A Peripheral CB1R Antagonist Increases Lipolysis, Oxygen Consumption Rate, and Markers of Beiging in 3T3-L1 Adipocytes Similar to RIM, Suggesting that Central Effects Can Be Avoided

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
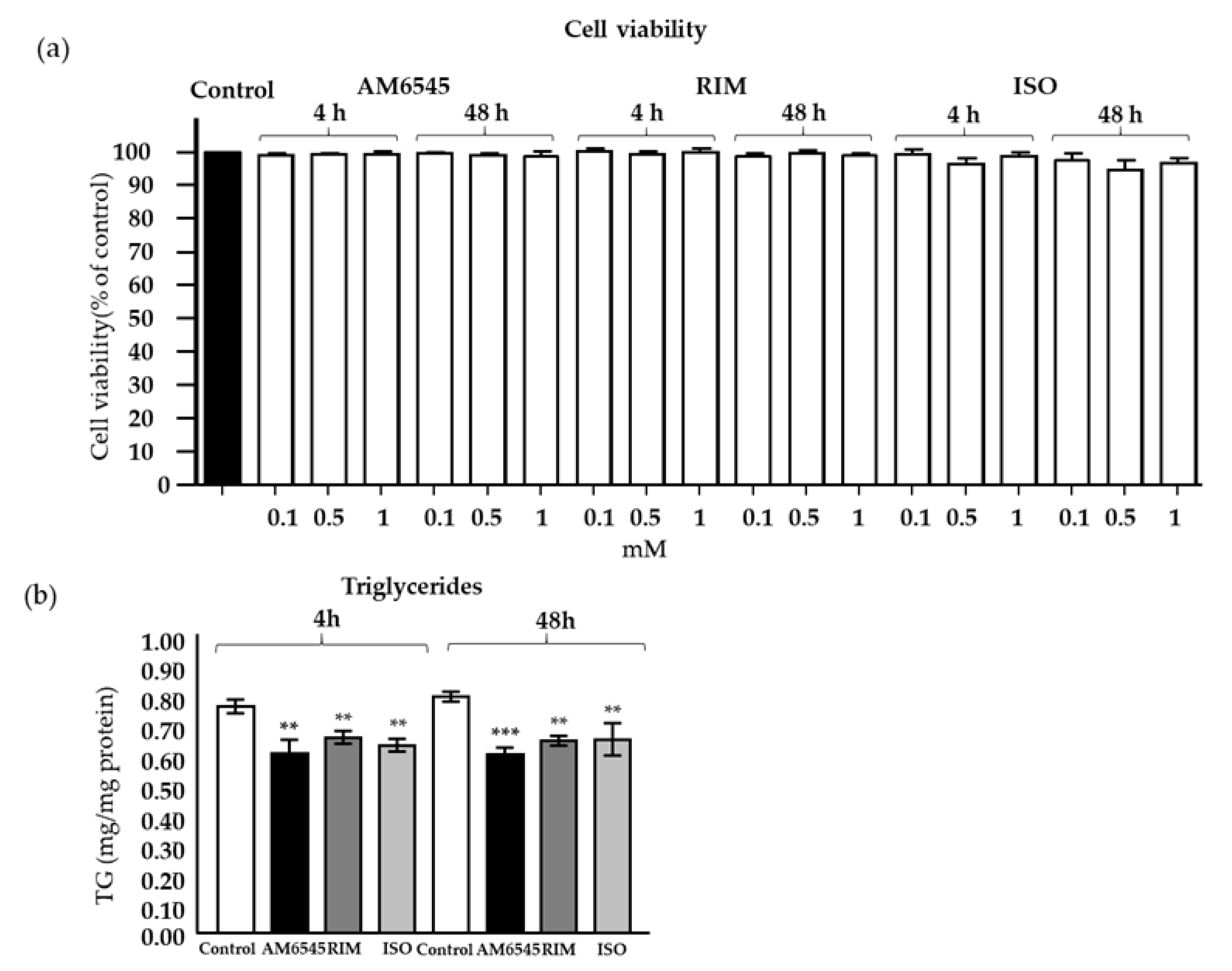
2.1. Treatment with CB1R Antagonists Does Not Affect Cell Viability
2.2. Treatment with CB1R Antagonists Decreased Triglyceride Content
2.3. The Peripheral CB1R Antagonist-Induced Genes Involved in Beiging in 3T3-L1 Mature Adipocytes
2.4. Lipolysis Increased by the Peripheral CB1R Antagonist
2.5. The Peripheral CB1 Antagonist Increased mtDNA and Mitochondrial Biogenesis Genes
2.6. The Peripheral CB1R Antagonist Increased Mitochondrial Respiration and Proton Leak in 3T3-L1 Adipocytes
2.7. CB1R Antagonism-Induced Improvements in Mitochondrial Function Rely on Lipolysis, as Shown During Real-Time OCR
3. Discussion
4. Materials and Methods
4.1. Preparation and Treatment of 3T3-L1 Adipocytes
4.2. Cell Viability and Triglyceride Measurements
4.3. Lipolysis Assays
4.4. Mitochondrial DNA Copy Number
4.5. Measurement of Oxygen Consumption Rate
4.6. Total RNA Isolation and Real Time PCR (RT-PCR)
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CB1R | Cannabinoid receptor 1 |
| RIM | Rimonabant |
| ISO | Isoproterenol |
| OCR | Oxygen consumption rate |
| T2D | Type 2 diabetes |
| WAT | White adipose tissue |
| BAT | Brown adipose tissue |
| ECS | Endocannabinoid system |
| β3 | Beta-3 adrenergic |
| NP | Natriuretic peptide |
| Pgc1α | Peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| Prdm16 | PR domain containing 16 |
| Ucp1 | Uncoupling protein 1 |
| Cidea | Cell death-inducing DFFA-Like effector A |
| Elovl3 | elongation of very long chain fatty Acids protein 3 |
| Cd137 | tumor necrosis factor receptor superfamily member 9 |
| Tbx1 | T-box Transcription factor |
| Tmem26 | Transmembrane protein 26 |
| Cited1 | Cbp/p300-interacting transactivator 1 |
| Serca2b | Sarco/endoplasmic reticulum Ca2+-ATPase2b |
| RyR2 | Ryanodine receptor 2 |
| ATGL | Adipose triglyceride lipase |
| HSL | Hormone sensitive lipase |
| FFCP | Carbonyl cyanide-4(trifluoromethoxy) phenylhydrazone |
| Tfam | transcription factor A, mitochondria |
| Nrf1 | nuclear respiratory factor 1 |
| Nd1 | dinucleotide dehydrogenase 1 |
| Actb | mouse β-actin |
References
- Grundy, S.M. Metabolic syndrome update. Trends Cardiovasc. Med. 2016, 26, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Gruzdeva, O.V.; Borodkina, D.; Uchasova, E.; Dyleva, Y.; Barbarash, O.L. Localization of fat depots and cardiovascular risk. Lipids Heal. Dis. 2018, 17, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kachur, S.; Lavie, C.J.; de Schutter, A.; Milani, R.V.; Ventura, H.O. Obesity and cardiovascular diseases. Minerva Med. 2017, 108, 212–228. [Google Scholar] [PubMed]
- Harms, M.; Seale, P. Brown and beige fat: Development, function and therapeutic potential. Nat. Med. 2013, 19, 1252–1263. [Google Scholar] [CrossRef] [Green Version]
- Calvani, R.; Leeuwenburgh, C.; Marzetti, E. Brown adipose tissue and the cold war against obesity. Diabetes 2014, 63, 3998–4000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueta, C.B.; Fernandes, G.W.; Capelo, L.P.; Fonseca, T.L.; Maculan, F.D.; Gouveia, C.H.; Brum, P.C.; Christoffolete, M.A.; Aoki, M.S.; Lancellotti, C.L.; et al. beta(1) Adrenergic receptor is key to cold- and diet-induced thermogenesis in mice. J. Endocrinol. 2012, 214, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Blüher, M.; Wilson-Fritch, L.; Leszyk, J.; Laustsen, P.G.; Corvera, S.; Kahn, C.R. Role of Insulin Action and Cell Size on Protein Expression Patterns in Adipocytes. J. Boil. Chem. 2004, 279, 31902–31909. [Google Scholar] [CrossRef] [Green Version]
- Zamora-Mendoza, R.; Rosas-Vargas, H.; Ramos-Cervantes, M.T.; Garcia-Zuniga, P.; Perez-Lorenzana, H.; Mendoza-Lorenzo, P.; Perez-Ortiz, A.C.; Estrada-Mena, F.J.; Miliar-Garcia, A.; Lara-Padilla, E.; et al. Dysregulation of mitochondrial function and biogenesis modulators in adipose tissue of obese children. Int. J. Obes. 2017, 42, 618–624. [Google Scholar] [CrossRef]
- Yin, X.; Lanza, I.R.; Swain, J.M.; Sarr, M.G.; Nair, K.S.; Jensen, M.D. Adipocyte mitochondrial function is reduced in human obesity independent of fat cell size. J. Clin. Endocrinol. Metab. 2013, 99, E209–E216. [Google Scholar] [CrossRef] [Green Version]
- Mazier, W.; Saucisse, N.; Gatta-Cherifi, B.; Cota, D. The Endocannabinoid System: Pivotal Orchestrator of Obesity and Metabolic Disease. Trends Endocrinol. Metab. 2015, 26, 524–537. [Google Scholar] [CrossRef]
- Ravinet, T.C.; Arnone, M.; Delgorge, C.; Gonalons, N.; Keane, P.; Maffrand, J.P.; Soubrie, P. Anti-obesity effect of SR141716, a CB1 receptor antagonist, in diet-induced obese mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R345–R353. [Google Scholar] [CrossRef] [Green Version]
- Gary-Bobo, M.; ElAchouri, G.; Gallas, J.F.; Janiak, P.; Marini, P.; Ravinet-Trillou, C.; Chabbert, M.; Cruccioli, N.; Pfersdorff, C.; Roque, C.; et al. Rimonabant reduces obesity-associated hepatic steatosis and features of metabolic syndrome in obese Zucker fa/fa rats. Hepatolology 2007, 46, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Richey, J.M.; Woolcott, O.O.; Stefanovski, D.; Harrison, L.N.; Zheng, D.; Lottati, M.; Hsu, I.R.; Kim, S.P.; Kabir, M.; Catalano, K.J.; et al. Rimonabant prevents additional accumulation of visceral and subcutaneous fat during high-fat feeding in dogs. Am. J. Physiol. Metab. 2009, 296, E1311–E1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabir, M.; Iyer, M.S.; Richey, J.M.; Woolcott, O.O.; Bediako, I.A.; Wu, Q.; Kim, S.P.; Stefanovski, D.; Kolka, C.M.; Hsu, I.R.; et al. CB1R antagonist increases hepatic insulin clearance in fat-fed dogs likely via upregulation of liver adiponectin receptors. Am. J. Physiol. Metab. 2015, 309, E747–E758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Gaal, L.F.; Rissanen, A.M.; Scheen, A.J.; Ziegler, O.; Rossner, S. Effects of the cannabinoid-1 receptor blocker rimonabant on weight reduction and cardiovascular risk factors in overweight patients: 1-year experience from the RIO-Europe study. Lancet 2005, 365, 1389–1397. [Google Scholar] [CrossRef]
- Kabir, M.; Stefanovski, D.; Hsu, I.R.; Iyer, M.S.; Woolcott, O.O.; Zheng, D.; Catalano, K.J.; Chiu, J.D.; Kim, S.P.; Harrison, L.N.; et al. Large size cells in the visceral adipose depot predict insulin resistance in the canine model. Obesity 2011, 19, 2121–2129. [Google Scholar] [CrossRef] [Green Version]
- Bajzer, M.; Olivieri, M.; Haas, M.K.; Pfluger, P.T.; Magrisso, I.J.; Foster, M.T.; Tschop, M.H.; Krawczewski-Carhuatanta, K.A.; Cota, D.; Obici, S. Cannabinoid receptor 1 (CB1) antagonism enhances glucose utilisation and activates brown adipose tissue in diet-induced obese mice. Diabetologia 2011, 54, 3121–3131. [Google Scholar] [CrossRef] [Green Version]
- Iyer, M.S.; Paszkiewicz, R.L.; Bergman, R.N.; Richey, J.M.; Woolcott, O.O.; Bediako, I.A.; Wu, Q.; Kim, S.P.; Stefanovski, D.; Kolka, C.M.; et al. Activation of NPRs and UCP1-independent pathway following CB1R antagonist treatment is associated with adipose tissue beiging in fat-fed male dogs. Am. J. Physiol. Metab. 2019, 317, E535–E547. [Google Scholar] [CrossRef]
- Lafontan, M.; Berlan, M. Fat cell adrenergic receptors and the control of white and brown fat cell function. J. Lipid Res. 1993, 34, 1057–1091. [Google Scholar]
- Louis, S.N.; Jackman, G.P.; Nero, T.L.; Iakovidis, D.; Louis, W.J. Role of beta-adrenergic receptor subtypes in lipolysis. Cardiovasc. Drugs Ther. 2000, 14, 565–577. [Google Scholar] [CrossRef]
- Sengenes, C.; Berlan, M.; de Glisezinski, I.; Lafontan, M.; Galitzky, J. Natriuretic peptides: A new lipolytic pathway in human adipocytes. FASEB J. 2000, 14, 1345–1351. [Google Scholar] [CrossRef] [PubMed]
- Jbilo, O.; Trillou, C.R.; Arnone, M.; Buisson, I.; Bribes, E.; Péleraux, A.; Pénarier, G.; Soubrié, P.; Le Fur, G.; Galiègue, S.; et al. The CB1 receptor antagonist rimonabant reverses the diet-induced obesity phenotype through the regulation of lipolysis and energy balance. FASEB J. 2005, 19, 1567–1569. [Google Scholar] [CrossRef] [PubMed]
- Moreira, F.A.; Crippa, J.A.S. The psychiatric side-effects of rimonabant. Rev. Bras. Psiquiatr. 2009, 31, 145–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perwitz, N.; Wenzel, J.; Wagner, I.; Büning, J.; Drenckhan, M.; Zarse, K.; Ristow, M.; Lilienthal, W.; Lehnert, H.; Klein, J. Cannabinoid type 1 receptor blockade induces transdifferentiation towards a brown fat phenotype in white adipocytes. Diabetes Obes. Metab. 2010, 12, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Boon, M.R.; Kooijman, S.; Van Dam, A.D.; Pelgrom, L.R.; Berbée, J.F.P.; Visseren, C.A.R.; Van Aggele, R.C.; Hoek, A.M.V.D.; Sips, H.C.M.; Lombès, M.; et al. Peripheral cannabinoid 1 receptor blockade activates brown adipose tissue and diminishes dyslipidemia and obesity. FASEB J. 2014, 28, 5361–5375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Kubota, N.; Ohsugi, M.; Kubota, T.; Takamoto, I.; Iwabu, M.; Awazawa, M.; Katsuyama, H.; Hasegawa, C.; Tokuyama, K.; et al. Rimonabant Ameliorates Insulin Resistance via both Adiponectin-dependent and Adiponectin-independent Pathways. J. Boil. Chem. 2008, 284, 1803–1812. [Google Scholar] [CrossRef] [Green Version]
- Müller, G.A.; Herling, A.W.; Wied, S.; Müller, T.D. CB1 Receptor-Dependent and Independent Induction of Lipolysis in Primary Rat Adipocytes by the Inverse Agonist Rimonabant (SR141716A). Moleculs 2020, 25, 896. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Zhang, G.; Mou, C.; Fu, X.; Chen, Y. Peripheral CB1 Receptor Neutral Antagonist, AM6545, Ameliorates Hypometabolic Obesity and Improves Adipokine Secretion in Monosodium Glutamate Induced Obese Mice. Front. Pharmacol. 2018, 9, 156. [Google Scholar] [CrossRef] [Green Version]
- Tam, J.; Cinar, R.; Liu, J.; Godlewski, G.; Wesley, D.; Jourdan, T.; Szanda, G.; Mukhopadhyay, B.; Chedester, L.; Liow, J.-S.; et al. Peripheral cannabinoid-1 receptor inverse agonism reduces obesity by reversing leptin resistance. Cell Metab. 2012, 16, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Yan, E.; Durazo, F.; Tong, M.; Hong, K. Nonalcoholic Fatty Liver Disease: Pathogenesis, Identification, Progression, and Management. Nutr. Rev. 2007, 65, 376–384. [Google Scholar] [CrossRef]
- Chen, Y.; Ikeda, K.; Yoneshiro, T.; Scaramozza, A.; Tajima, K.; Wang, Q.; Kim, K.; Shinoda, K.; Sponton, C.H.; Brown, Z.; et al. Thermal stress induces glycolytic beige fat formation via a myogenic state. Nature 2018, 565, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Kazak, L. UCP1-independent thermogenesis. Biochem. J. 2020, 477, 709–725. [Google Scholar] [CrossRef]
- Kazak, L.; Rahbani, J.F.; Samborska, B.; Lu, G.Z.; Jedrychowski, M.P.; Lajoie, M.; Zhang, S.; Ramsay, L.; Dou, F.Y.; Tenen, D.; et al. Ablation of adipocyte creatine transport impairs thermogenesis and causes diet-induced obesity. Nat. Metab. 2019, 1, 360–370. [Google Scholar] [CrossRef] [PubMed]
- MacPherson, R.E.K.; Gamu, D.; Frendo-Cumbo, S.; Castellani, L.; Kwon, F.; Tupling, A.R.; Wright, D.C. Sarcolipin knockout mice fed a high-fat diet exhibit altered indices of adipose tissue inflammation and remodeling. Obesity 2016, 24, 1499–1505. [Google Scholar] [CrossRef]
- Song, A.; Dai, W.; Jang, M.J.; Medrano, L.; Li, Z.; Zhao, H.; Shao, M.; Tan, J.; Li, A.; Ning, T.; et al. Low- and high-thermogenic brown adipocyte subpopulations coexist in murine adipose tissue. J. Clin. Investig. 2019, 130, 247–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartelt, A.; Bruns, O.T.; Reimer, R.; Hohenberg, H.; Ittrich, H.; Peldschus, K.; Kaul, M.G.; Tromsdorf, U.I.; Weller, H.; Waurisch, C.; et al. Brown adipose tissue activity controls triglyceride clearance. Nat. Med. 2011, 17, 200–205. [Google Scholar] [CrossRef]
- Seale, P.; Kajimura, S.; Spiegelman, B.M. Transcriptional control of brown adipocyte development and physiological function--of mice and men. Genes Dev. 2009, 23, 788–797. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, R.; Wang, F.; Hong, J.; Li, X.; Chen, M.; Ke, Y.; Zhang, X.; Ma, Q.; Wang, R.; et al. Ablation of LGR4 promotes energy expenditure by driving white-to-brown fat switch. Nat. Cell Biol. 2013, 15, 1455–1463. [Google Scholar] [CrossRef]
- Gómez-Ambrosi, J.; Frühbeck, G.; Martínez, J.A. Rapid in vivo PGC-1 mRNA upregulation in brown adipose tissue of Wistar rats by a beta(3)-adrenergic agonist and lack of effect of leptin. Mol. Cell. Endocrinol. 2001, 176, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Sharma, B.K.; Patil, M.; Satyanarayana, A. Negative Regulators of Brown Adipose Tissue (BAT)-Mediated Thermogenesis. J. Cell. Physiol. 2014, 229, 1901–1907. [Google Scholar] [CrossRef] [Green Version]
- Puigserver, P.; Spiegelman, B.M. Peroxisome Proliferator-Activated Receptor-γ Coactivator 1α (PGC-1α): Transcriptional Coactivator and Metabolic Regulator. Endocr. Rev. 2003, 24, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms Controlling Mitochondrial Biogenesis and Respiration through the Thermogenic Coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Gallardo-Montejano, V.I.; Saxena, G.; Kusminski, C.M.; Yang, C.; McAfee, J.; Hahner, L.; Hoch, K.; Dubinsky, W.; Narkar, V.A.; Bickel, P.E. Nuclear Perilipin 5 integrates lipid droplet lipolysis with PGC-1α/SIRT1-dependent transcriptional regulation of mitochondrial function. Nat. Commun. 2016, 7, 12723. [Google Scholar] [CrossRef] [PubMed]
- Bogacka, I.; Xie, H.; Bray, G.A.; Smith, S.R. Pioglitazone Induces Mitochondrial Biogenesis in Human Subcutaneous Adipose Tissue In Vivo. Diabetes 2005, 54, 1392–1399. [Google Scholar] [CrossRef] [Green Version]
- Wilson-Fritch, L.; Burkart, A.; Bell, G.; Mendelson, K.; Leszyk, J.; Nicoloro, S.; Czech, M.; Corvera, S. Mitochondrial Biogenesis and Remodeling during Adipogenesis and in Response to the Insulin Sensitizer Rosiglitazone. Mol. Cell. Biol. 2003, 23, 1085–1094. [Google Scholar] [CrossRef] [Green Version]
- Cluny, N.; Vemuri, V.; Chambers, A.; Limebeer, C.; Bedard, H.; Wood, J.; Lutz, B.; Zimmer, A.; Parker, L.; Makriyannis, A.; et al. A novel peripherally restricted cannabinoid receptor antagonist, AM6545, reduces food intake and body weight, but does not cause malaise, in rodents. Br. J. Pharmacol. 2010, 161, 629–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, J.; Szanda, G.; Drori, A.; Liu, Z.; Cinar, R.; Kashiwaya, Y.; Reitman, M.L.; Kunos, G. Peripheral cannabinoid-1 receptor blockade restores hypothalamic leptin signaling. Mol. Metab. 2017, 6, 1113–1125. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.N.; Yang, J.-Y.; England, E.; Yin, A.; Baile, C.A.; Rayalam, S. Isoproterenol Increases Uncoupling, Glycolysis, and Markers of Beiging in Mature 3T3-L1 Adipocytes. PLoS ONE 2015, 10, e0138344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.-S.; Shin, Y.; Jung, S.; Kim, S.-Y.; Jo, Y.-H.; Kim, C.-T.; Yun, M.-K.; Lee, S.-J.; Sohn, J.; Yu, H.-J.; et al. The Inhibitory Effect of Tartary Buckwheat Extracts on Adipogenesis and Inflammatory Response. Molecules 2017, 22, 1160. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-H.; Wu, S.-B.; Hong, C.-H.; Liao, W.-T.; Wu, C.Y.; Chen, G.-S.; Wei, Y.-H.; Yu, H.-S. Aberrant Cell Proliferation by Enhanced Mitochondrial Biogenesis via mtTFA in Arsenical Skin Cancers. Am. J. Pathol. 2011, 178, 2066–2076. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.D.S.; Frank, A.P.; Fátima, L.A.; Palmer, B.F.; Öz, O.K.; Clegg, D.J. Activation of estrogen receptor alpha induces beiging of adipocytes. Mol. Metab. 2018, 18, 51–59. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PGC1a | Mm01,208,835_m1 |
| Prdm16 | Mm00,712,556_m1 |
| Ucp1 | Mm01,244,861_m1 |
| Cidea | Mm00,432,554_m1 |
| Elovl3 | Mm00,468,164_m1 |
| Tbx1 | Mm00,448,949_m1 |
| Cd137 | Mm00,441,899_m1 |
| Tmem26 | Mm01,173,641_m1 |
| Cited1 | Mm01,235,642_g1 |
| Serca2b | Mm01,201,431_m1 |
| Ryr2 | Mm00,465,877_m1 |
| ATGL | Mm00,503,040_m1 |
| HSL | Mm00,495,359_m1 |
| b3R | Mm02,601,819_g1 |
| Tfam | Mm00,447,485_m1 |
| Nrf | Mm01,135,607_m1 |
| Nd1 | Mm04,225,274_s1 |
| Actb | Mm02,619,580_g1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paszkiewicz, R.L.; Bergman, R.N.; Santos, R.S.; Frank, A.P.; Woolcott, O.O.; Iyer, M.S.; Stefanovski, D.; Clegg, D.J.; Kabir, M. A Peripheral CB1R Antagonist Increases Lipolysis, Oxygen Consumption Rate, and Markers of Beiging in 3T3-L1 Adipocytes Similar to RIM, Suggesting that Central Effects Can Be Avoided. Int. J. Mol. Sci. 2020, 21, 6639. https://doi.org/10.3390/ijms21186639
Paszkiewicz RL, Bergman RN, Santos RS, Frank AP, Woolcott OO, Iyer MS, Stefanovski D, Clegg DJ, Kabir M. A Peripheral CB1R Antagonist Increases Lipolysis, Oxygen Consumption Rate, and Markers of Beiging in 3T3-L1 Adipocytes Similar to RIM, Suggesting that Central Effects Can Be Avoided. International Journal of Molecular Sciences. 2020; 21(18):6639. https://doi.org/10.3390/ijms21186639
Chicago/Turabian StylePaszkiewicz, Rebecca L., Richard N. Bergman, Roberta S. Santos, Aaron P. Frank, Orison O. Woolcott, Malini S. Iyer, Darko Stefanovski, Deborah J. Clegg, and Morvarid Kabir. 2020. "A Peripheral CB1R Antagonist Increases Lipolysis, Oxygen Consumption Rate, and Markers of Beiging in 3T3-L1 Adipocytes Similar to RIM, Suggesting that Central Effects Can Be Avoided" International Journal of Molecular Sciences 21, no. 18: 6639. https://doi.org/10.3390/ijms21186639