Phlorotannins from Fucus vesiculosus: Modulation of Inflammatory Response by Blocking NF-κB Signaling Pathway

 ,
, 
 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Antioxidant Properties of Fucus Vesiculosus Crude Extract (CRD) and Phlorotannin-Enriched Fraction (EtOAc)
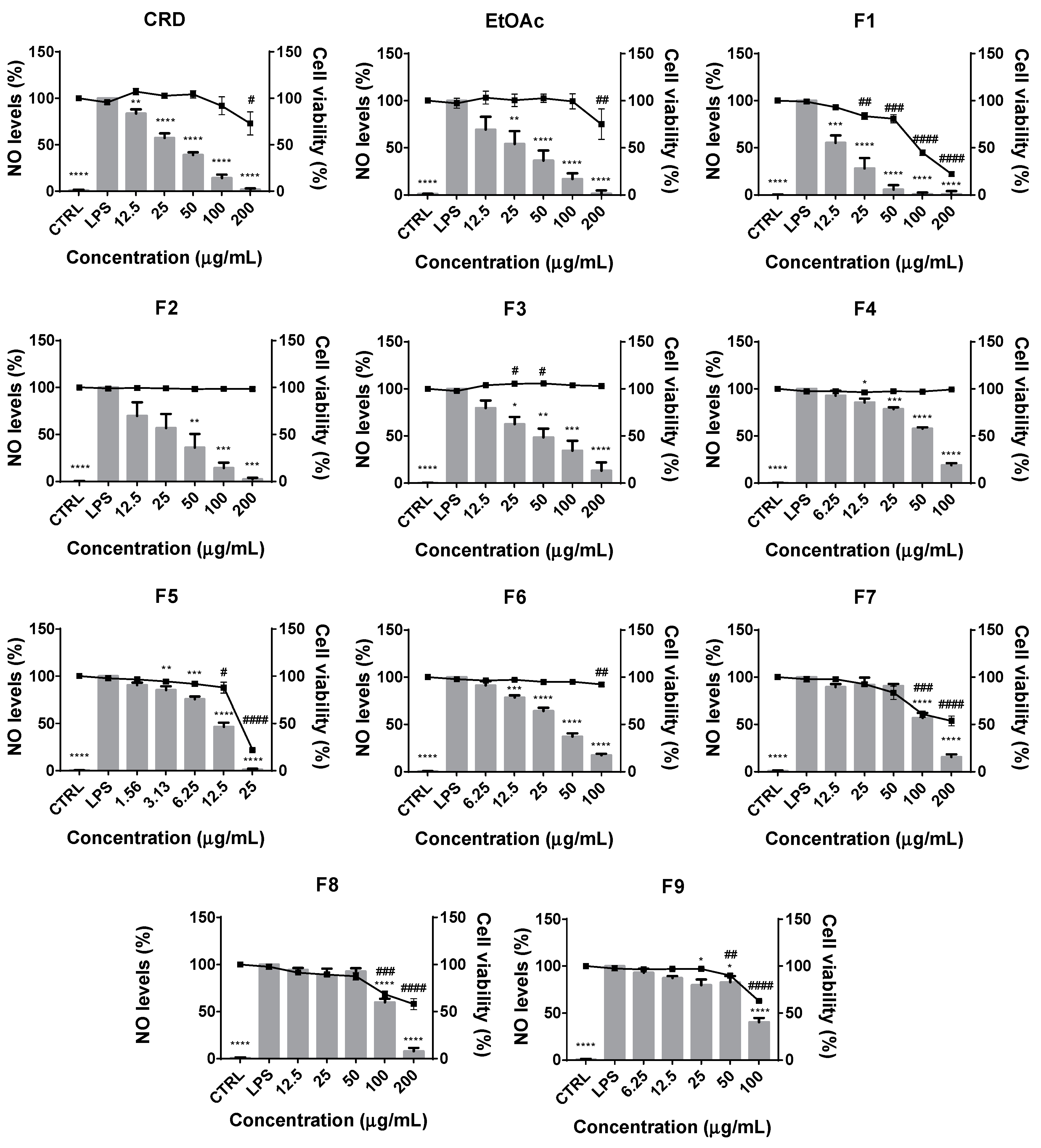
2.2. Anti-Inflammatory Properties of Fucus Vesiculosus CRD and Its Fractions in Raw 264.7 Cells
2.2.1. Effects on Cell Viability and LPS-Induced NO• Production
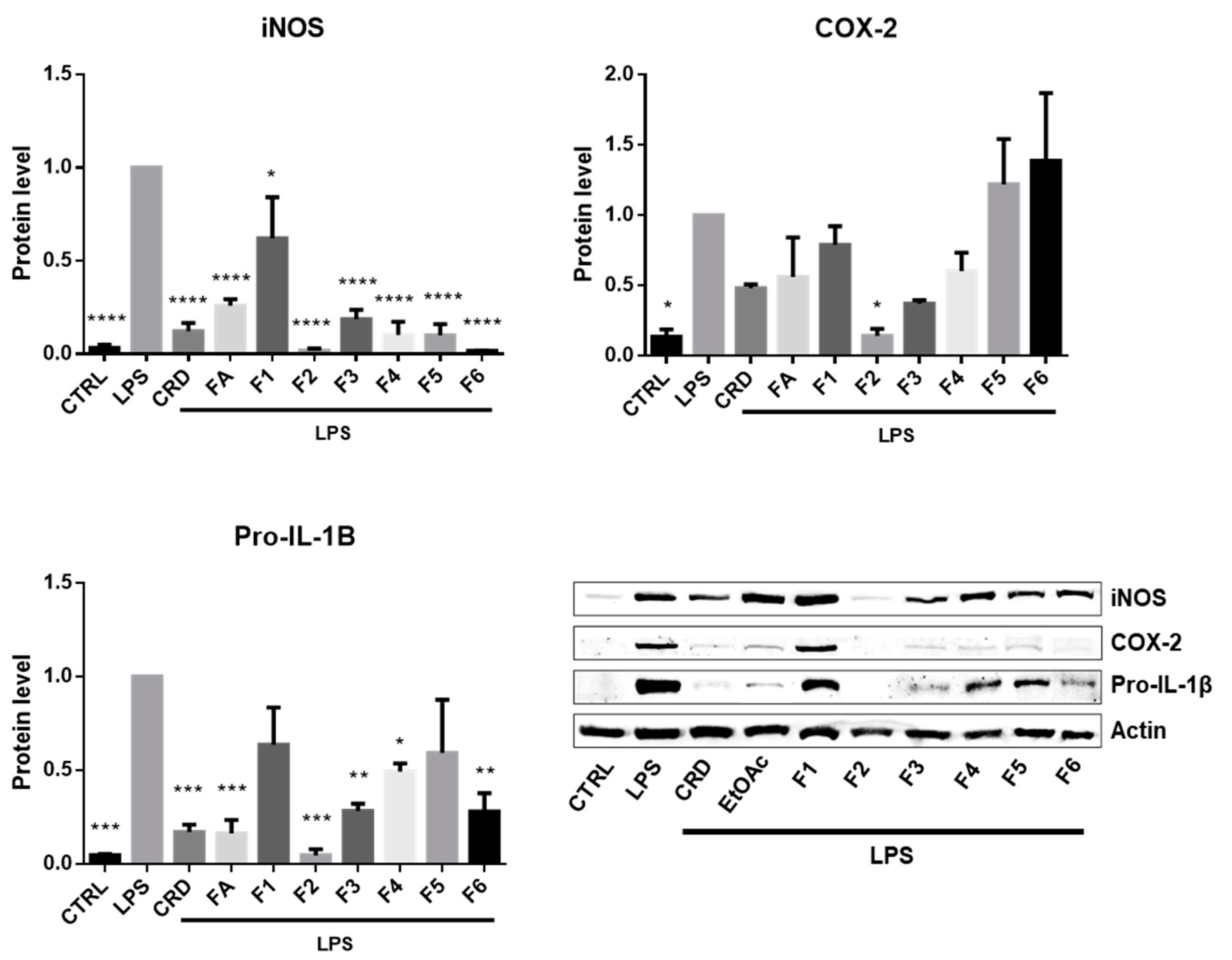
2.2.2. Effects on the Expression of iNOS, COX-2 and IL-1β
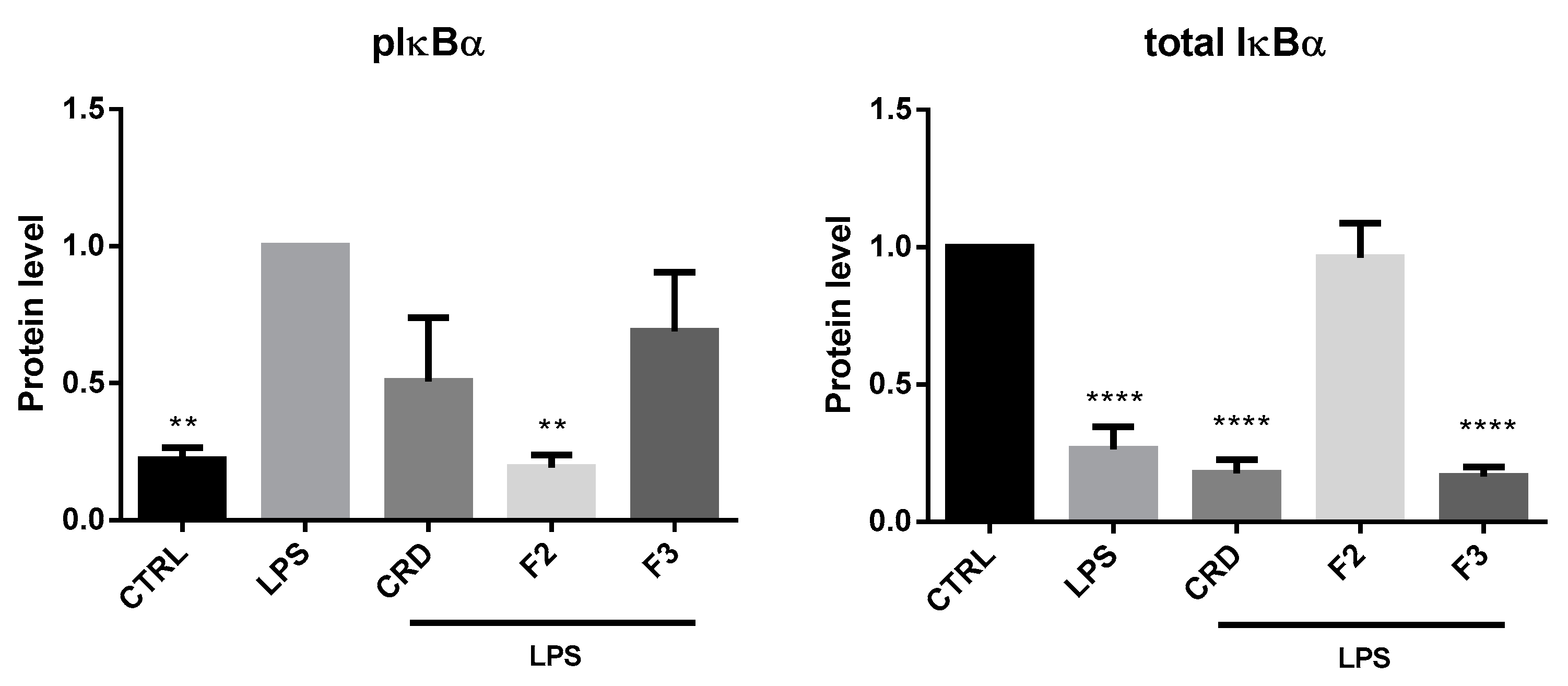
2.2.3. Effects on the NF-κB Signaling Pathway
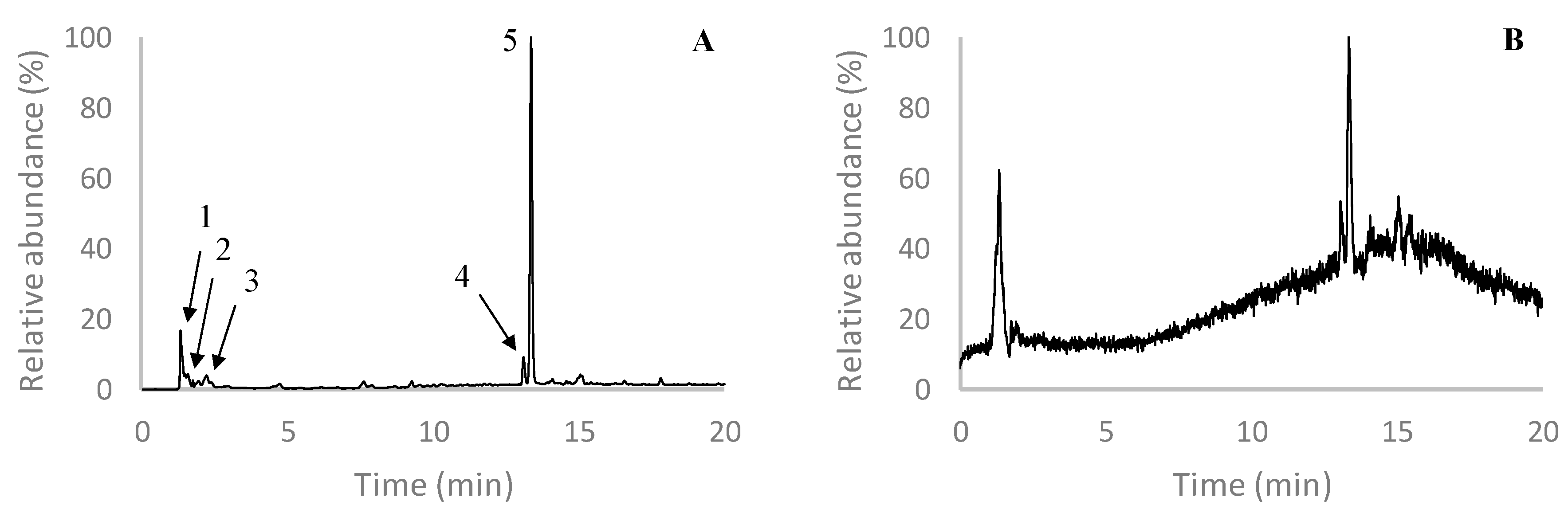
2.3. Characterization of F2
3. Materials and Methods
3.1. Chemicals
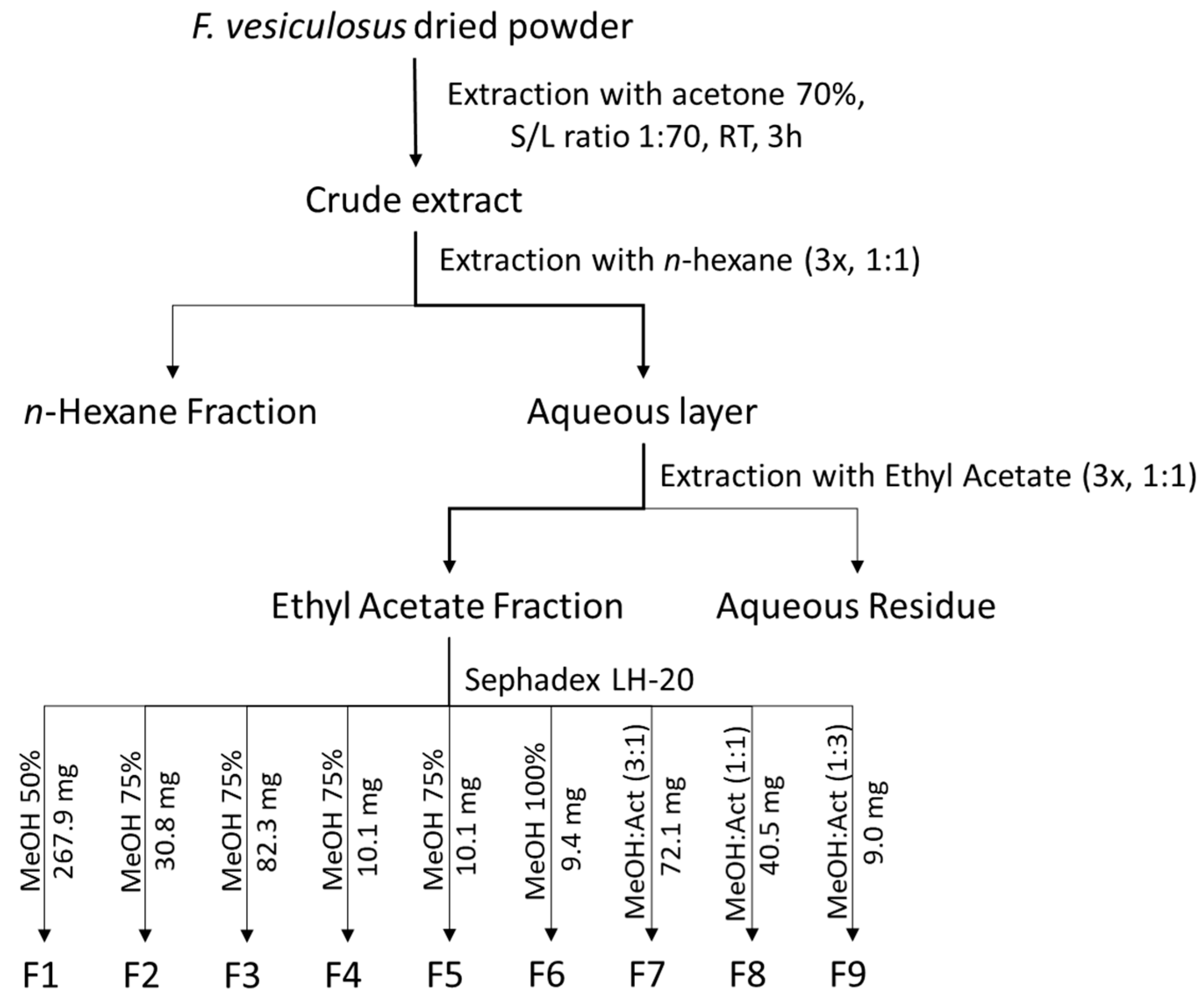
3.2. Extraction and Purification of Phlorotannins from Fucus vesiculosus
3.3. Antioxidant Experiments
3.3.1. ORAC
3.3.2. Superoxide Anion (O2•−) Scavenging Assay
3.3.3. Xanthine Oxidase Assay
3.3.4. Nitric Oxide (NO•) Assay
3.4. Anti-Inflammatory Experiments
3.4.1. Cell Culture
3.4.2. Assessment of Cell Viability
3.4.3. Inhibition of LPS-Stimulated NO•
3.4.4. Preparation of Total protein Extracts and Western Blotting
3.5. UHPLC-DAD-ESI/MS Analysis
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- GBIF Secretariat. GBIF Backbone Taxonomy-Fucus vesiculosus L. Available online: https://doi.org/10.15468/dl.r83ttf (accessed on 21 January 2020).
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Phycochemical Constituents and Biological Activities of Fucus spp. Mar. Drugs 2018, 16, 249. [Google Scholar] [CrossRef] [PubMed]
- Catarino, D.M.; Silva, M.A.; Cardoso, M.S. Fucaceae: A Source of Bioactive Phlorotannins. Int. J. Mol. Sci. 2017, 18, 1327. [Google Scholar] [CrossRef] [PubMed]
- Pal Singh, I.; Bharate, S.B. Phloroglucinol compounds of natural origin. Nat. Prod. Rep. 2006, 23, 558–591. [Google Scholar] [CrossRef] [PubMed]
- Ummat, V.; Tiwari, B.K.; Jaiswal, A.K.; Condon, K.; Garcia-Vaquero, M.; O’Doherty, J.; O’Donnell, C.; Rajauria, G. Optimisation of ultrasound frequency, extraction time and solvent for the recovery of polyphenols, phlorotannins and associated antioxidant activity from brown seaweeds. Mar. Drugs 2020. [Google Scholar] [CrossRef]
- Boi, V.N.; Cuong, D.X.; Vinh, P.T.K.V. Effects of extraction conditions over the phlorotannin content and antioxidant activity of extract from brown algae Sargassum serratum. Free Radic. Antioxid. 2017, 7, 115–122. [Google Scholar] [CrossRef]
- Sánchez-Camargo, A.D.P.; Montero, L.; Stiger-Pouvreau, V.; Tanniou, A.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Considerations on the use of enzyme-assisted extraction in combination with pressurized liquids to recover bioactive compounds from algae. Food Chem. 2016, 192, 67–74. [Google Scholar] [CrossRef]
- He, Z.; Chen, Y.; Chen, Y.; Liu, H.; Yuan, G.; Fan, Y.; Chen, K. Optimization of the microwave-assisted extraction of phlorotannins from Saccharina japonica Aresch and evaluation of the inhibitory effects of phlorotannin-containing extracts on HepG2 cancer cells. Chin. J. Oceanol. Limnol. 2013, 31, 1045–1054. [Google Scholar] [CrossRef]
- Obluchinskaya, E.D.; Daurtseva, A.V.; Pozharitskaya, O.N.; Flisyuk, E.V.; Shikov, A.N. Natural Deep Eutectic Solvents as Alternatives for Extracting Phlorotannins from Brown Algae. Pharm. Chem. J. 2019. [Google Scholar] [CrossRef]
- Heavisides, E.; Rouger, C.; Reichel, A.F.; Ulrich, C.; Wenzel-Storjohann, A.; Sebens, S.; Tasdemir, D. Seasonal variations in the metabolome and bioactivity profile of fucus vesiculosus extracted by an optimised, pressurised liquid extraction protocol. Mar. Drugs 2018. [Google Scholar] [CrossRef]
- Vázquez-Rodríguez, B.; Gutiérrez-Uribe, J.A.; Antunes-Ricardo, M.; Santos-Zea, L.; Cruz-Suárez, L.E. Ultrasound-assisted extraction of phlorotannins and polysaccharides from Silvetia compressa (Phaeophyceae). J. Appl. Phycol. 2020. [Google Scholar] [CrossRef]
- Koivikko, R.; Loponen, J.; Pihlaja, K.; Jormalainen, V. High-performance liquid chromatographic analysis of phlorotannins from the brown alga Fucus vesiculosus. Phytochem. Anal. 2007, 18, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Koivikko, R.; Loponen, J.; Honkanen, T.; Jormalainen, V. Contents of soluble, cell-wall-bound and exuded phlorotannins in the brown alga Fucus vesiculosus, with implications on their ecological functions. J. Chem. Ecol. 2005, 31, 195–212. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.; Silva, A.; Mateus, N.; Cardoso, S. Optimization of Phlorotannins Extraction from Fucus vesiculosus and Evaluation of Their Potential to Prevent Metabolic Disorders. Mar. Drugs 2019, 17, 162. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jónsdóttir, R.; Liu, H.; Gu, L.; Kristinsson, H.G.; Raghavan, S.; Ólafsdóttir, G. Antioxidant capacities of phlorotannins extracted from the brown algae Fucus vesiculosus. J. Agric. Food Chem. 2012, 60, 5874–5883. [Google Scholar] [CrossRef] [PubMed]
- Breton, F.; Cérantola, S.; Ar Gall, E. Distribution and radical scavenging activity of phenols in Ascophyllum nodosum (Phaeophyceae). J. Exp. Mar. Bio. Ecol. 2011, 399, 167–172. [Google Scholar] [CrossRef]
- Ferreres, F.; Lopes, G.; Gil-Izquierdo, A.; Andrade, P.B.; Sousa, C.; Mouga, T.; Valentão, P. Phlorotannin extracts from fucales characterized by HPLC-DAD-ESI-MSn: Approaches to hyaluronidase inhibitory capacity and antioxidant properties. Mar. Drugs 2012, 10, 2766–2781. [Google Scholar] [CrossRef]
- Yang, Y.I.; Jung, S.H.; Lee, K.T.; Choi, J.H. 8,8′-Bieckol, isolated from edible brown algae, exerts its anti-inflammatory effects through inhibition of NF-κB signaling and ROS production in LPS-stimulated macrophages. Int. Immunopharmacol. 2014, 23, 460–468. [Google Scholar] [CrossRef]
- Yayeh, T.; Im, E.J.; Kwon, T.H.; Roh, S.S.; Kim, S.; Kim, J.H.; Hong, S.B.; Cho, J.Y.; Park, N.H.; Rhee, M.H. Hemeoxygenase 1 partly mediates the anti-inflammatory effect of dieckol in lipopolysaccharide stimulated murine macrophages. Int. Immunopharmacol. 2014, 22, 51–58. [Google Scholar] [CrossRef]
- Lee, M.S.; Kwon, M.S.; Choi, J.W.; Shin, T.; No, H.K.; Choi, J.S.; Byun, D.S.; Il Kim, J.; Kim, H.R. Anti-inflammatory activities of an ethanol extract of Ecklonia stolonifera in lipopolysaccharide-stimulated RAW 264.7 murine macrophage cells. J. Agric. Food Chem. 2012, 60, 9120–9129. [Google Scholar] [CrossRef]
- Ashley, N.T.; Weil, Z.M.; Nelson, R.J. Inflammation: Mechanisms, Costs, and Natural Variation. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 385–406. [Google Scholar] [CrossRef]
- Medzhitov, R. Inflammation 2010: New adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef]
- Catarino, M.D.; Talhi, O.; Rabahi, A.; Silva, A.M.S.; Cardoso, S.M. The anti-inflammatory potential of flavonoids: Mechanistic aspects. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2016; Vol. 48, pp. 65–99. ISBN 9780444636027. [Google Scholar]
- Gupta, S.C.; Sundaram, C.; Reuter, S.; Aggarwal, B.B. Inhibiting NF-κB activation by small molecules as a therapeutic strategy. Biochim. Biophys. Acta-Gene Regul. Mech. 2010, 1799, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Serasanambati, M.; Chilakapati, S.R. Function of Nuclear Factor Kappa B (NF-kB) in Human Diseases-A Review. South. Indian J. Biol. Sci. 2016. [Google Scholar] [CrossRef]
- Phasanasophon, K.; Kim, S.M. Anti-inflammatory activity of the phlorotannin trifuhalol A using LPS-stimulated RAW264.7 cells through NF-κB and MAPK main signaling pathways. Nat. Prod. Commun. 2019. [Google Scholar] [CrossRef]
- Sugiura, Y.; Tanaka, R.; Katsuzaki, H.; Imai, K.; Matsushita, T. The anti-inflammatory effects of phlorotannins from Eisenia arborea on mouse ear edema by inflammatory inducers. J. Funct. Foods 2013, 5, 2019–2023. [Google Scholar] [CrossRef]
- Kim, S.K.; Himaya, S.W.A. Medicinal effects of phlorotannins from marine brown algae. In Advances in Food and Nutrition Research; Kim, S., Ed.; Academic Press: San Diego, CA, USA, 2011; Vol. 64, pp. 97–109. ISBN 9780123876690. [Google Scholar]
- Barbosa, M.; Lopes, G.; Ferreres, F.; Andrade, P.B.; Pereira, D.M.; Gil-Izquierdo, Á.; Valentão, P. Phlorotannin extracts from Fucales: Marine polyphenols as bioregulators engaged in inflammation-related mediators and enzymes. Algal Res. 2017, 28, 1–8. [Google Scholar] [CrossRef]
- Lopes, G.; Sousa, C.; Silva, L.R.; Pinto, E.; Andrade, P.B.; Bernardo, J.; Mouga, T.; Valentão, P. Can phlorotannins purified extracts constitute a novel pharmacological alternative for microbial infections with associated inflammatory conditions? PLoS ONE 2012, 7, e31145. [Google Scholar] [CrossRef]
- Zaragozá, M.C.; López, D.; Sáiz, M.P.; Poquet, M.; Pérez, J.; Puig-Parellada, P.; Màrmol, F.; Simonetti, P.; Gardana, C.; Lerat, Y.; et al. Toxicity and antioxidant activity in vitro and in vivo of two Fucus vesiculosus extracts. J. Agric. Food Chem. 2008, 56, 7773–7780. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Cruz, M.T.; Cardoso, S.M. Antioxidant and anti-inflammatory activities of Geranium robertianum L. decoctions. Food Funct. 2017, 8, 3355–3365. [Google Scholar] [CrossRef]
- Wang, T.; Jónsdóttir, R.; Ólafsdóttir, G. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Ho, S.-C.; Tsai, T.-H.; Tsai, P.-J.; Lin, C.-C. Protective capacities of certain spices against peroxynitrite-mediated biomolecular damage. Food Chem. Toxicol. 2008, 46, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.F.; Chen, Y.; Black, R.M.; Brown, P.H.; Lyle, B.J.; Liu, R.H.; Ou, B. Type 2 diabetes-related bioactivities of coffee: Assessment of antioxidant activity, NF-κB inhibition, and stimulation of glucose uptake. Food Chem. 2011, 124, 914–920. [Google Scholar] [CrossRef]
- Lopes, G.; Barbosa, M.; Andrade, P.B.; Valentão, P. Phlorotannins from Fucales: Potential to control hyperglycemia and diabetes-related vascular complications. J. Appl. Phycol. 2019, 31, 3143–3152. [Google Scholar] [CrossRef]
- Catarino, M.D.; Alves-Silva, J.M.; Pereira, O.R.; Cardoso, S.M. Antioxidant Capacities of Flavones and Benefits in Oxidative-Stress Related Diseases. Curr. Top. Med. Chem. 2015, 15, 105–119. [Google Scholar] [CrossRef]
- Lin, H.C.; Tsai, S.H.; Chen, C.S.; Chang, Y.C.; Lee, C.M.; Lai, Z.Y.; Lin, C.M. Structure-activity relationship of coumarin derivatives on xanthine oxidase-inhibiting and free radical-scavenging activities. Biochem. Pharmacol. 2008, 75, 1416–1425. [Google Scholar] [CrossRef]
- Tanniou, A.; Vandanjon, L.; Incera, M.; Serrano Leon, E.; Husa, V.; Le Grand, J.; Nicolas, J.L.; Poupart, N.; Kervarec, N.; Engelen, A.; et al. Assessment of the spatial variability of phenolic contents and associated bioactivities in the invasive alga Sargassum muticum sampled along its European range from Norway to Portugal. J. Appl. Phycol. 2014, 26, 1215–1230. [Google Scholar] [CrossRef]
- Kellogg, J.; Esposito, D.; Grace, M.H.; Komarnytsky, S.; Lila, M.A. Alaskan seaweeds lower inflammation in RAW 264.7 macrophages and decrease lipid accumulation in 3T3-L1 adipocytes. J. Funct. Foods 2015, 15, 396–407. [Google Scholar] [CrossRef]
- Dutot, M.; Fagon, R.; Hemon, M.; Rat, P. Antioxidant, anti-inflammatory, and anti-senescence activities of a phlorotannin-rich natural extract from Brown Seaweed Ascophyllum nodosum. Appl. Biochem. Biotechnol. 2012, 167, 2234–2240. [Google Scholar] [CrossRef]
- Cillard, J.; Bonnaure-mallet, M. Silver-zeolite combined to polyphenol-rich extracts of Ascophyllum nodosum: Potential active role in prevention of periodontal diseases. PLoS ONE 2014, 9, 1–8. [Google Scholar] [CrossRef]
- Bahar, B.; O’Doherty, J.V.; Smyth, T.J.; Sweeney, T. A comparison of the effects of an Ascophyllum nodosum ethanol extract and its molecular weight fractions on the inflammatory immune gene expression in-vitro and ex-vivo. Innov. Food Sci. Emerg. Technol. 2016, 37, 276–285. [Google Scholar] [CrossRef]
- Masters, S.L.; Simon, A.; Aksentijevich, I.; Kastner, D.L. Horror Autoinflammaticus: The Molecular Pathophysiology of Autoinflammatory Disease. Annu. Rev. Immunol. 2009. [Google Scholar] [CrossRef] [PubMed]
- Rudolphi, K.; Gerwin, N.; Verzijl, N.; van der Kraan, P.; van den Berg, W. Pralnacasan, an inhibitor of interleukin-1β converting enzyme, reduces joint damage in two murine models of osteoarthritis. Osteoarthr. Cartil. 2003. [Google Scholar] [CrossRef]
- Callejas, N.A.; Castrillo, A.; Boscá, L.; Martín-Sanz, P. Inhibition of prostaglandin synthesis up-regulates cyclooxygenase-2 induced by lipopolysaccharide and peroxisomal proliferators. J. Pharmacol. Exp. Ther. 1999. [Google Scholar]
- Joung, E.-J.J.; Lee, M.-S.S.; Choi, J.-W.W.; Kim, J.S.J.-I.I.J.-S.; Shin, T.; Jung, B.-M.M.; Kim, J.S.J.-I.I.J.-S.; Kim, H.-R.R. Anti-inflammatory effects of phlorofucofuroeckol B-rich ethyl acetate fraction obtained from Myagropsis myagroides on lipopolysaccharide-stimulated RAW 264.7 cells and mouse edema. Int. Immunopharmacol. 2012, 14, 471–480. [Google Scholar] [CrossRef]
- Bahar, B.; O’Doherty, J.V.; Hayes, M.; Sweeney, T. Extracts of brown seaweeds can attenuate the bacterial lipopolysaccharide-induced pro-inflammatory response in the porcine colon ex vivo. J. Anim. Sci. 2012, 90, 46–48. [Google Scholar] [CrossRef]
- Bahar, B.; Doherty, J.V.O.; Smyth, T.J.; Ahmed, A.M.; Sweeney, T. A cold water extract of Fucus vesiculosus inhibits lipopolysaccharide (LPS) induced pro-inflammatory responses in the porcine colon ex-vivo model. Innov. Food Sci. Emerg. Technol. 2016, 37, 229–236. [Google Scholar] [CrossRef]
- Pautz, A.; Art, J.; Hahn, S.; Nowag, S.; Voss, C.; Kleinert, H. Regulation of the expression of inducible nitric oxide synthase. Nitric Oxide-Biol. Chem. 2010. [Google Scholar] [CrossRef]
- Wei, R.; Lee, M.S.; Lee, B.; Oh, C.W.; Choi, C.G.; Kim, H.R. Isolation and identification of anti-inflammatory compounds from ethyl acetate fraction of Ecklonia stolonifera and their anti-inflammatory action. J. Appl. Phycol. 2016. [Google Scholar] [CrossRef]
- Jung, W.K.; Ahn, Y.W.; Lee, S.H.; Choi, Y.H.; Kim, S.K.; Yea, S.S.; Choi, I.; Park, S.G.; Seo, S.K.; Lee, S.W.; et al. Ecklonia cava ethanolic extracts inhibit lipopolysaccharide-induced cyclooxygenase-2 and inducible nitric oxide synthase expression in BV2 microglia via the MAP kinase and NF-κB pathways. Food Chem. Toxicol. 2008, 47, 410–417. [Google Scholar] [CrossRef]
- Yang, Y.I.; Woo, J.H.; Seo, Y.J.; Lee, K.T.; Lim, Y.; Choi, J.H. Protective Effect of Brown Alga Phlorotannins against Hyper-inflammatory Responses in Lipopolysaccharide-Induced Sepsis Models. J. Agric. Food Chem. 2016, 64, 570–578. [Google Scholar] [CrossRef]
- Eom, S.H.; Lee, E.H.; Park, K.; Kwon, J.Y.; Kim, P.H.; Jung, W.K.; Kim, Y.M. Eckol from Eisenia bicyclis inhibits inflammation through the Akt/NF-κB signaling in Propionibacterium acnes-induced human keratinocyte Hacat cells. J. Food Biochem. 2017, 41, 1–10. [Google Scholar] [CrossRef]
- Kim, A.-R.; Lee, M.-S.; Shin, T.-S.; Hua, H.; Jang, B.-C.; Choi, J.-S.; Byun, D.-S.; Utsuki, T.; Ingram, D.; Kim, H.-R. Phlorofucofuroeckol A inhibits the LPS-stimulated iNOS and COX-2 expressions in macrophages via inhibition of NF-κB, Akt, and p38 MAPK. Toxicol. Vitr. 2011, 25, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Joe, M.J.; Kim, S.N.; Choi, H.Y.; Shin, W.S.; Park, G.M.; Kang, D.W.; Yong, K.K. The inhibitory effects of eckol and dieckol from Ecklonia stolonifera on the expression of matrix metalloproteinase-1 in human dermal fibroblasts. Biol. Pharm. Bull. 2006. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, K.B.W.R.; Jeong, D.H.; Ahn, D.H. Anti-inflammatory activity of ethanolic extract of Sargassum sagamianum in RAW 264.7 cells. Food Sci. Biotechnol. 2013, 22, 1113–1120. [Google Scholar] [CrossRef]
- Kim, M.E.; Jung, Y.C.; Jung, I.; Lee, H.W.; Youn, H.Y.; Lee, J.S. Anti-inflammatory effects of ethanolic extract from Sargassum horneri (Turner) C. Agardh on lipopolysaccharide-stimulated macrophage activation via NF-κB pathway regulation. Immunol. Invest. 2015, 44, 137–146. [Google Scholar] [CrossRef]
- Lopes, G.; Barbosa, M.; Vallejo, F.; Gil-Izquierdo, Á.; Andrade, P.B.; Valentão, P.; Pereira, D.M.; Ferreres, F. Profiling phlorotannins from Fucus spp. of the Northern Portuguese coastline: Chemical approach by HPLC-DAD-ESI/MSnand UPLC-ESI-QTOF/MS. Algal Res. 2018, 29, 113–120. [Google Scholar] [CrossRef]
- Hermund, D.B.; Plaza, M.; Turner, C.; Jónsdóttir, R.; Kristinsson, H.G.; Jacobsen, C.; Nielsen, K.F. Structure dependent antioxidant capacity of phlorotannins from Icelandic Fucus vesiculosus by UHPLC-DAD-ECD-QTOFMS. Food Chem. 2018, 240, 904–909. [Google Scholar] [CrossRef]
- Agregán, R.; Munekata, P.E.S.; Franco, D.; Dominguez, R.; Carballo, J.; Lorenzo, J.M. Phenolic compounds from three brown seaweed species using LC-DAD–ESI-MS/MS. Food Res. Int. 2017, 99, 979–985. [Google Scholar] [CrossRef]
- Montero, L.; Sánchez-Camargo, A.P.; García-Cañas, V.; Tanniou, A.; Stiger-Pouvreau, V.; Russo, M.; Rastrelli, L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Anti-proliferative activity and chemical characterization by comprehensive two-dimensional liquid chromatography coupled to mass spectrometry of phlorotannins from the brown macroalga Sargassum muticum collected on North-Atlantic coasts. J. Chromatogr. A 2016, 1428, 115–125. [Google Scholar] [CrossRef]
- Mezghani, S.; Csupor, D.; Bourguiba, I.; Hohmann, J.; Amri, M.; Bouaziz, M. Characterization of Phenolic Compounds of Ulva rigida (Chlorophycae) and Its Antioxidant Activity. Eur. J. Med. Plants 2016. [Google Scholar] [CrossRef]
- Kim, H.H.; Kim, H.S.; Ko, J.Y.; Kim, C.Y.; Lee, J.H.; Jeon, Y.J. A single-step isolation of useful antioxidant compounds from Ishige okamurae by using centrifugal partition chromatography. Fish. Aquat. Sci. 2016. [Google Scholar] [CrossRef]
- Heo, S.J.; Kim, J.P.; Jung, W.K.; Lee, N.H.; Kang, H.S.; Jun, E.M.; Park, S.H.; Kang, S.M.; Lee, Y.J.; Park, P.J.; et al. Identification of chemical structure and free radical scavenging activity of diphlorethohydroxycarmalol isolated from a brown alga, Ishige okamurae. J. Microbiol. Biotechnol. 2008. [Google Scholar]
- Pereira, O.; Catarino, M.; Afonso, A.; Silva, A.; Cardoso, S. Salvia elegans, Salvia greggii and Salvia officinalis Decoctions: Antioxidant Activities and Inhibition of Carbohydrate and Lipid Metabolic Enzymes. Molecules 2018, 23, 3169. [Google Scholar] [CrossRef]
- Barbosa, J.M. Preparação e Avaliação da Atividade Anti-Inflamatória de novos Derivados Polifenólicos Glicosilados. Master’s Thesis, Faculty of Pharmacy, University of Coimbra, Coimbra, Portugal, 2016. [Google Scholar]
- Jamalzadeh, L.; Ghafoori, H.; Sariri, R.; Rabuti, H.; Nasirzade, J.; Hasani, H.; Aghamaali, M.R. Cytotoxic Effects of Some Common Organic Solvents on MCF-7, RAW-264.7 and Human Umbilical Vein Endothelial Cells. Avicenna J. Med. Biochem. 2016. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | RCOO• (μmol TE/g ext) (1) | O2•− (IC50 μg/mL) (2) | NO• (IC50 μg/mL) (3) | XO (IC50 μg/mL) (4) |
|---|---|---|---|---|
| CRD | 3395.04 ± 211.4 a | 98.7 ± 11.1a | 75.2 ± 5.1a | 2.8 ± 0.4a |
| EtOAc | 2986.04 ± 338.7 b | 268.0 ± 20.1b | 235.9 ± 19.5b | 1.2 ± 0.2b |
| Standard | - | 7.8 ± 0.5c | 212.1 ± 9.7b | 0.1 ± 0.01c |
| Peak | RT (min) | [M − H]− (m/z) | MS/MS Ions (-loss)* | Tentative Assignment |
|---|---|---|---|---|
| 1 | 1.4 | 497 | 479 (-18), 331 (-PGU-44), 461 (-18-18), 435 (-44-18), 453 (-44), 413 (-84), 395 (-84-18), 347 (-150), 305 (-192), 165(-2PGU-84), 315 (-PGU-44-18) | Tetrafucol |
| 2 | 2.0 | 497 | 479 (-18), 331 (-PGU-44), 461 (-18-18), 435 (-44-18), 453 (-44), 395 (-84-18), 305 (-192), 165 (-2PGU-84), 315 (-PGU-44-18), 371 (-PGU), 353 (-PGU-18) | Fucophlorethol |
| 3 | 2.4 | 511 | 493 (-18), 449 (-44-18), 475 (-18-18), 467 (-44), 439 (-72), 411 (-84-16), 405 (-106), 345 (-PGU-44), 301 (-210), 331 (-180), 347 (-164), 385 (-PGU), 395 (-98-18), 369 (-PGU-16), 351 (-PGU-84), 329 (-PGU-44-18), 313 (-PGU-72) | Diphlorethohydroxycarmalol |
| 529 | 511 (-18), 493 (-18-18), 411 (-84-16-18), 467 (-44), 429 (-84-16), 439 (-90), 347 (-PGU-16-44) | Hydroxytetrafuhalol | ||
| 4 | 13.1 | 587 | 507 (-80), 523 (-64), 229 (-PGU-108-80-44), 277 (-230-80), 489 (-80-18), 383 (-PGU-80), 399 (-108-80), 463 (-PGU), 275 (-232-80), 569 (-18) | Phlorotannin derivative |
| 5 | 13.4 | 507 | 489 (-18), 277 (-230), 229 (-PGU-108-44), 461 (-46), 463 (-44), 479 (-28), 445 (-44-18), 275 (-232), 261 (-246), 245 (-262), 421 (-86), 297 (-PGU-84) | Phlorotannin derivative |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catarino, M.D.; Silva, A.; Cruz, M.T.; Mateus, N.; Silva, A.M.S.; Cardoso, S.M. Phlorotannins from Fucus vesiculosus: Modulation of Inflammatory Response by Blocking NF-κB Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 6897. https://doi.org/10.3390/ijms21186897
Catarino MD, Silva A, Cruz MT, Mateus N, Silva AMS, Cardoso SM. Phlorotannins from Fucus vesiculosus: Modulation of Inflammatory Response by Blocking NF-κB Signaling Pathway. International Journal of Molecular Sciences. 2020; 21(18):6897. https://doi.org/10.3390/ijms21186897
Chicago/Turabian StyleCatarino, Marcelo D., Ana Silva, Maria T. Cruz, Nuno Mateus, Artur M. S. Silva, and Susana M. Cardoso. 2020. "Phlorotannins from Fucus vesiculosus: Modulation of Inflammatory Response by Blocking NF-κB Signaling Pathway" International Journal of Molecular Sciences 21, no. 18: 6897. https://doi.org/10.3390/ijms21186897
APA StyleCatarino, M. D., Silva, A., Cruz, M. T., Mateus, N., Silva, A. M. S., & Cardoso, S. M. (2020). Phlorotannins from Fucus vesiculosus: Modulation of Inflammatory Response by Blocking NF-κB Signaling Pathway. International Journal of Molecular Sciences, 21(18), 6897. https://doi.org/10.3390/ijms21186897