Expression of Fbp2, a Newly Discovered Constituent of Memory Formation Mechanisms, Is Regulated by Astrocyte–Neuron Crosstalk

 , , ,
, , ,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
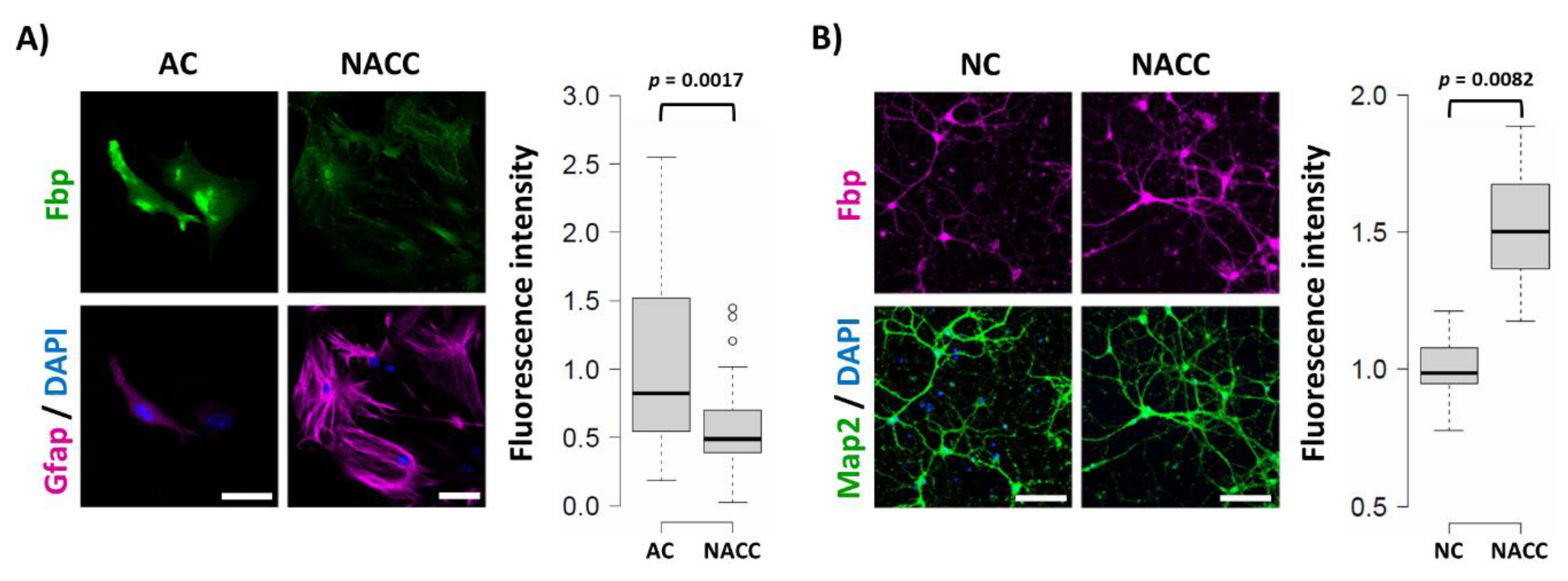
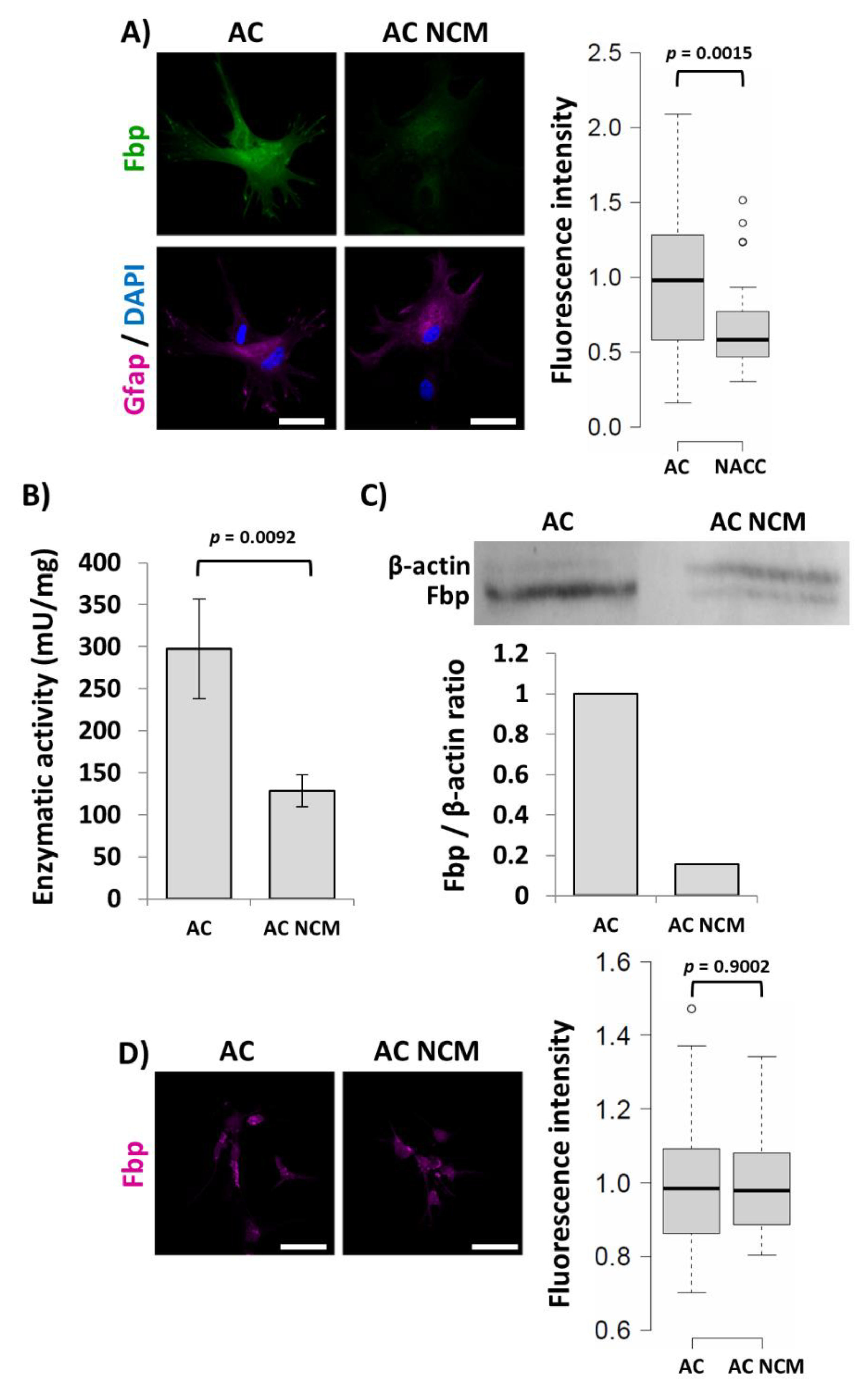
2.1. Fbp2 Expression in Hippocampal Cells Depends on Cell Culture Conditions
2.2. Fbp2 Activity and Protein Level Reduction Is Not Reflected by mRNA Changes
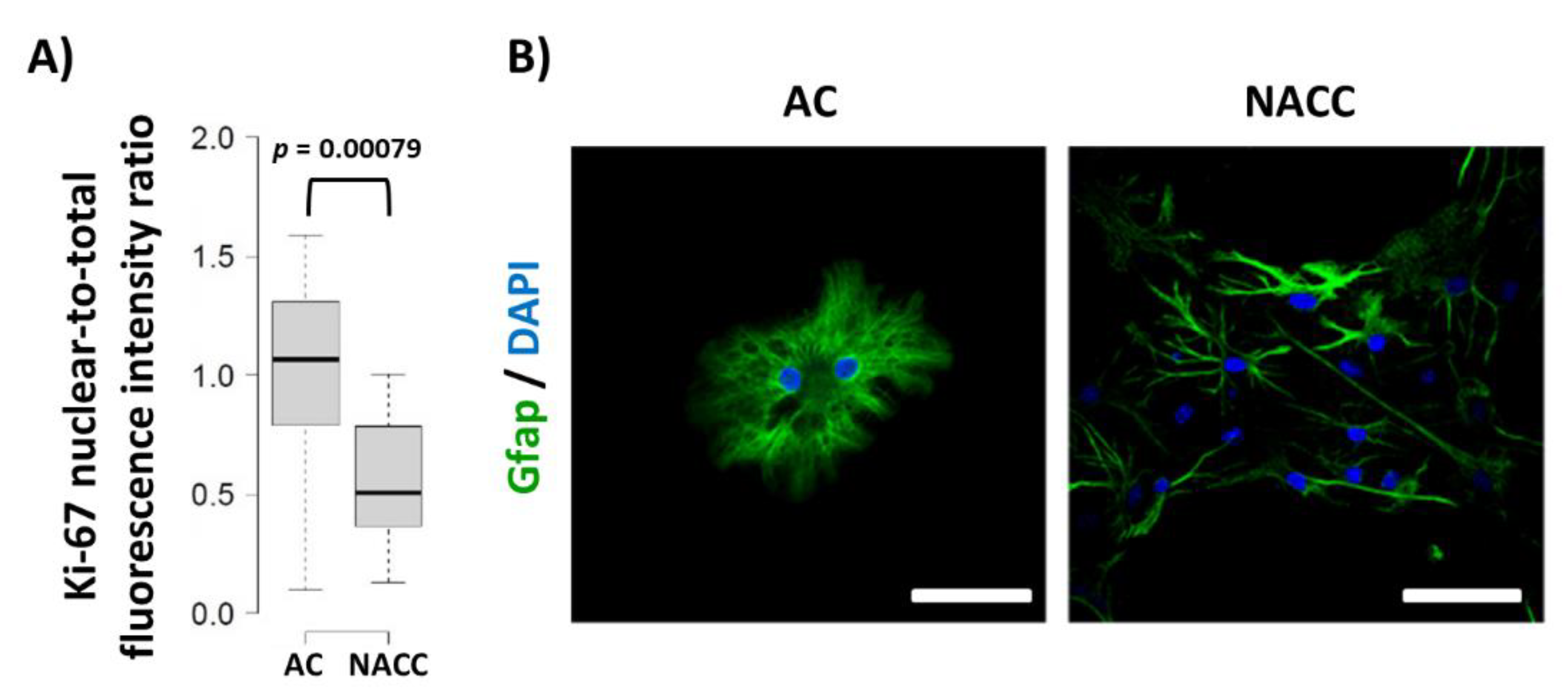
2.3. Reduction of Fbp2 Expression Is Correlated with Decrease in the Proliferative Potential of Astrocytes
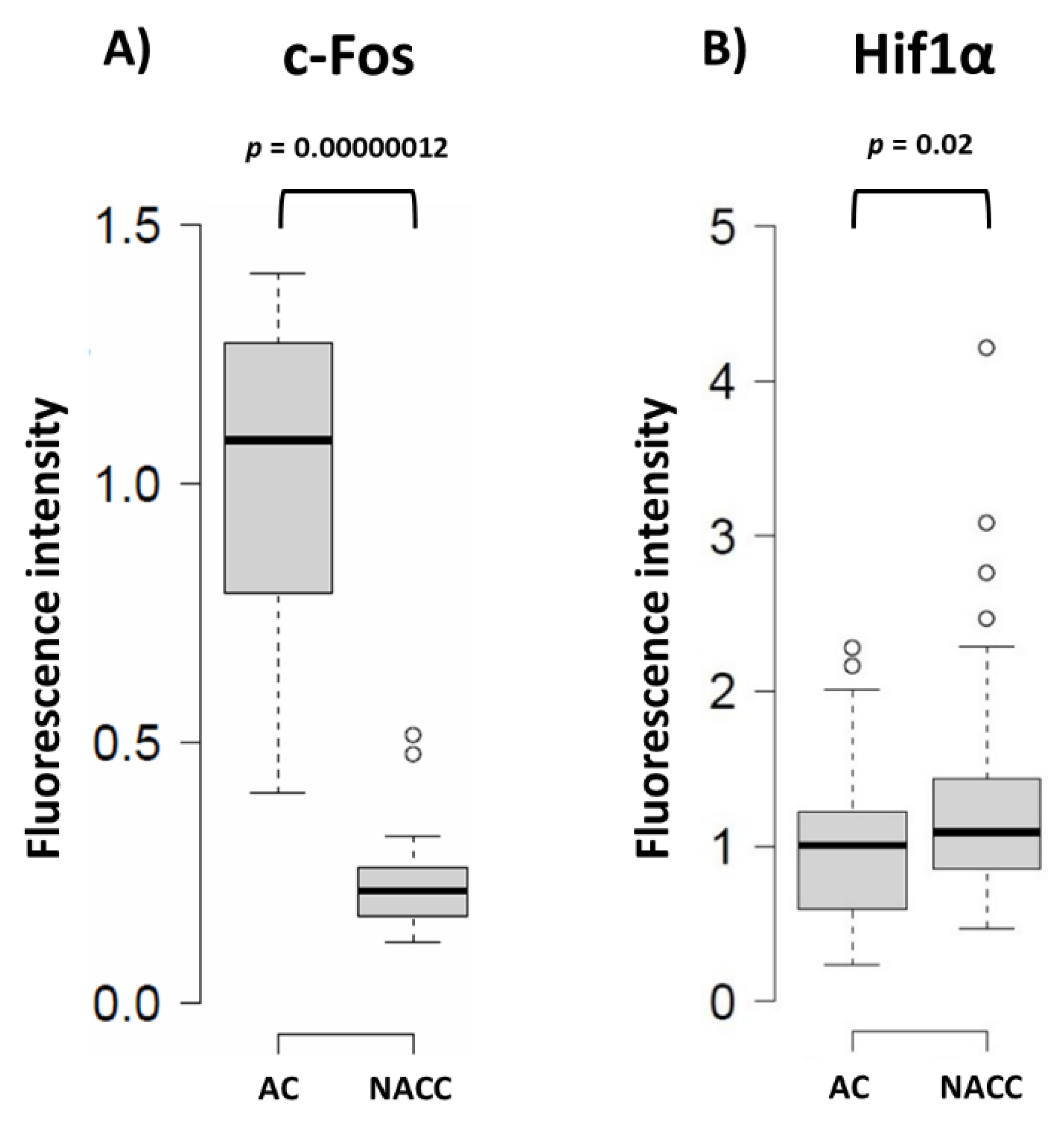
2.4. Reduction of Fbp2 Protein Level in Astrocytes Correlates with Induction of Hif1a Expression and Changes in ROS Production
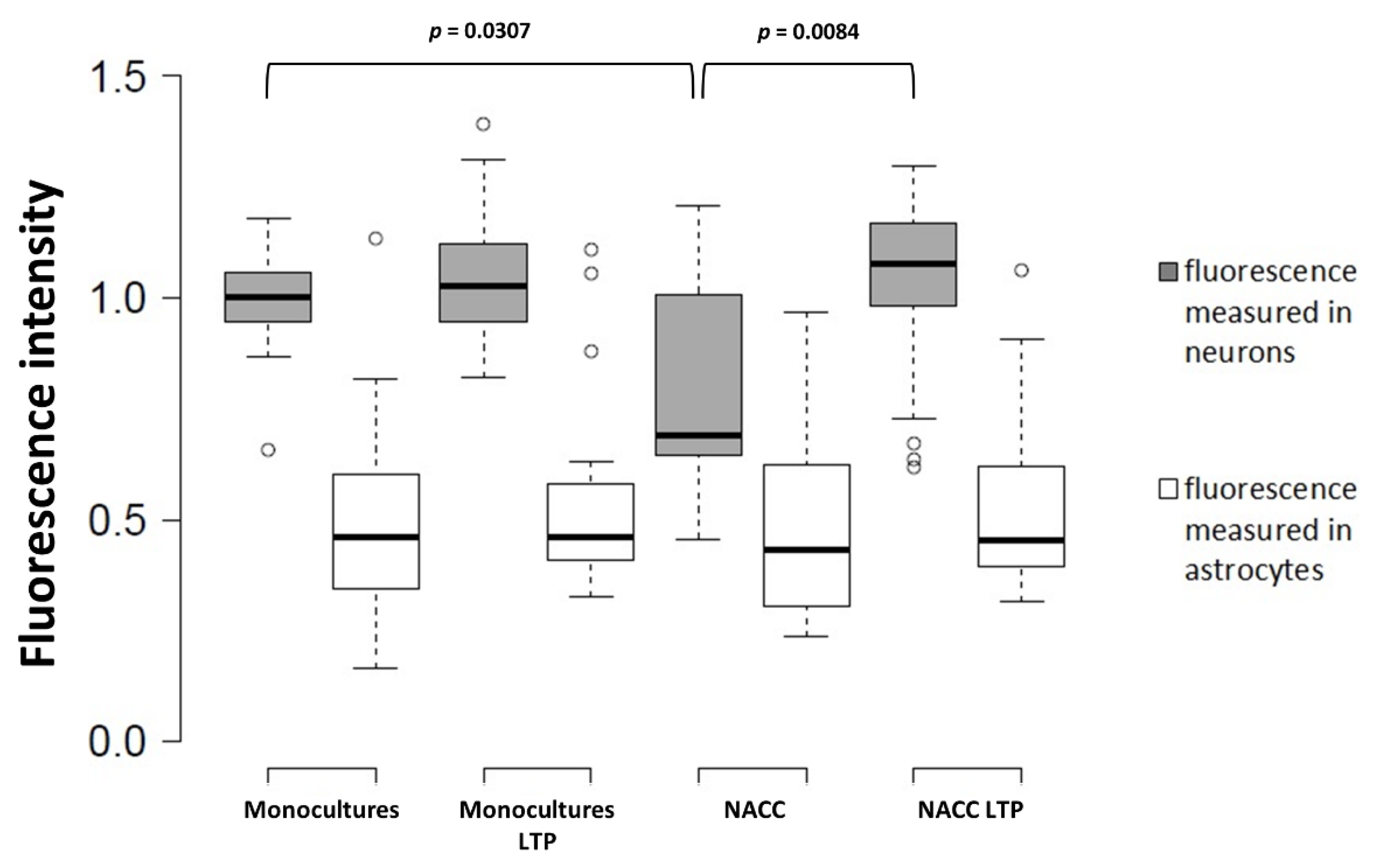
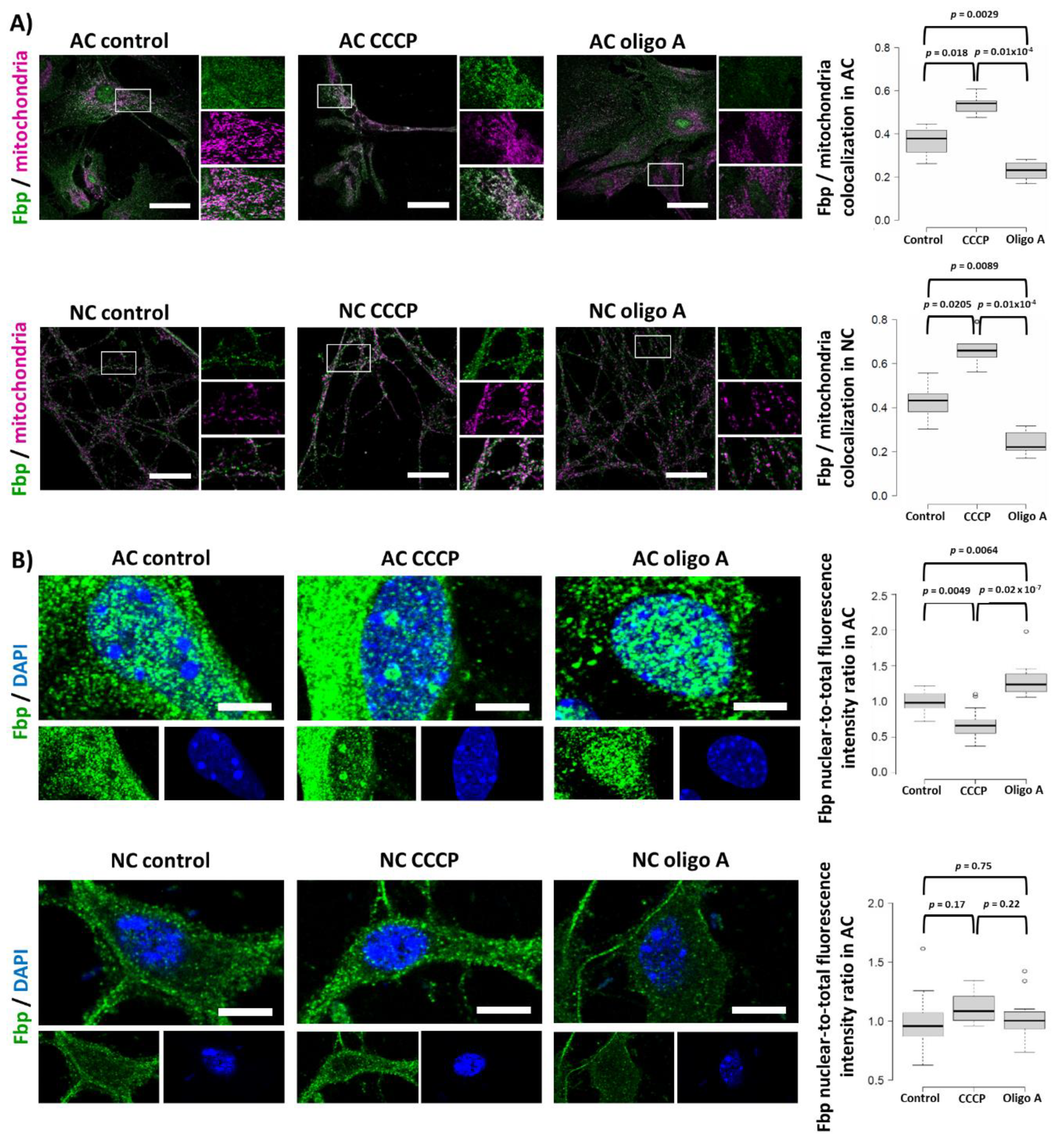
2.5. Fbp2 Is Directed to Mitochondria by a Universal, Membrane Depolarization-Related Mechanism But Its Nucleo-Cytoplasmic Shuttling Regulation Is Cell-Specific
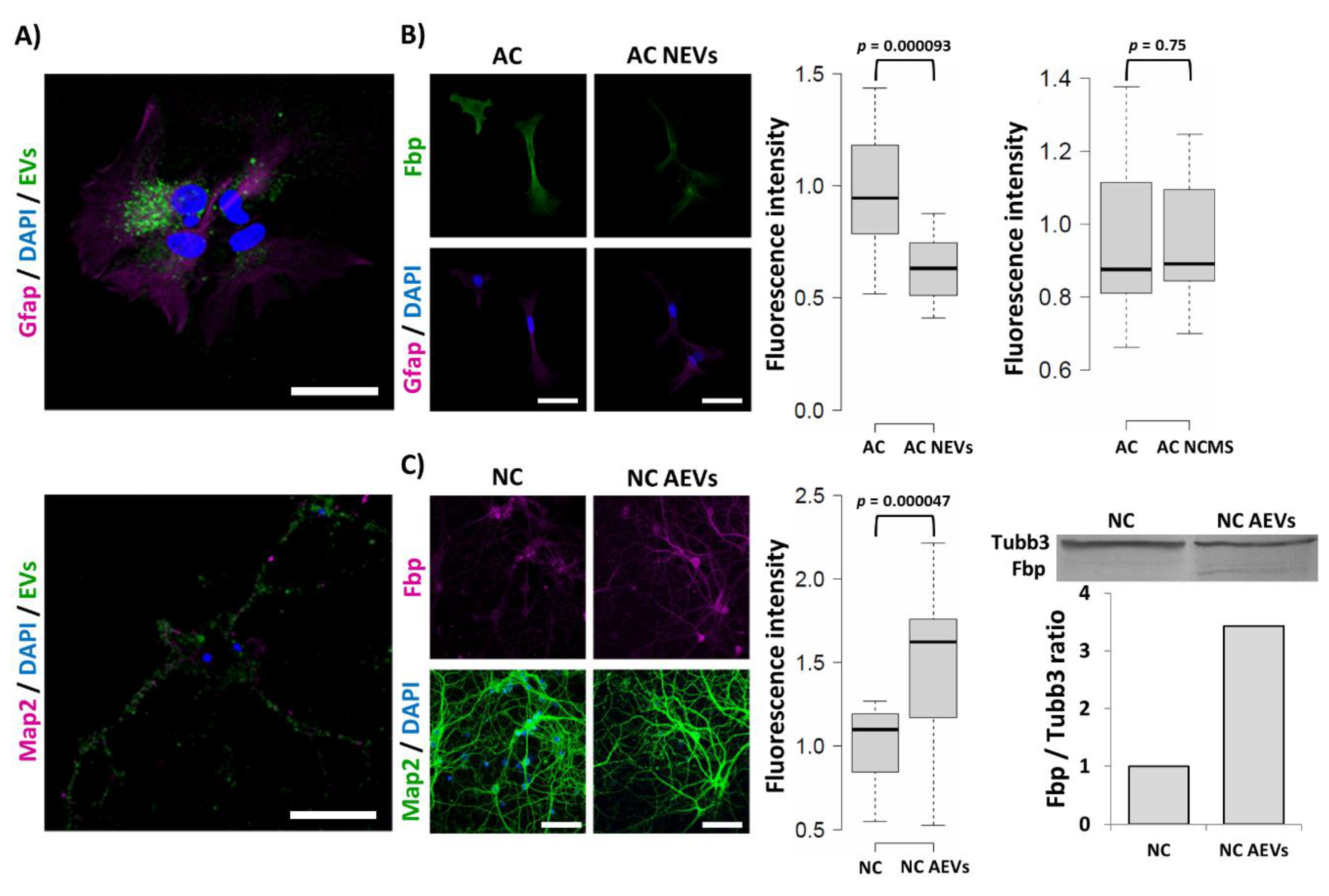
2.6. Fbp2 Protein Level in Hippocampal Cells Is Regulated by Extracellular Vesicle Cargo
3. Materials and Methods
3.1. Cell Culture and Treatment
3.2. Immunofluorescence
3.3. ROS Visualization
3.4. Fluorescence In Situ Hybridization (FISH)
3.5. Isolation and Visualization of Extracellular Vesicles
3.6. Confocal Microscopy
3.7. Fbp2 Activity Measurement
3.8. Western Blot
3.9. Statistical Analysis
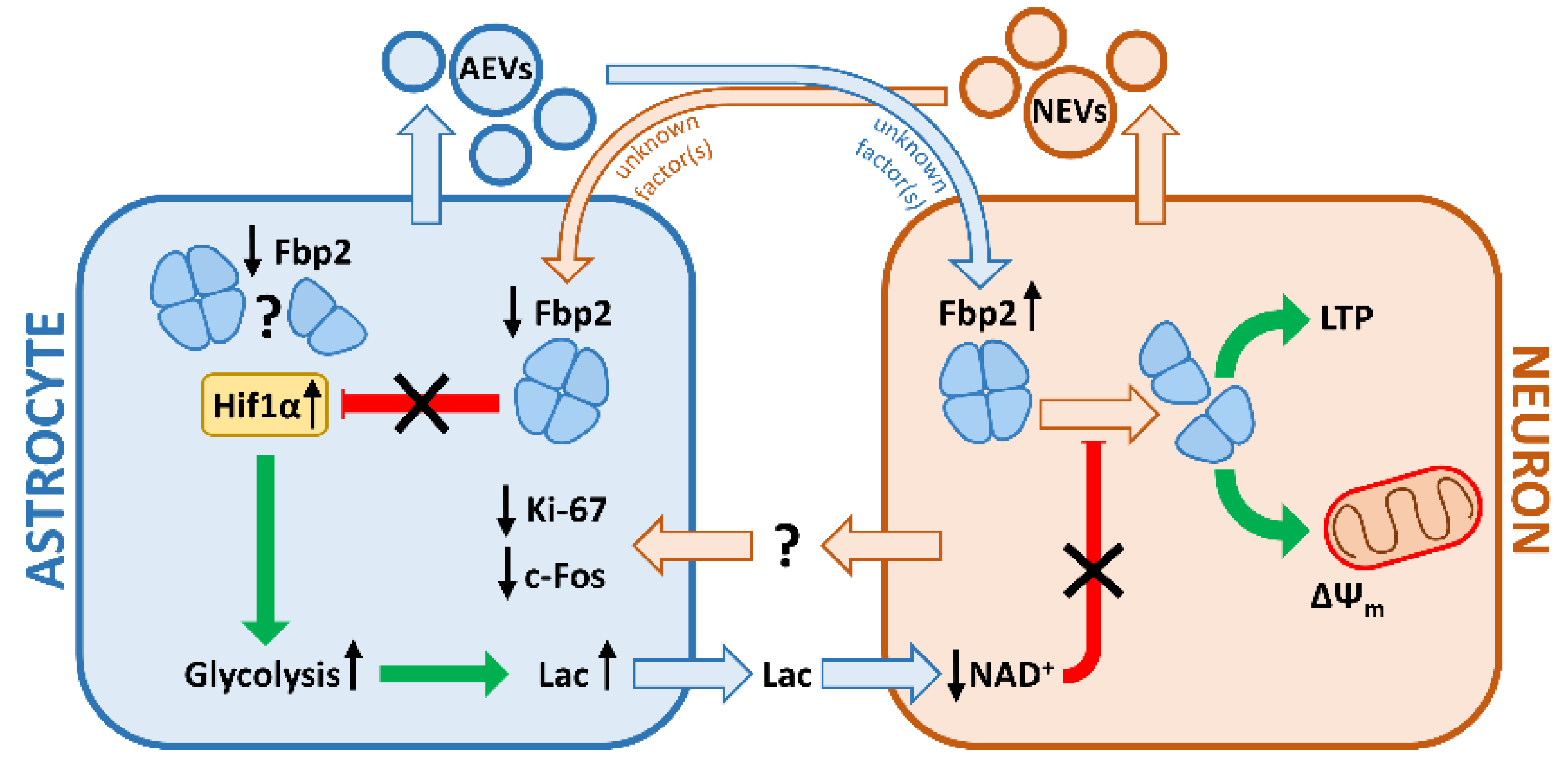
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Duda, P.; Janczara, J.; McCubrey, J.A.; Gizak, A.; Rakus, D. The Reverse Warburg Effect Is Associated with Fbp2-Dependent Hif1α Regulation in Cancer Cells Stimulated by Fibroblasts. Cells 2020, 9, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamczur, P.; Sok, A.J.; Rzechonek, A.; Rakus, D. Cell cycle-dependent expression and subcellular localization of fructose 1,6-bisphosphatase. Histochem. Cell Biol. 2012, 137, 121–136. [Google Scholar] [CrossRef] [Green Version]
- Huangyang, P.; Li, F.; Lee, P.; Nissim, I.; Weljie, A.M.; Mancuso, A.; Li, B.; Keith, B.; Yoon, S.S.; Celeste Simon, M. Fructose-1,6-Bisphosphatase 2 Inhibits Sarcoma Progression by Restraining Mitochondrial Biogenesis. Cell Metab. 2020, 31, 174–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gizak, A.; Pirog, M.; Rakus, D. Muscle FBPase binds to cardiomyocyte mitochondria under glycogen synthase kinase-3 inhibition or elevation of cellular Ca 2+ level. FEBS Lett. 2012, 586, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Wisniewski, J.; Piróg, M.; Holubowicz, R.; Dobryszycki, P.; McCubrey, J.A.; Rakus, D.; Gizak, A. Dimeric and tetrameric forms of muscle fructose-1,6- bisphosphatase play different roles in the cell. Oncotarget 2017, 8, 115420–115433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duda, P.; Wójtowicz, T.; Janczara, J.; Krowarsch, D.; Czyrek, A.; Gizak, A.; Rakus, D. Fructose 1,6-Bisphosphatase 2 Plays a Crucial Role in the Induction and Maintenance of Long-Term Potentiation. Cells 2020, 9, 1375. [Google Scholar] [CrossRef] [PubMed]
- Bliss, T.V.P.; Collingridge, G.L. A synaptic model of memory: Long-term potentiation in the hippocampus. Nature 1993, 361, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Stern, S.A.; Bozdagi, O.; Huntley, G.W.; Walker, R.H.; Magistretti, P.J.; Alberini, C.M. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell 2011, 144, 810–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Ruchti, E.; Petit, J.-M.; Jourdain, P.; Grenningloh, G.; Allaman, I.; Magistretti, P.J. Lactate promotes plasticity gene expression by potentiating NMDA signaling in neurons. Proc. Natl. Acad. Sci. USA 2014, 111, 12228–12233. [Google Scholar] [CrossRef] [Green Version]
- Mamczur, P.; Borsuk, B.; Paszko, J.; Sas, Z.; Mozrzymas, J.; Wiśniewski, J.R.; Gizak, A.; Rakus, D. Astrocyte-neuron crosstalk regulates the expression and subcellular localization of carbohydrate metabolism enzymes. Glia 2015, 63, 328–340. [Google Scholar] [CrossRef]
- Zawişlak, A.; Jakimowicz, P.; McCubrey, J.A.; Rakus, D. Neuron-derived transthyretin modulates astrocytic glycolysis in hormone-independent manner. Oncotarget 2017, 8, 106625–106638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatten, M.E. Neuronal inhibition of astroglial cell proliferation is membrane mediated. J. Cell Biol. 1987, 104, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Matsutani, S.; Yamamoto, N. Neuronal regulation of astrocyte morphology in vitro is mediated by GABAergic signaling. Glia 1997, 20, 1–9. [Google Scholar] [CrossRef]
- Hisanaga, K.; Sagar, S.M.; Hicks, K.J.; Swanson, R.A.; Sharp, F.R. c-fos proto-oncogene expression in astrocytes associated with differentiation or proliferation but not depolarization. Mol. Brain Res. 1990, 8, 69–75. [Google Scholar] [CrossRef]
- Sheng, M.; Greenberg, M.E. The regulation and function of c-fos and other immediate early genes in the nervous system. Neuron 1990, 4, 477–485. [Google Scholar] [CrossRef]
- Gizak, A.; Mccubrey, J.A.; Rakus, D. Cell-to-cell lactate shuttle operates in heart and is important in age-related heart failure. Aging (Albany NY) 2020, 12, 3388–3406. [Google Scholar] [CrossRef]
- Li, B.; Qiu, B.; Lee, D.S.M.; Walton, Z.E.; Ochocki, J.D.; Mathew, L.K.; Mancuso, A.; Gade, T.P.F.; Keith, B.; Nissim, I.; et al. Fructose-1,6-bisphosphatase opposes renal carcinoma progression. Nature 2014, 513, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Marin-Hernandez, A.; Gallardo-Perez, J.; Ralph, S.; Rodriguez-Enriquez, S.; Moreno-Sanchez, R. HIF-1α Modulates Energy Metabolism in Cancer Cells by Inducing Over-Expression of Specific Glycolytic Isoforms. Mini Rev. Med. Chem. 2009, 9, 1084–1101. [Google Scholar] [CrossRef] [Green Version]
- Hirayama, Y.; Koizumi, S. Hypoxia-independent mechanisms of HIF-1α expression in astrocytes after ischemic preconditioning. Glia 2017, 65, 523–530. [Google Scholar] [CrossRef]
- Tello, D.; Balsa, E.; Acosta-Iborra, B.; Fuertes-Yebra, E.; Elorza, A.; Ordóñez, Á.; Corral-Escariz, M.; Soro, I.; López-Bernardo, E.; Perales-Clemente, E.; et al. Induction of the mitochondrial NDUFA4L2 protein by HIF-1α decreases oxygen consumption by inhibiting complex i activity. Cell Metab. 2011, 14, 768–779. [Google Scholar] [CrossRef] [Green Version]
- Bilan, D.S.; Matlashov, M.E.; Gorokhovatsky, A.Y.; Schultz, C.; Enikolopov, G.; Belousov, V.V. Genetically encoded fluorescent indicator for imaging NAD+/NADH ratio changes in different cellular compartments. Biochim. Biophys. Acta 2014, 1840, 951–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evers, M.; Salma, N.; Osseiran, S.; Casper, M.; Birngruber, R.; Evans, C.L.; Manstein, D. Enhanced quantification of metabolic activity for individual adipocytes by label-free FLIM. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.H.; Hazell, A.S. Excitotoxic mechanisms and the role of astrocytic glutamate transporters in traumatic brain injury. Neurochem. Int. 2006, 48, 394–403. [Google Scholar] [CrossRef]
- Herrup, K.; Yang, Y. Cell cycle regulation in the postmitotic neuron: Oxymoron or new biology? Nat. Rev. Neurosci. 2007, 8, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Schiera, G.; Di Liegro, C.M.; Di Liegro, I. Cell-to-cell communication in learning and memory: From neuro-and glio-transmission to information exchange mediated by extracellular vesicles. Int. J. Mol. Sci. 2020, 21, 266. [Google Scholar] [CrossRef] [Green Version]
- Hasel, P.; Dando, O.; Jiwaji, Z.; Baxter, P.; Todd, A.C.; Heron, S.; Márkus, N.M.; McQueen, J.; Hampton, D.W.; Torvell, M.; et al. Neurons and neuronal activity control gene expression in astrocytes to regulate their development and metabolism. Nat. Commun. 2017, 8, 151332. [Google Scholar] [CrossRef] [PubMed]
- Mozrzymas, J.; Szczesny, T.; Rakus, D. The effect of glycogen phosphorolysis on basal glutaminergic transmission. Biochem. Biophys. Res. Commun. 2011, 404, 652–655. [Google Scholar] [CrossRef]
- Lu, W.Y.; Man, H.Y.; Ju, W.; Trimble, W.S.; MacDonald, J.F.; Wang, Y.T. Activation of synaptic NMDA receptors induces membrane insertion of new AMPA receptors and LTP in cultured hippocampal neurons. Neuron 2001, 29, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.Q.; Wang, S.H.; Yao, W.; Wang, J.J.; Ji, F.; Yan, J.Z.; Ren, S.Q.; Chen, Z.; Liu, S.Y.; Lu, W. Role of glycine receptors in glycine-induced LTD in hippocampal CA1 pyramidal neurons. Neuropsychopharmacology 2011, 36, 1948–1958. [Google Scholar] [CrossRef] [Green Version]
- Perry, S.W.; Norman, J.P.; Barbieri, J.; Brown, E.B.; Gelbard, H.A. Mitochondrial membrane potential probes and the proton gradient: A practical usage guide. Biotechniques 2011, 50, 98–115. [Google Scholar] [CrossRef]
- Gizak, A.; Dzugaj, A. FBPase is in the nuclei of cardiomyocytes. FEBS Lett. 2003, 539, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Waldenström, A.; Gennebäck, N.; Hellman, U.; Ronquist, G. Cardiomyocyte microvesicles contain DNA/RNA and convey biological messages to target cells. PLoS ONE 2012, 7, e34653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiśniewski, J.R.; Gizak, A.; Rakus, D. Integrating Proteomics and Enzyme Kinetics Reveals Tissue-Specific Types of the Glycolytic and Gluconeogenic Pathways. J. Proteome Res. 2015, 14, 3263–3273. [Google Scholar] [CrossRef]
- Gizak, A.; Wiśniewski, J.; Heron, P.; Mamczur, P.; Sygusch, J.; Rakus, D. Targeting a moonlighting function of aldolase induces apoptosis in cancer cells. Cell Death Dis. 2019, 10, 712. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajka, D.; Duda, P.; Wójcicka, O.; Drulis-Fajdasz, D.; Rakus, D.; Gizak, A. Expression of Fbp2, a Newly Discovered Constituent of Memory Formation Mechanisms, Is Regulated by Astrocyte–Neuron Crosstalk. Int. J. Mol. Sci. 2020, 21, 6903. https://doi.org/10.3390/ijms21186903
Hajka D, Duda P, Wójcicka O, Drulis-Fajdasz D, Rakus D, Gizak A. Expression of Fbp2, a Newly Discovered Constituent of Memory Formation Mechanisms, Is Regulated by Astrocyte–Neuron Crosstalk. International Journal of Molecular Sciences. 2020; 21(18):6903. https://doi.org/10.3390/ijms21186903
Chicago/Turabian StyleHajka, Daria, Przemysław Duda, Olga Wójcicka, Dominika Drulis-Fajdasz, Dariusz Rakus, and Agnieszka Gizak. 2020. "Expression of Fbp2, a Newly Discovered Constituent of Memory Formation Mechanisms, Is Regulated by Astrocyte–Neuron Crosstalk" International Journal of Molecular Sciences 21, no. 18: 6903. https://doi.org/10.3390/ijms21186903
APA StyleHajka, D., Duda, P., Wójcicka, O., Drulis-Fajdasz, D., Rakus, D., & Gizak, A. (2020). Expression of Fbp2, a Newly Discovered Constituent of Memory Formation Mechanisms, Is Regulated by Astrocyte–Neuron Crosstalk. International Journal of Molecular Sciences, 21(18), 6903. https://doi.org/10.3390/ijms21186903