran, J.; Magalon, G.; Dignat-Georges, F.; Sabatier, F. Characterization and comparison of 5 platelet-rich plasma preparations in a single-donor model. Arthroscopy 2014, 30, 629–638. [Google Scholar] [CrossRef]
- Braschi, B.D.P.; Gray, K.; Jones, T.; Seal, R.; Tweedie, S.; Yates, B.; Bruford, E. Genenames. org: The HGNC and VGNC resources in 2019. Nucleic Acids Res. 2019, 47, D786–D792. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.P.; Patil, V.M. Matrix metalloproteinase inhibitors. Experientia Supplementum 2012, 103, 35–56. [Google Scholar]
- Lustosa, S.A.S.; de Souza Viana, L.; Affonso, R.J.; Silva, S.R.M.; Denadai, M.V.A.; de Toledo, S.R.C.; Oliveira, I.D.; Matos, D. Expression profiling using a cdna array and immunohistochemistry for the extracellular matrix genes fn-1, itga-3, itgb-5, mmp-2, and mmp-9 in colorectal carcinoma progression and dissemination. Sci. World J. 2014, 2014, 102541. [Google Scholar] [CrossRef]
- Mohan, V.T.-F.D.; Arkadash, V.; Papo, N.; Sagi, I. Matrix metalloproteinase protein inhibitors: Highlighting a new beginning for metalloproteinases in medicine. Met. Medicine. 2016, 3, 31–47. [Google Scholar]
- Mautner, K.; Malanga, G.A.; Smith, J.; Shiple, B.; Ibrahim, V.; Sampson, S.; Bowen, J.E. A call for a standard classification system for future biologic research: The rationale for new prp nomenclature. PM&R 2015, 7, S53–S59. [Google Scholar]
- Giordano, S.; Romeo, M.; di Summa, P.; Salval, A.; Lankinen, P. A meta-analysis on evidence of platelet-rich plasma for androgenetic alopecia. Int. J. Trichology 2018, 10, 1–10. [Google Scholar] [CrossRef]
- Frautschi, R.S.; Hashem, A.M.; Halasa, B.; Cakmakoglu, C.; Zins, J.E. Current evidence for clinical efficacy of platelet rich plasma in aesthetic surgery: A systematic review. Aesthetic Surg. J. 2017, 37, 353–362. [Google Scholar] [CrossRef]
- Gupta, A.K.; Carviel, J.L. Meta-analysis of efficacy of platelet-rich plasma therapy for androgenetic alopecia. J. Dermatol. Treat. 2017, 28, 55–58. [Google Scholar] [CrossRef]
- DeLong, J.M.; Russell, R.P.; Mazzocca, A.D. Platelet-rich plasma: The paw classification system. Arthroscopy 2012, 28, 998–1009. [Google Scholar] [CrossRef]
- Dohan Ehrenfest, D.M.; Rasmusson, L.; Albrektsson, T. Classification of platelet concentrates: From pure platelet-rich plasma (p-prp) to leucocyte- and platelet-rich fibrin (l-prf). Trends Biotechnol. 2009, 27, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Braun, H.J.; Kim, H.J.; Chu, C.R.; Dragoo, J.L. The effect of platelet-rich plasma formulations and blood products on human synoviocytes: Implications for intra-articular injury and therapy. Am. J. Sports Med. 2014, 42, 1204–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cross, J.A.; Cole, B.J.; Spatny, K.P.; Sundman, E.; Romeo, A.A.; Nicholson, G.P.; Wagner, B.; Fortier, L.A. Leukocyte-reduced platelet-rich plasma normalizes matrix metabolism in torn human rotator cuff tendons. Am. J. Sports Med. 2015, 43, 2898–2906. [Google Scholar] [CrossRef] [PubMed]
- Cieslik-Bielecka, A.; Glik, J.; Skowronski, R.; Bielecki, T. Benefit of leukocyte- and platelet-rich plasma in operative wound closure in oral and maxillofacial surgery. Biomed. Res. Int. 2016, 2016, 7649206. [Google Scholar] [CrossRef]
- Dragoo, J.L.; Braun, H.J.; Durham, J.L.; Ridley, B.A.; Odegaard, J.I.; Luong, R.; Arnoczky, S.P. Comparison of the acute inflammatory response of two commercial platelet-rich plasma systems in healthy rabbit tendons. Am. J. Sports Med. 2012, 40, 1274–1281. [Google Scholar] [CrossRef]
- Dohan Ehrenfest, D.M.; Bielecki, T.; Jimbo, R.; Barbé, G.; Del Corso, M.; Inchingolo, F.; Sammartino, G. Do the fibrin architecture and leukocyte content influence the growth factor release of platelet concentrates? An evidence-based answer comparing a pure platelet-rich plasma (p-prp) gel and a leukocyte- and platelet-rich fibrin (l-prf). Curr. Pharm. Biotechnol. 2012, 13, 1145–1152. [Google Scholar] [CrossRef]
- Noh, K.C.; Liu, X.N.; Zhuan, Z.; Yang, C.J.; Kim, Y.T.; Lee, G.W.; Choi, K.H.; Kim, K.O. Leukocyte-poor platelet-rich plasma-derived growth factors enhance human fibroblast proliferation in vitro. Clin. Orthop. Surg. 2018, 10, 240–247. [Google Scholar] [CrossRef]
- Xu, Z.; Yin, W.; Zhang, Y.; Qi, X.; Chen, Y.; Xie, X.; Zhang, C. Comparative evaluation of leukocyte- and platelet-rich plasma and pure platelet-rich plasma for cartilage regeneration. Sci. Rep. 2017, 7, 43301. [Google Scholar] [CrossRef]
- Van Linthout, S.; Miteva, K.; Tschöpe, C. Crosstalk between fibroblasts and inflammatory cells. Cardiovasc. Res. 2014, 102, 258–269. [Google Scholar] [CrossRef] [Green Version]
- Willenborg, S.; Eming, S.A. Macrophages—Sensors and effectors coordinating skin damage and repair. J. Dtsch. Derm. Ges. 2014, 12, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Andia, I.; Rubio-Azpeitia, E.; Martin, J.I.; Abate, M. Current concepts and translational uses of platelet rich plasma biotechnology. In Biotechnology Deniz Ekinci, IntechOpen; IntechOpen Limited: London, UK, 2015; pp. 1–31. [Google Scholar]
- Zhao, Y.; Glesne, D.; Huberman, E. A human peripheral blood monocyte-derived subset acts as pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2003, 100, 2426–2431. [Google Scholar] [CrossRef] [Green Version]
- Medina, A.; Brown, E.; Carr, N.; Ghahary, A. Circulating monocytes have the capacity to be transdifferentiated into keratinocyte-like cells. Wound Repair Regen 2009, 17, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Moojen, D.J.; Everts, P.A.; Schure, R.M.; Overdevest, E.P.; van Zundert, A.; Knape, J.T.; Castelein, R.M.; Creemers, L.B.; Dhert, W.J. Antimicrobial activity of platelet-leukocyte gel against staphylococcus aureus. J. Orthop Res. 2008, 26, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Stone, R.C.; Stojadinovic, O.; Ramirez, H.; Pastar, I.; Maione, A.G.; Smith, A.; Yanez, V.; Veves, A.; Kirsner, R.S.; et al. Integrative analysis of mirna and mrna paired expression profiling of primary fibroblast derived from diabetic foot ulcers reveals multiple impaired cellular functions. Wound Repair Regen. 2016, 24, 943–953. [Google Scholar] [CrossRef]
- Giusti, I.; Rughetti, A.; D’Ascenzo, S.; Di Stefano, G.; Nanni, M.R.; Millimaggi, D.; Dell’orso, L.; Dolo, V. The effects of platelet gel-released supernatant on human fibroblasts. Wound Repair Regen. 2013, 21, 300–308. [Google Scholar] [CrossRef]
- Nguyen, P.A.; Pham, T.A.V. Effects of platelet-rich plasma on human gingival fibroblast proliferation and migration in vitro. J. Appl. Oral Sci. 2018, 26, e20180077. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Je, Y.J.; Kim, C.D.; Lee, Y.H.; Seo, Y.J.; Lee, J.H.; Lee, Y. Can platelet-rich plasma be used for skin rejuvenation? Evaluation of effects of platelet-rich plasma on human dermal fibroblast. Ann. Derm. 2011, 23, 424–431. [Google Scholar] [CrossRef] [Green Version]
- Cho, E.B.; Park, G.S.; Park, S.S.; Jang, Y.J.; Kim, K.H.; Kim, K.J.; Park, E.J. Effect of platelet-rich plasma on proliferation and migration in human dermal fibroblasts. J. Cosmet. Dermatol. 2019, 18, 1105–1112. [Google Scholar] [CrossRef]
- Oostendorp, C.; Meyer, S.; Sobrio, M.; van Arendonk, J.; Reichmann, E.; Daamen, W.F.; van Kuppevelt, T.H. Evaluation of cultured human dermal- and dermo-epidermal substitutes focusing on extracellular matrix components: Comparison of protein and rna analysis. Burns 2017, 43, 520–530. [Google Scholar] [CrossRef]
- Pifer, M.A.; Maerz, T.; Baker, K.C.; Anderson, K. Matrix metalloproteinase content and activity in low-platelet, low-leukocyte and high-platelet, high-leukocyte platelet rich plasma (prp) and the biologic response to prp by human ligament fibroblasts. Am. J. Sports Med. 2014, 42, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Bellayr, I.; Pan, H.; Choi, Y.; Li, Y. Regeneration of soft tissues is promoted by mmp1 treatment after digit amputation in mice. PLoS ONE 2013, 8, e59105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, T.; Zhang, H.; Cetin, N.; Miller, E.; Moak, T.; Suen, J.Y.; Richter, G.T. Elevated expression of matrix metalloproteinase-9 not matrix metalloproteinase-2 contributes to progression of extracranial arteriovenous malformation. Sci. Rep. 2016, 6, 24378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.E. Mmp-12, an old enzyme plays a new role in the pathogenesis of rheumatoid arthritis? Am. J. Pathol. 2004, 165, 1069–1070. [Google Scholar] [CrossRef] [Green Version]
- Cipriani, V.; Ranzato, E.; Balbo, V.; Mazzucco, L.; Cavaletto, M.; Patrone, M. Long-term effect of platelet lysate on primary fibroblasts highlighted with a proteomic approach. J. Tissue Eng. Regen. Med. 2009, 3, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Caceres, M.; Martinez, C.; Martinez, J.; Smith, P.C. Effects of platelet-rich and -poor plasma on the reparative response of gingival fibroblasts. Clin. Oral Implant. Res. 2012, 23, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Giancotti, F.G.; Ruoslahti, E. Integrin signaling. Science 1999, 285, 1028. [Google Scholar] [CrossRef]
- Kenny, F.N.; Connelly, J.T. Integrin-mediated adhesion and mechano-sensing in cutaneous wound healing. Cell Tissue Res. 2015, 360, 571–582. [Google Scholar] [CrossRef]
- Koivisto, L.; Heino, J.; Häkkinen, L.; Larjava, H. Integrins in wound healing. Adv. Wound Care 2014, 3, 762–783. [Google Scholar] [CrossRef] [Green Version]
- Mahmoudian-Sani, M.R.; Rafeei, F.; Amini, R.; Saidijam, M. The effect of mesenchymal stem cells combined with platelet-rich plasma on skin wound healing. J. Cosmet. Dermatol. 2018, 17, 650–659. [Google Scholar] [CrossRef]
- Liu, S.Y.; Ge, D.; Chen, L.N.; Zhao, J.; Su, L.; Zhang, S.L.; Miao, J.Y.; Zhao, B.X. A small molecule induces integrin β4 nuclear translocation and apoptosis selectively in cancer cells with high expression of integrin β4. Oncotarget 2016, 7, 16282–16296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venter, N.G.; Marques, R.G.; Santos, J.S.d.; Monte-Alto-Costa, A. Use of platelet-rich plasma in deep second- and third-degree burns. Burns 2016, 42, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Boo, S.; Dagnino, L. Integrins as modulators of transforming growth factor beta signaling in dermal fibroblasts during skin regeneration after injury. Adv. Wound Care 2013, 2, 238–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranzato, E.; Martinotti, S.; Volante, A.; Mazzucco, L.; Burlando, B. Platelet lysate modulates mmp-2 and mmp-9 expression, matrix deposition and cell-to-matrix adhesion in keratinocytes and fibroblasts. Exp Derm. 2011, 20, 308–313. [Google Scholar] [CrossRef]
- Kong, D.-H.; Kim, Y.K.; Kim, M.R.; Jang, J.H.; Lee, S. Emerging roles of vascular cell adhesion molecule-1 (vcam-1) in immunological disorders and cancer. Int. J. Mol. Sci. 2018, 19, 1057. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.; Moon, S.O.; Kim, S.H.; Kim, H.J.; Koh, Y.S.; Koh, G.Y. Vascular endothelial growth factor expression of intercellular adhesion molecule 1 (icam-1), vascular cell adhesion molecule 1 (vcam-1), and e-selectin through nuclear factor-kappa b activation in endothelial cells. J. Biol Chem. 2001, 276, 7614–7620. [Google Scholar] [CrossRef] [Green Version]
- Hosking, B.M.; Wang, S.C.M.; Downes, M.; Koopman, P.; Muscat, G.E.O. The vcam-1 gene that encodes the vascular cell adhesion molecule is a target of the sry-related high mobility group box gene, sox18. J. Biol. Chem. 2004, 279, 5314–5322. [Google Scholar] [CrossRef] [Green Version]
- Francavilla, C.; Loeffler, S.; Fau-Piccini, D.; Piccini, D.; Fau-Kren, A.; Kren, A.; Fau-Christofori, G.; Christofori, G.; Fau-Cavallaro, U.; Cavallaro, U. Neural cell adhesion molecule regulates the cellular response to fibroblast growth factor. J. Cell Sci. 2007, 120, 4388–4394. [Google Scholar] [CrossRef] [Green Version]
- Chooi, W.H.; Chew, S.Y. Review: Modulation of cell-cell interactions for neural tissue engineering: Potential therapeutic applications of cell adhesion molecules in nerve regeneration. Biomaterials 2019, 197, 327–344. [Google Scholar] [CrossRef]
- Jørgensen, O.S. Neural cell adhesion molecule (ncam) as a quantitative marker in synaptic remodeling. Neurochem. Res. 1995, 20, 533–547. [Google Scholar] [CrossRef]
- Beljaards, R.C.; Kirtschig, G.; Boorsma, D.M. Expression of neural cell adhesion molecule (cd56) in basal and squamous cell carcinoma. Dermatol. Surg. 2008, 34, 1577–1579. [Google Scholar] [PubMed]
- Kurokawa, M.; Nabeshima, K.; Akiyama, Y.; Maeda, S.; Nishida, T.; Nakayama, F.; Amano, M.; Ogata, K.; Setoyama, M. Cd56: A useful marker for diagnosing merkel cell carcinoma. J. Dermatol. Sci. 2003, 31, 219–224. [Google Scholar] [CrossRef]
- Kakudo, N.; Minakata, T.; Mitsui, T.; Kushida, S.; Notodihardjo, F.Z.; Kusumoto, K. Proliferation-promoting effect of platelet-rich plasma on human adipose–derived stem cells and human dermal fibroblasts. Plast. Reconstr. Surg. 2008, 122, 1352–1360. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, K.; Prakash, M.; Kuol, N.; Nurgali, K.; Stojanovska, L.; Apostolopoulos, V. Anti-tumor effects of vitamin b2, b6 and b9 in promonocytic lymphoma cells. Int. J. Mol. Sci. 2019, 20, 3763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grada, A.; Otero-Vinas, M.; Prieto-Castrillo, F.; Obagi, Z.; Falanga, V. Research techniques made simple: Analysis of collective cell migration using the wound healing assay. J. Investig. Dermatol. 2017, 137, e11–e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimble, J.M.; Katz, A.J.; Bunnell, B.A. Adipose-derived stem cells for regenerative medicine. Circ. Res. 2007, 100, 1249–1260. [Google Scholar] [CrossRef]
- Dargahi, N.; Matsoukas, J.; Apostolopoulos, V. Streptococcusthermophilus st285 alters pro-inflammatory to anti-inflammatory cytokine secretion against multiple sclerosis peptide in mice. Brain Sci. 2020, 10, 126. [Google Scholar] [CrossRef] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Devereaux, J.; Dargahi, N.; Fraser, S.; Nurgali, K.; Kiatos, D.; Apostolopoulos, V. Leucocyte-Rich Platelet-Rich Plasma Enhances Fibroblast and Extracellular Matrix Activity: Implications in Wound Healing. Int. J. Mol. Sci. 2020, 21, 6519. https://doi.org/10.3390/ijms21186519
Devereaux J, Dargahi N, Fraser S, Nurgali K, Kiatos D, Apostolopoulos V. Leucocyte-Rich Platelet-Rich Plasma Enhances Fibroblast and Extracellular Matrix Activity: Implications in Wound Healing. International Journal of Molecular Sciences. 2020; 21(18):6519. https://doi.org/10.3390/ijms21186519
Chicago/Turabian StyleDevereaux, Jeannie, Narges Dargahi, Sarah Fraser, Kulmira Nurgali, Dimitrios Kiatos, and Vasso Apostolopoulos. 2020. "Leucocyte-Rich Platelet-Rich Plasma Enhances Fibroblast and Extracellular Matrix Activity: Implications in Wound Healing" International Journal of Molecular Sciences 21, no. 18: 6519. https://doi.org/10.3390/ijms21186519
APA StyleDevereaux, J., Dargahi, N., Fraser, S., Nurgali, K., Kiatos, D., & Apostolopoulos, V. (2020). Leucocyte-Rich Platelet-Rich Plasma Enhances Fibroblast and Extracellular Matrix Activity: Implications in Wound Healing. International Journal of Molecular Sciences, 21(18), 6519. https://doi.org/10.3390/ijms21186519