Blocking ADAM17 Function with a Monoclonal Antibody Improves Sepsis Survival in a Murine Model of Polymicrobial Sepsis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
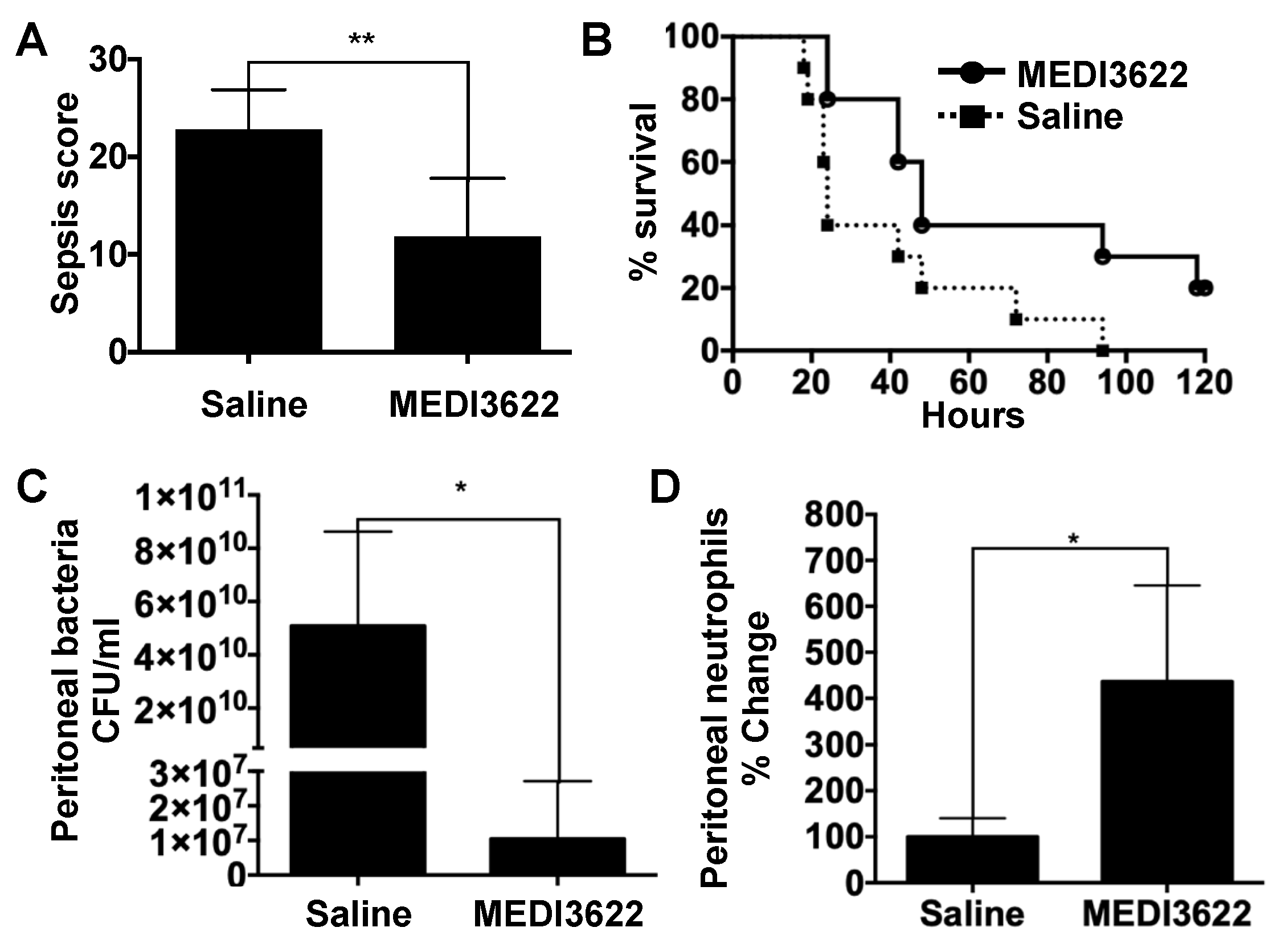
2.1. Administration of a mAb That Blocks ADAM17 Function Increases Sepsis Survival
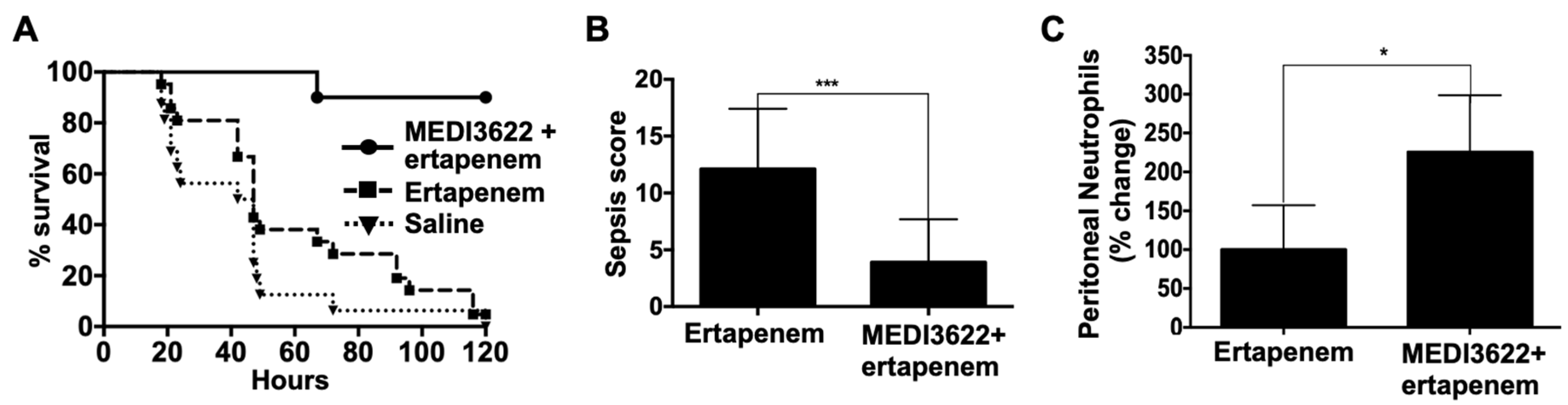
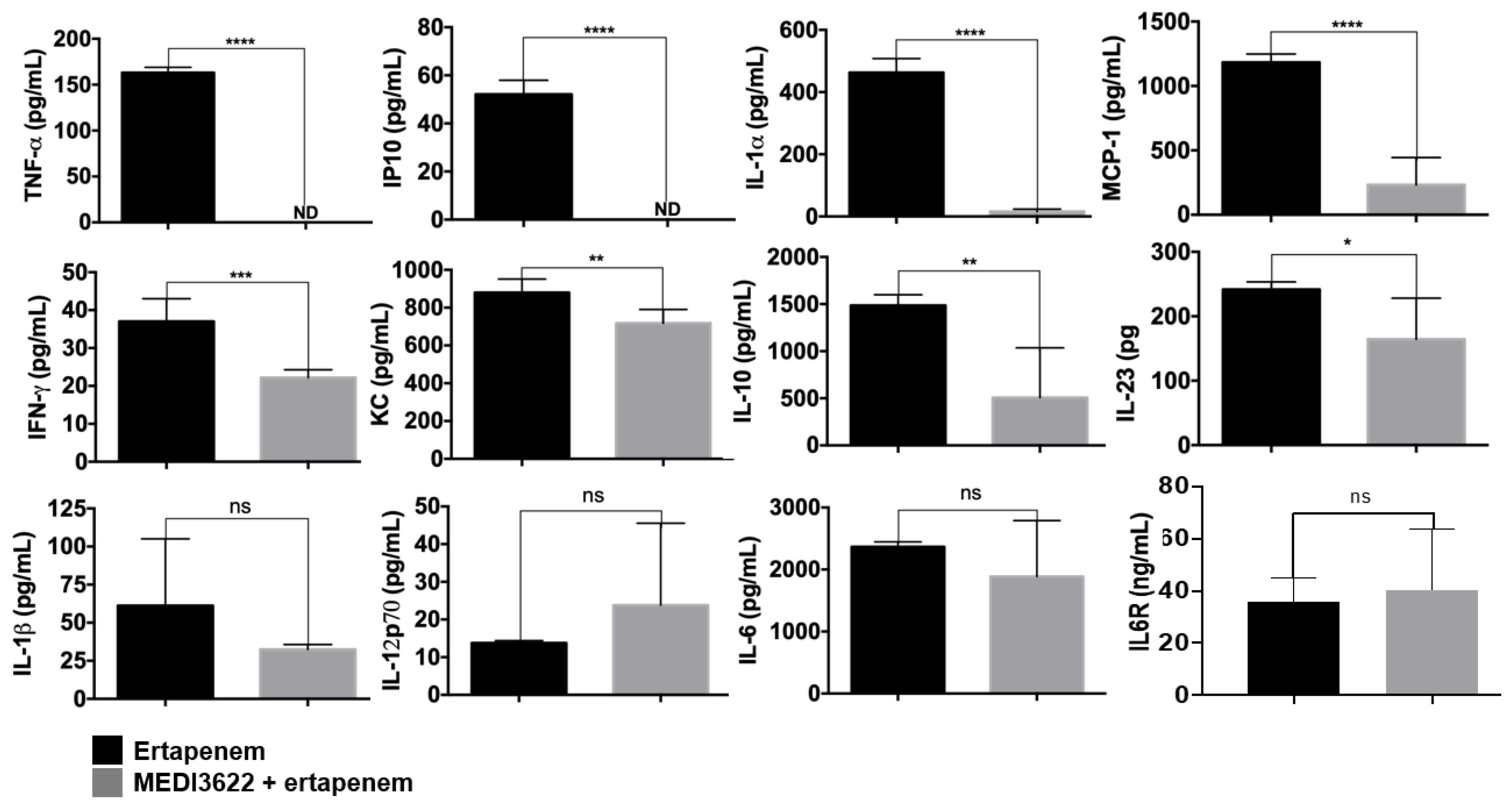
2.2. MEDI3622 in Combination with Antibiotic Treatment Further Enhances Sepsis Resistance
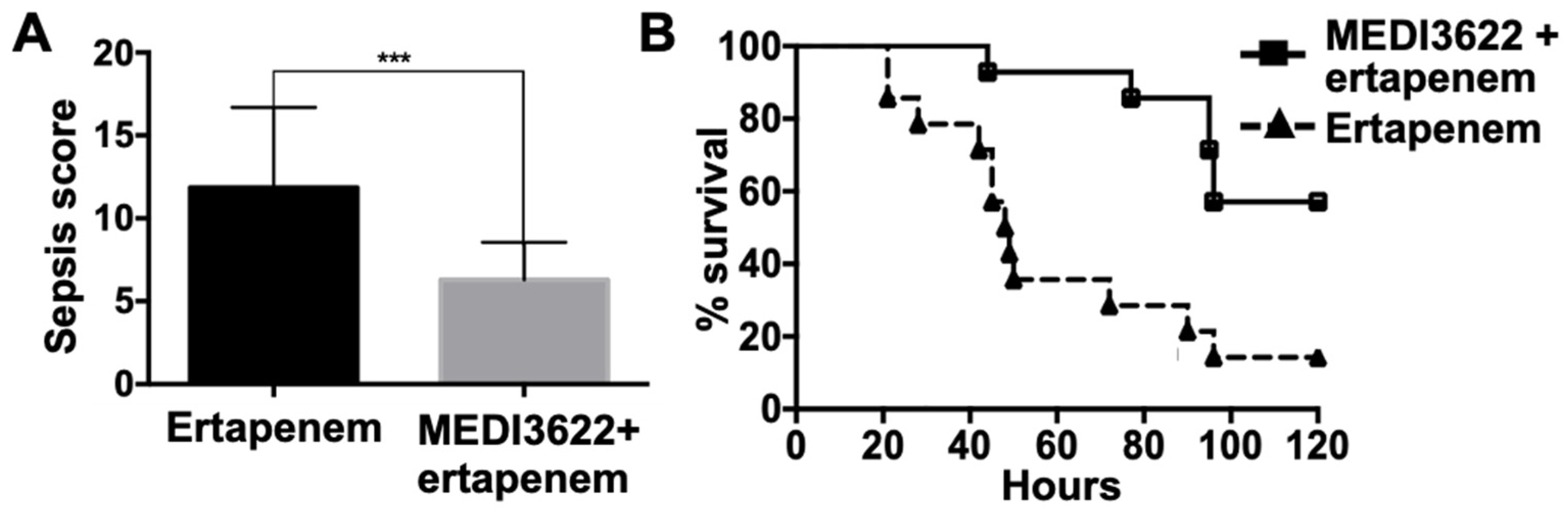
2.3. MEDI3622 Administration Post-CLP Prolongs Survival
3. Materials and Methods
3.1. Animals
3.2. Administration of MEDI3622 and Antibiotics
3.3. Quantification of Plasma Analytes
3.4. Enumeration of Blood and Peritoneal Fluid Levels of Neutrophils and Bacteria
3.5. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Delano, M.J.; Ward, P.A. The immune system’s role in sepsis progression, resolution, and long-term outcome. Immunol. Rev. 2016, 274, 330–353. [Google Scholar] [CrossRef]
- Paoli, C.J.; Reynolds, M.A.; Sinha, M.; Gitlin, M.; Crouser, E. Epidemiology and Costs of Sepsis in the United States-An Analysis Based on Timing of Diagnosis and Severity Level. Crit. Care Med. 2018, 46, 1889–1897. [Google Scholar] [CrossRef]
- Tiru, B.; DiNino, E.K.; Orenstein, A.; Mailloux, P.T.; Pesaturo, A.; Gupta, A.; McGee, W.T. The Economic and Humanistic Burden of Severe Sepsis. Pharmacoeconomics 2015, 33, 925–937. [Google Scholar] [CrossRef]
- Lammermann, T. In the eye of the neutrophil swarm-navigation signals that bring neutrophils together in inflamed and infected tissues. J. Leukoc. Biol. 2016, 100, 55–63. [Google Scholar] [CrossRef]
- Mayadas, T.N.; Cullere, X.; Lowell, C.A. The multifaceted functions of neutrophils. Annu. Rev. Pathol. 2014, 9, 181–218. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Karlin, A.; Loike, J.D.; Silverstein, S.C. A critical concentration of neutrophils is required for effective bacterial killing in suspension. Proc. Natl. Acad. Sci. USA 2002, 99, 8289–8294. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Karlin, A.; Loike, J.D.; Silverstein, S.C. Determination of the critical concentration of neutrophils required to block bacterial growth in tissues. J. Exp. Med. 2004, 200, 613–622. [Google Scholar] [CrossRef]
- Sonego, F.; Castanheira, F.V.; Ferreira, R.G.; Kanashiro, A.; Leite, C.A.; Nascimento, D.C.; Colon, D.F.; Borges Vde, F.; Alves-Filho, J.C.; Cunha, F.Q. Paradoxical Roles of the Neutrophil in Sepsis: Protective and Deleterious. Front. Immunol. 2016, 7, 155. [Google Scholar] [CrossRef] [Green Version]
- Mishra, H.K.; Ma, J.; Walcheck, B. Ectodomain Shedding by ADAM17: Its Role in Neutrophil Recruitment and the Impairment of This Process during Sepsis. Front. Cell Infect. Microbiol. 2017, 7, 138. [Google Scholar] [CrossRef]
- Zunke, F.; Rose-John, S. The shedding protease ADAM17: Physiology and pathophysiology. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864 Pt B, 2059–2070. [Google Scholar] [CrossRef]
- Ivetic, A.; Hoskins Green, H.L.; Hart, S.J. L-selectin: A Major Regulator of Leukocyte Adhesion, Migration and Signaling. Front. Immunol. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, H.K.; Long, C.; Bahaie, N.S.; Walcheck, B. Regulation of CXCR2 expression and function by a disintegrin and metalloprotease-17 (ADAM17). J. Leukoc. Biol. 2015, 97, 447–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wu, J.; Newton, R.; Bahaie, N.S.; Long, C.; Walcheck, B. ADAM17 cleaves CD16b (FcgammaRIIIb) in human neutrophils. Biochim. Biophys. Acta 2013, 1833, 680–685. [Google Scholar] [CrossRef] [Green Version]
- Scheller, J.; Chalaris, A.; Garbers, C.; Rose-John, S. ADAM17: A molecular switch to control inflammation and tissue regeneration. Trends Immunol. 2011, 32, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Long, C.; Wang, Y.; Herrera, A.H.; Horiuchi, K.; Walcheck, B. In vivo role of leukocyte ADAM17 in the inflammatory and host responses during E. coli-mediated peritonitis. J. Leukoc. Biol. 2010, 87, 1097–1101. [Google Scholar] [CrossRef] [Green Version]
- Mishra, H.K.; Johnson, T.J.; Seelig, D.M.; Walcheck, B. Targeting ADAM17 in leukocytes increases neutrophil recruitment and reduces bacterial spread during polymicrobial sepsis. J. Leukoc. Biol. 2016, 100, 999–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, C.; Hosseinkhani, M.R.; Wang, Y.; Sriramarao, P.; Walcheck, B. ADAM17 activation in circulating neutrophils following bacterial challenge impairs their recruitment. J. Leukoc. Biol. 2012, 92, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; He, J.; Chen, F.; Cai, Y.; Zhao, J.; Lin, Y.; Yin, Z.; Tao, H.; Shao, X.; Huang, P.; et al. Association Study Between Promoter Polymorphisms of ADAM17 and Progression of Sepsis. Cell Physiol. Biochem. 2016, 39, 1247–1261. [Google Scholar] [CrossRef] [PubMed]
- Kermarrec, N.; Selloum, S.; Plantefeve, G.; Chosidow, D.; Paoletti, X.; Lopez, A.; Mantz, J.; Desmonts, J.M.; Gougerot-Pocidalo, M.A.; Chollet-Martin, S. Regulation of peritoneal and systemic neutrophil-derived tumor necrosis factor-alpha release in patients with severe peritonitis: Role of tumor necrosis factor-alpha converting enzyme cleavage. Crit. Care Med. 2005, 33, 1359–1364. [Google Scholar] [CrossRef]
- Peng, L.; Cook, K.; Xu, L.; Cheng, L.; Damschroder, M.; Gao, C.; Wu, H.; Dall’Acqua, W.F. Molecular basis for the mechanism of action of an anti-TACE antibody. MAbs 2016, 8, 1598–1605. [Google Scholar] [CrossRef]
- Rios-Doria, J.; Sabol, D.; Chesebrough, J.; Stewart, D.; Xu, L.; Tammali, R.; Cheng, L.; Du, Q.; Schifferli, K.; Rothstein, R.; et al. A Monoclonal Antibody to ADAM17 Inhibits Tumor Growth by Inhibiting EGFR and Non-EGFR-Mediated Pathways. Mol. Cancer Ther. 2015, 14, 1637–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, H.K.; Pore, N.; Michelotti, E.F.; Walcheck, B. Anti-ADAM17 monoclonal antibody MEDI3622 increases IFNgamma production by human NK cells in the presence of antibody-bound tumor cells. Cancer Immunol. Immunother. 2018, 67, 1407–1416. [Google Scholar] [CrossRef]
- Dreymueller, D.; Theodorou, K.; Donners, M.; Ludwig, A. Fine Tuning Cell Migration by a Disintegrin and Metalloproteinases. Mediators Inflamm. 2017, 2017, 9621724. [Google Scholar] [CrossRef]
- Buscher, K.; Wang, H.; Zhang, X.; Striewski, P.; Wirth, B.; Saggu, G.; Lutke-Enking, S.; Mayadas, T.N.; Ley, K.; Sorokin, L.; et al. Protection from septic peritonitis by rapid neutrophil recruitment through omental high endothelial venules. Nat. Commun. 2016, 7, 10828. [Google Scholar] [CrossRef]
- Rello, J.; Valenzuela-Sanchez, F.; Ruiz-Rodriguez, M.; Moyano, S. Sepsis: A Review of Advances in Management. Adv. Ther. 2017, 34, 2393–2411. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Gutierrez, B.; Bonomo, R.A.; Carmeli, Y.; Paterson, D.L.; Almirante, B.; Martinez-Martinez, L.; Oliver, A.; Calbo, E.; Pena, C.; Akova, M.; et al. Ertapenem for the treatment of bloodstream infections due to ESBL-producing Enterobacteriaceae: A multinational pre-registered cohort study. J. Antimicrob. Chemother. 2016, 71, 1672–1680. [Google Scholar] [CrossRef]
- Wen, H. Sepsis induced by cecal ligation and puncture. Methods Mol. Biol. 2013, 1031, 117–124. [Google Scholar]
- Halbach, J.L.; Wang, A.W.; Hawisher, D.; Cauvi, D.M.; Lizardo, R.E.; Rosas, J.; Reyes, T.; Escobedo, O.; Bickler, S.W.; Coimbra, R.; et al. Why Antibiotic Treatment Is Not Enough for Sepsis Resolution: An Evaluation in an Experimental Animal Model. Infect. Immun. 2017, 85, e00664-17. [Google Scholar] [CrossRef] [Green Version]
- Chousterman, B.G.; Swirski, F.K.; Weber, G.F. Cytokine storm and sepsis disease pathogenesis. Semin. Immunopathol. 2017, 39, 517–528. [Google Scholar] [CrossRef]
- Black, R.A.; Rauch, C.T.; Kozlosky, C.J.; Peschon, J.J.; Slack, J.L.; Wolfson, M.F.; Castner, B.J.; Stocking, K.L.; Reddy, P.; Srinivasan, S.; et al. A metalloproteinase disintegrin that releases tumour-necrosis factor-alpha from cells. Nature 1997, 385, 729–733. [Google Scholar] [CrossRef]
- Lerman, Y.V.; Kim, M. Neutrophil migration under normal and sepsis conditions. Cardiov. Hematol. Disord. Drug Targets 2015, 15, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Li, X.; Zhang, X.; Li, Z.; Wang, L.; Sun, Y.; Liu, Z.; Ma, X. Elevated levels of plasma TNF-alpha are associated with microvascular endothelial dysfunction in patients with sepsis through activating the NF-kappaB and p38 mitogen-activated protein kinase in endothelial cells. Shock 2014, 41, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Nedeva, C.; Menassa, J.; Puthalakath, H. Sepsis: Inflammation Is a Necessary Evil. Front. Cell Dev. Biol. 2019, 7, 108. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Gu, H.; Yu, M.; Yang, P.; Zhang, M.; Ba, H.; Yin, Y.; Wang, J.; Yin, B.; Zhou, X.; et al. Inhibition of transmembrane TNF-alpha shedding by a specific antibody protects against septic shock. Cell Death Dis. 2019, 10, 586. [Google Scholar] [CrossRef] [Green Version]
- Ramnath, R.D.; Ng, S.W.; Guglielmotti, A.; Bhatia, M. Role of MCP-1 in endotoxemia and sepsis. Int. Immunopharmacol. 2008, 8, 810–818. [Google Scholar] [CrossRef]
- Souto, F.O.; Alves-Filho, J.C.; Turato, W.M.; Auxiliadora-Martins, M.; Basile-Filho, A.; Cunha, F.Q. Essential role of CCR2 in neutrophil tissue infiltration and multiple organ dysfunction in sepsis. Am. J. Respir. Crit. Care Med. 2011, 183, 234–242. [Google Scholar] [CrossRef]
- Ayala, A.; Chung, C.S.; Song, G.Y.; Chaudry, I.H. IL-10 mediation of activation-induced TH1 cell apoptosis and lymphoid dysfunction in polymicrobial sepsis. Cytokine 2001, 14, 37–48. [Google Scholar] [CrossRef]
- Li, X.; Xu, Z.; Pang, X.; Huang, Y.; Yang, B.; Yang, Y.; Chen, K.; Liu, X.; Mao, P.; Li, Y. Interleukin-10/lymphocyte ratio predicts mortality in severe septic patients. PLoS ONE 2017, 12, e0179050. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, N.; Rose-John, S. ADAM17 Activity and IL-6 Trans-Signaling in Inflammation and Cancer. Cancers 2019, 11, 1736. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, N.; Schmidt, S.; Schwarz, J.; Dohr, D.; Lokau, J.; Scheller, J.; Garbers, C.; Chalaris, A.; Rose-John, S.; Rabe, B. Circulating Soluble IL-6R but Not ADAM17 Activation Drives Mononuclear Cell Migration in Tissue Inflammation. J. Immunol. 2016, 197, 3705–3715. [Google Scholar] [CrossRef]
- Zhanel, G.G.; Johanson, C.; Embil, J.M.; Noreddin, A.; Gin, A.; Vercaigne, L.; Hoban, D.J. Ertapenem: Review of a new carbapenem. Expert Rev. Anti Infect. Ther. 2005, 3, 23–39. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mishra, H.K.; Ma, J.; Mendez, D.; Hullsiek, R.; Pore, N.; Walcheck, B. Blocking ADAM17 Function with a Monoclonal Antibody Improves Sepsis Survival in a Murine Model of Polymicrobial Sepsis. Int. J. Mol. Sci. 2020, 21, 6688. https://doi.org/10.3390/ijms21186688
Mishra HK, Ma J, Mendez D, Hullsiek R, Pore N, Walcheck B. Blocking ADAM17 Function with a Monoclonal Antibody Improves Sepsis Survival in a Murine Model of Polymicrobial Sepsis. International Journal of Molecular Sciences. 2020; 21(18):6688. https://doi.org/10.3390/ijms21186688
Chicago/Turabian StyleMishra, Hemant K., Jing Ma, Daniel Mendez, Robert Hullsiek, Nabendu Pore, and Bruce Walcheck. 2020. "Blocking ADAM17 Function with a Monoclonal Antibody Improves Sepsis Survival in a Murine Model of Polymicrobial Sepsis" International Journal of Molecular Sciences 21, no. 18: 6688. https://doi.org/10.3390/ijms21186688