Human Fibroblasts In Vitro Exposed to 2.45 GHz Continuous and Pulsed Wave Signals: Evaluation of Biological Effects with a Multimethodological Approach


 ,
,  ,
, 

Abstract
:1. Introduction
2. Results
2.1. Cell Cycle Analysis
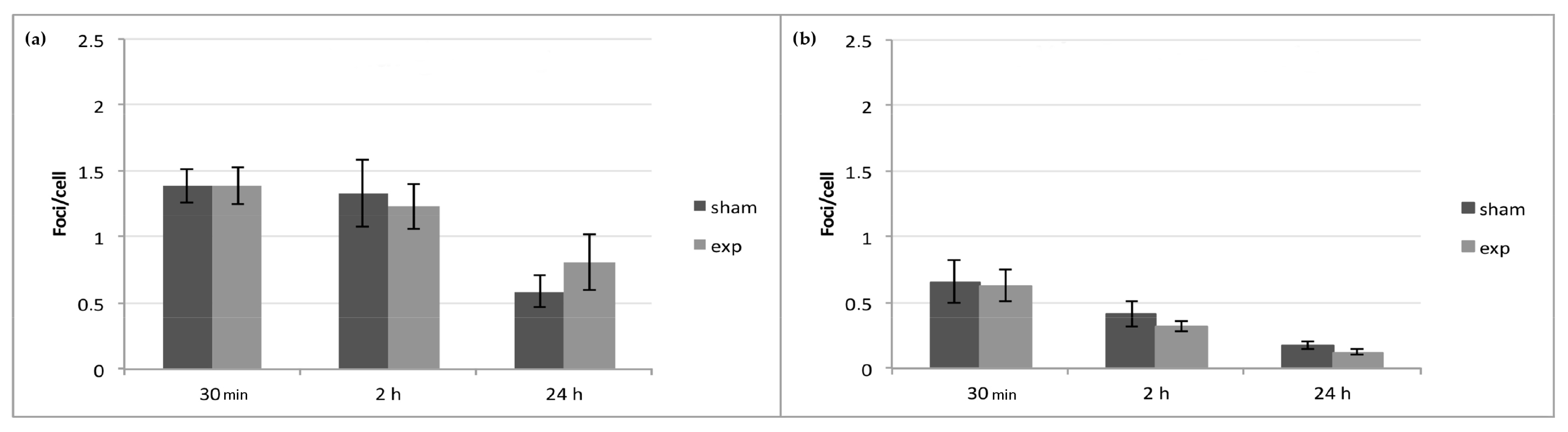
2.2. γ-H2AX/53BP1 Assay
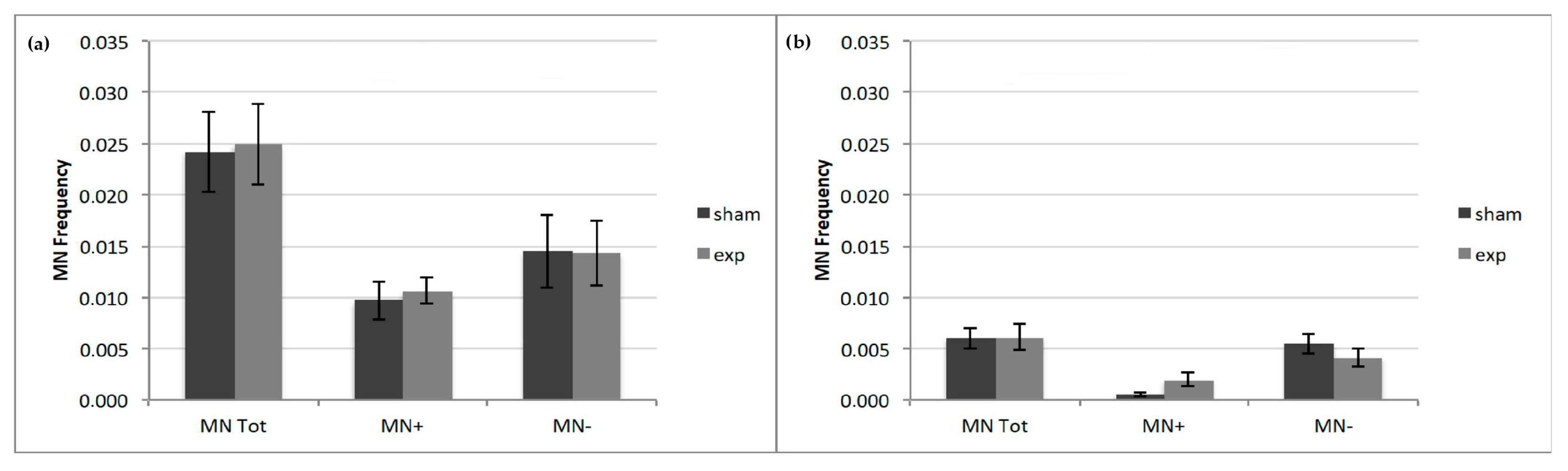
2.3. Micronuclei Anti-Kinetochore Antibody (CREST) Analysis
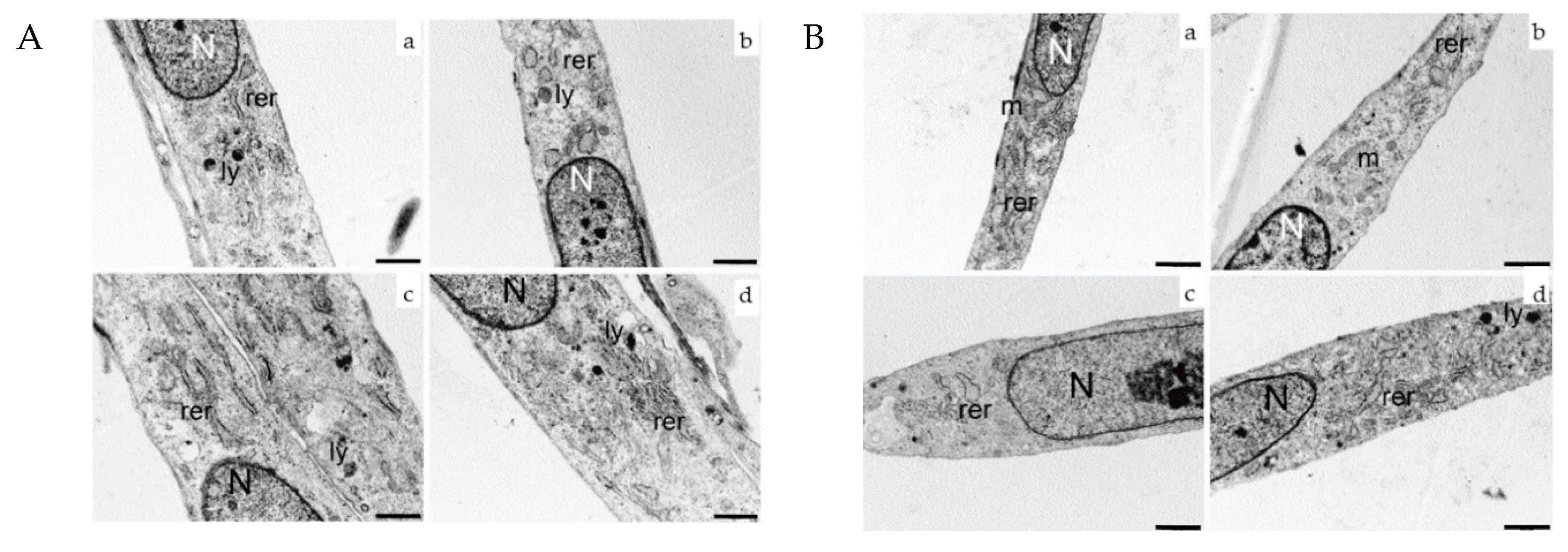
2.4. Ultrastructural Analysis
2.5. Gene Expression Profiling
2.5.1. Differential Gene Expression Analysis
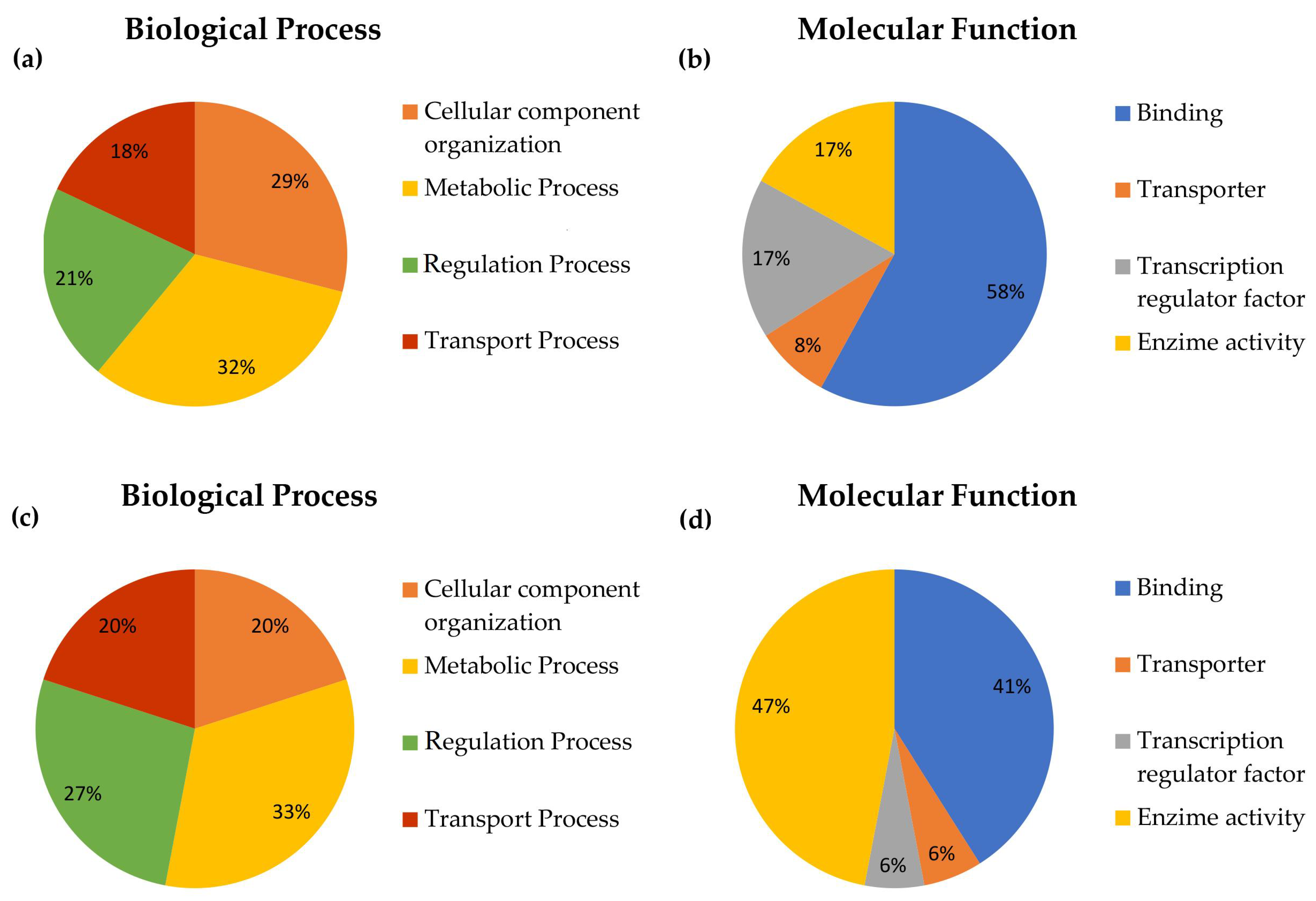
2.5.2. DGE Functional Classification
2.5.3. Quantitative RT-PCR Validation
3. Discussion
4. Materials and Methods
4.1. Exposure System and Dosimetry
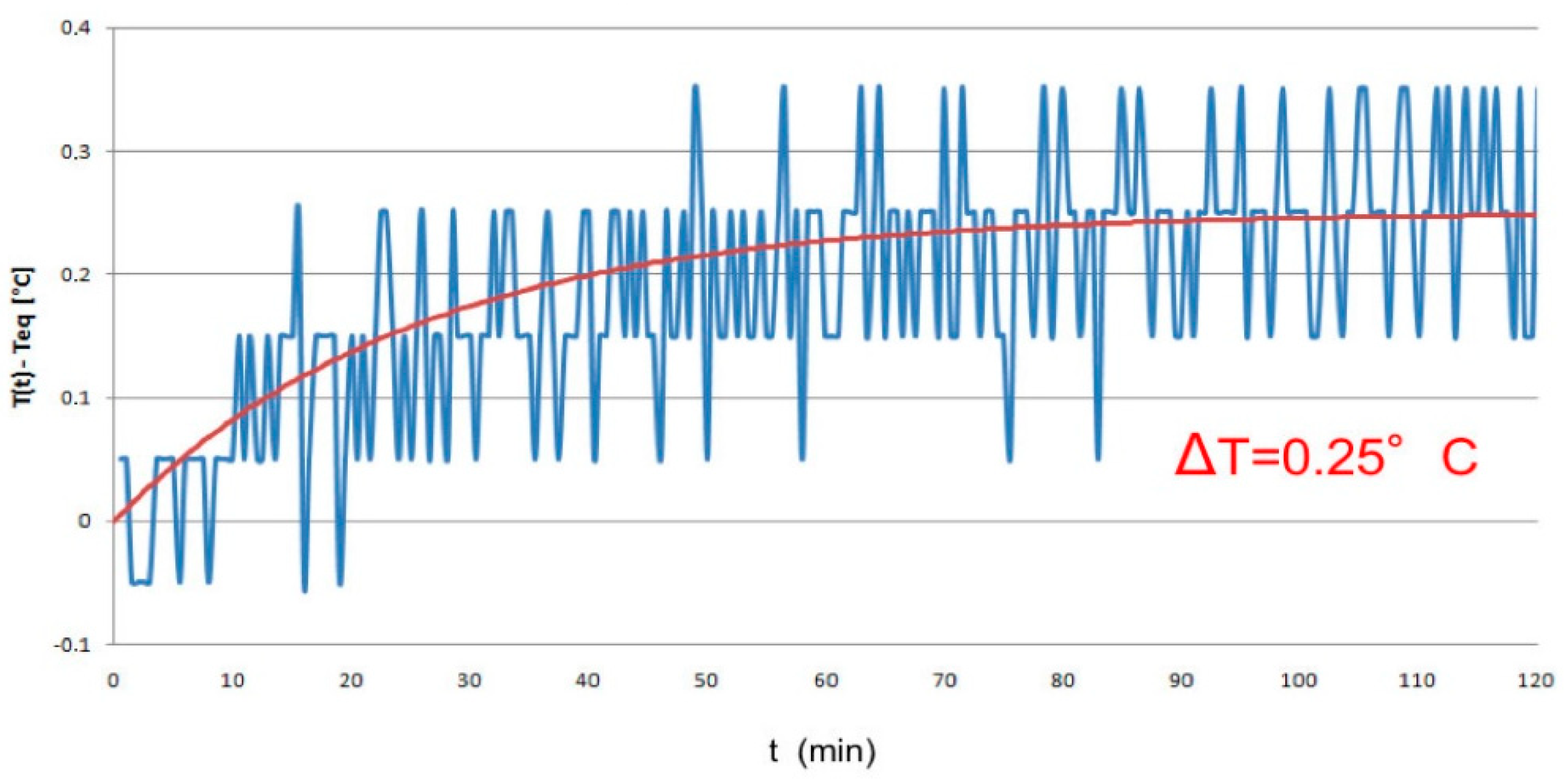
4.2. Temperature Monitoring
4.3. Cell Cultures and Exposure Protocol
4.4. Cell Cycle Analysis
4.5. γ-H2AX/53BP1 Immunofluorescence Staining
4.6. Micronuclei Anti-Kinetochore Antibody (CREST) Analysis
4.7. Statistical Analysis
4.8. Transmission Electron Microscopy
4.9. Gene Expression Profiling
4.9.1. RNA Extraction
4.9.2. mRNA Sequencing
4.9.3. Data Analysis and Statistical Methodology
4.9.4. Functional Analysis of Differentially Expressed Genes
4.9.5. Validation Experiments Using Real-Time Quantitative PCR (qRT-PCR)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- ICNIRP. Guidelines for limiting exposure to electromagnetic fields (100 kHz to 300 GHz). Health Phys. 2020. pre-print. [Google Scholar] [CrossRef]
- Repacholi, M.H. Low-level exposure to radiofrequency electromagnetic fields: Health effects and research needs. Bioelectromagnetics 1998, 19, 1–19. [Google Scholar] [CrossRef]
- Sheppard, A.R.; Swicord, M.L.; Balzano, Q. Quantitative evaluations of mechanisms of radiofrequency interactions with biological molecules and processes. Health Phys. 2008, 95, 364–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apollonio, F.; Liberti, M.; Paffi, A.; Merla, C.; Marracino, P.; Denzi, A.; Marino, C.; D’Inzeo, G. Feasibility for Microwaves Energy to Affect Biological Systems via Non-Thermal Mechanisms: A Systematic Approach. IEEE Trans. Microw. Theory Tech. 2013, 61, 5. [Google Scholar] [CrossRef]
- Juutilainen, J.; Hoyto, A.; Kumlin, T.; Naarala, J. Review of possible modulation-dependent biological effects of radiofrequency fields. Bioelectromagnetics 2011, 32, 511–534. [Google Scholar] [CrossRef]
- Guy, A.W.; Chou, C.K.; McDougall, J.A. A quarter century of in vitro research: A new look at exposure methods. Bioelectromagnetics 1999, 20, 21–39. [Google Scholar] [CrossRef]
- WHO International EMF Project. 2010 WHO Research Agenda for Radiofrequency Fields; WHO International EMF Project: Geneva, Switzerland, 2010; Available online: https://apps.who.int/iris/bitstream/handle/10665/44396/9789241599948_eng.pdf (accessed on 24 May 2020).
- Paffi, A.; Apollonio, F.; Lovisolo, G.A.; Marino, C.; Pinto, R.; Repacholi, M.; Liberti, M. Considerations for Developing a Radiofrequency Exposure System: A Review for in vitro Biological Experiments. IEEE Trans. Microw. Theory Tech. 2010, 58, 2702–2714. [Google Scholar] [CrossRef]
- Kuster, N.; Schönborn, F. Recommended minimal requirements and development guidelines for exposure setups of bio-experiments addressing the health risk concern of wireless communications. Bioelectromagnetics 2000, 21, 508–514. [Google Scholar] [CrossRef]
- Vijayalaxmi. Biological and health effects of radiofrequency fields: Good study design and quality publications. Mutat. Res. Toxicol. Environ. Mutagen. 2016, 810, 6–12. [Google Scholar] [CrossRef]
- Pikor, L.; Thu, K.; Vucic, E.; Lam, W. The detection and implication of genome instability in cancer. Cancer Metastasis Rev. 2013, 32, 341–352. [Google Scholar] [CrossRef] [Green Version]
- Non-Ionizing Radiation, Part 2: Radiofrequency Electromagnetic Fields; IARC (International Agency for Research on Cancer) Monograph on the evaluation of carcinogenic risks to humans; IARC: Lyon, France, 2013; Volume 102.
- Ruediger, H.W. Genotoxic effects of radiofrequency electromagnetic fields. Pathophysiol. 2009, 16, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Vershaeve, L.; Juutilainen, J.; Lagroye, I.; Miyakoshi, J.; Saunders, R.; de Seze, R.; Tenforde, T.; van Rongen, E.; Veyret, B.; Xu, Z. In vitro and in vivo genotoxicity of radiofrequency fields. Mutat. Res. 2010, 705, 252–268. [Google Scholar] [CrossRef] [PubMed]
- Danese, E.; Lippi, G.; Buonocore, R.; Benati, M.; Bovo, C.; Bonaguri, C.; Salvagno, G.L.; Brocco, G.; Roggenbuck, D.; Montagnana, M. Mobile phone radiofrequency exposure has no effect on DNA double strand breaks (DSB) in human lymphocytes. Ann. Transl. Med. 2017, 5, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franchini, V.; Regalbuto, E.; De Amicis, A.; De Sanctis, S.; Di Cristofaro, S.; Coluzzi, E.; Marinaccio, J.; Sgura, A.; Ceccuzzi, S.; Doria, A.; et al. Genotoxic Effects In Human Fibroblasts Exposed To Microwave Radiation. Health Phys. 2018, 115, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Markovà, E.; Hillert, L.; Malmgren, L.; Persson, B.R.R.; Belyaev, I.Y. Microwaves from GSM mobile phones affect 53BP1 and γ-H2AX foci in human lymphocytes from hypersensitive and healthy persons. Environ. Health Perspect. 2005, 113, 1172–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belyaev, I.Y.; Markovà, E.; Hillert, L.; Malmgren, L.O.; Persson, B.R. Microwaves from UMTS/GSM mobile phones induce long-lasting inhibition of 53BP1/gamma-H2AX DNA repair foci in human lymphocytes. Bioelectromagnetics 2009, 30, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M.; Morley, A.A. Measurement of micronuclei in lymphocytes. Mutat. Res. 1985, 247, 29–36. [Google Scholar]
- Maes, A.; Verschaeve, L.; Arroyo, A.; De Wagter, C.; Vercruyssen, L. In vitro cytogenetic effects of 2450 MHz waves on human peripheral blood lymphocytes. Bioelectromagnetics 1993, 14, 495–501. [Google Scholar] [CrossRef]
- D’Ambrosio, G.; Lioi, M.B.; Massa, R.; Scarfì, M.R.; Zeni, O. Genotoxic Effects of Amplitude-Modulated Microwaves on Human Lymphocytes Exposed in Vitro under Controlled Conditions. Electro Magn. 1995, 14, 157–164. [Google Scholar] [CrossRef]
- Garaj-Vrhovac, V.; Fucić, A.; Horvat, D. The correlation between the frequency of micronuclei and specific chromosome aberrations in human lymphocytes exposed to microwave radiation in vitro. Mutat. Res. 1992, 281, 181–186. [Google Scholar] [CrossRef]
- D’Ambrosio, G.; Massa, R.; Scarfì, M.R.; Zeni, O. Cytogenetic damage in human lymphocytes following GMSK phase modulated microwave exposure. Bioelectromagnetics 2002, 23, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Tice, R.R.; Hook, G.G.; Donner, M.; McRee, D.I.; Guy, A.W. Genotoxicity of radiofrequency signals. I. Investigation of DNA damage and micronuclei induction in cultured human blood cells. Bioelectromagnetics 2002, 23, 113–126. [Google Scholar] [CrossRef]
- Zotti-Martelli, L.; Peccatori, M.; Scarpato, R.; Migliore, L. Induction of micronuclei in human lymphocytes exposed in vitro to microwave radiation. Mutat. Res. 2000, 472, 51–58. [Google Scholar] [CrossRef]
- Zotti-Martelli, L.; Peccatori, M.; Maggini, V.; Ballardin, M.; Barale, R. Individual responsiveness to induction of micronuclei in human lymphocytes after exposure in vitro to 1800-MHz microwave radiation. Mutat Res. 2005, 582, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Vijayalaxmi; Frei, M.R.; Dusch, S.J.; Guel, V.; Meltz, M.L.; Jauchem, J.R. Frequency of micronuclei in the peripheral blood and bone marrow of cancer-prone mice chronically exposed to 2450 MHz radiofrequency radiation. Radiat. Res. 1998, 149, 308. [Google Scholar] [CrossRef]
- Vijayalaxmi; Pickard, W.F.; Bisht, K.S.; Prihoda, T.J.; Meltz, M.L.; LaRegina, M.C.; Roti Roti, J.L.; Straube, W.L.; Moros, E.G. Micronuclei in the peripheral blood and bone marrow cells of rats exposed to 2450 MHz radiofrequency radiation. Int. J. Radiat. Biol. 2001, 77, 1109–1115. [Google Scholar] [CrossRef]
- McNamee, J.P.; Bellier, P.V.; Gajda, G.B.; Miller, S.M.; Lemay, E.P.; Lavallée, B.F.; Marro, L.; Thansandote, A. DNA damage and micronucleus induction in human leukocytes after acute in vitro exposure to a 1.9 GHz continuous-wave radiofrequency field. Radiat. Res. 2002, 158, 523–533. [Google Scholar] [CrossRef]
- McNamee, J.P.; Bellier, P.V.; Gajda, G.B.; Lavalleè, B.E.; Lemay, E.P.; Marro, L.; Thansandotea, A. DNA Damage in Human Leukocytes after Acute In Vitro Exposure to a 1.9 GHz Pulse-Modulated Radiofrequency Field. Radiat. Res. 2003, 158, 534–537. [Google Scholar] [CrossRef]
- Zeni, O.; Schiavoni, A.S.; Sannino, A.; Antolini, A.; Forigo, D.; Bersani, F.; Scarfi, M.R. Lack of genotoxic effects (micronucleus induction) in human lymphocytes exposed in vitro to 900 MHz electromagnetic fields. Radiat. Res. 2003, 160, 152–158. [Google Scholar] [CrossRef]
- Scarfì, M.R.; Fresegna, A.M.; Villani, P.; Pinto, R.; Marino, C.; Sarti, M.; Altavista, P.; Sannino, A.; Lovisolo, G.A. Exposure to radiofrequency radiation (900 MHz, GSM signal) does not affect micronucleus frequency and cell proliferation in human peripheral blood lymphocytes: An interlaboratory study. Radiat. Res. 2006, 165, 655–663. [Google Scholar] [CrossRef]
- Speit, G.; Schutz, P.; Hoffmann, H. Genotoxic effects of exposure to radiofrequency electromagnetic fields (RFEMF) in cultured mammalian cells are not independently reproducible. Mutat. Res. 2007, 626, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, C.; Kratochvil, E.; Pilger, A.; Kuster, N.; Adlkofer, F.; Rudiger, H.W. Radiofrequency electromagnetic fields (UMTS, 1,950 MHz) induce genotoxic effects in vitro in human fibroblasts but not in lymphocytes. Int. Arch. Occup. Environ. Health 2008, 81, 755–767. [Google Scholar] [CrossRef]
- Mashevich, M.; Folkman, D.; Kesar, A.; Barbul, A.; Korenstein, R.; Jerby, E.; Avivi, L. Exposure of Human Peripheral Blood Lymphocytes to Electromagnetic Fields Associated with Cellular Phones Leads to Chromosomal Instability. Bioelectromagnetics 2003, 24, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Mazor, R.; Korenstein-Ilan, A.; Barbul, A.; Eshet, Y.; Shahadi, A.; Jerbyb, E.; Korenstein, R. Increased Levels of Numerical Chromosome Aberrations after In Vitro Exposure of Human Peripheral Blood Lymphocytes to Radiofrequency Electromagnetic Fields for 72 Hours. Radiat. Res. 2008, 169, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Manti, L.; Braselmann, H.; Calabrese, M.L.; Massa, R.; Pugliese, M.; Scampoli, P.; Sicignano, G.; Grossia, G. Effects of Modulated Microwave Radiation at Cellular Telephone Frequency (1.95 GHz) on X-Ray-Induced Chromosome Aberrations in Human Lymphocytes In Vitro. Radiat. Res. 2008, 169, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Bourthoumieu, S.; Terro, F.; Leveque, P.; Collin, A.; Joubert, V.; Yardin, C. Aneuploidy studies in human cells exposed in vitro to GSM-900 MHz radiofrequency radiation using FISH. Int. J. Radiat. Biol. 2011, 87, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Velizarov, S.; Raskmark, P.; Kwee, S. The effects of radiofrequency fields on cell proliferation are non-thermal. Bioelectrochem. Bioenerg. 1999, 48, 177–180. [Google Scholar] [CrossRef]
- Esmekaya, M.A.; Seyhan, N.; Kayhan, H.; Tuysuz, M.Z.; Kurşun, A.C.; Yağcı, M. Investigation of the effects of 2.1 GHz microwave radiation on mitochondrial membrane potential (ΔΨm), apoptotic activity and cell viability in human breast fibroblast cells. Cell Biochem. Biophys. 2013, 67, 1371–1378. [Google Scholar] [CrossRef]
- Buttiglione, M.; Roca, L.; Montemurno, E.; Vitiello, F.; Capozzi, V.; Cibelli, G. Radiofrequency radiation (900 MHz) induces Egr-1 gene expression and affects cell-cycle control in human neuroblastoma cells. J. Cell. Physiol. 2007, 213, 759–767. [Google Scholar] [CrossRef]
- Ballardin, M.; Tusa, I.; Fontana, N.; Monorchio, A.; Pelletti, C.; Rogovich, A.; Barale, R.; Scarpato, R. Non-thermal effects of 2.45 GHz microwaves on spindle assembly, mitotic cells and viability of Chinese hamster V-79 cells. Mutat. Res. 2011, 716, 1–9. [Google Scholar] [CrossRef]
- Miyakoshi, J.; Takemasa, K.; Takashima, Y.; Ding, G.R.; Hirose, H.; Koyama, S. Effects of exposure to a 1950 MHz radio frequency field on expression of Hsp70 and Hsp27 in human glioma cells. Bioelectromagnetics 2005, 26, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Merola, P.; Marino, C.; Lovisolo, G.A.; Pinto, R.; Laconi, C.; Negroni, A. Proliferation and apoptosis in a neuroblastoma cell line exposed to 900 MHz modulated radiofrequency field. Bioelectromagnetics 2006, 27, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.; Milochau, A.; Ruffie, G.; Poulletier de Gannes, F.; Lagroye, I.; Haro, E.; Surleve-Bazeille, J.E.; Billaudel, B.; Lassegues, M.; Veyret, B. Human skin cell stress response to GSM-900 mobile phone signals. FEBS J. 2006, 273, 5491–5507. [Google Scholar] [CrossRef]
- Gurisik, E.; Warton, K.; Martin, D.K.; Velenzuela, S.M. An in vitro study of the effects of exposure to a GSM signal in two human cell lines: Monocytic U937 and neuroblastoma SK-N-SH. Cell Biol. Int. 2006, 30, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Kim, B.C.; Han, N.K.; Lee, Y.S.; Kim, T.; Yun, J.H.; Kim, N.; Pack, J.K.; Lee, J.S. Effects of combined radiofrequency radiation exposure on the cell cycle and its regulatory proteins. Bioelectromagnetics 2011, 32, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Erdine, S.; Bilir, A.; Cosman, E.R.; Cosman, E.R., Jr. Ultrastructural changes in axons following exposure to pulsed radiofrequency fields. Pain Pract. 2009, 9, 407–417. [Google Scholar] [CrossRef]
- Zhao, R.; Zhang, S.; Xu, Z.; Ju, L.; Lu, D.; Yao, G. Studying gene expression profile of rat neuron exposed to 1800 MHz radiofrequency electromagnetic fields with cDNA microassay. Toxicology 2007, 235, 167–175. [Google Scholar] [CrossRef]
- Remondini, D.; Nylund, R.; Reivinen, J.; Poulletier de Gannes, F.; Veyret, B.; Lagroye, I.; Haro, E.; Trillo, A.M.; Capri, M.; Maercker, C. Gene expression changes in human cells after exposure to mobile phone microwaves. Proteomics 2006, 6, 4745–4754. [Google Scholar] [CrossRef]
- Nylund, R.; Leszczynski, D. Mobile phone radiation causes changes in gene and protein expression in human endothelial cell lines and the response seems to be genome and proteome dependent. Proteomics 2006, 6, 4769–4780. [Google Scholar] [CrossRef]
- Le Quement, C.; Nicolas Nicolaz, C.; Zhadobov, M.; Desmots, F.; Sauleau, R.; Aubry, M.; Michel, D.; Le Dréan, Y. Whole-genome expression analysis in primary human keratinocyte cell cultures exposed to 60 GHz radiation. Bioelectromagnetics 2012, 33, 147–158. [Google Scholar] [CrossRef]
- Habauzit, D.; Le Quement, C.; Zhadobov, M.; Martin, C.; Aubry, M.; Sauleau, R.; Le Drean, Y. Transcriptome Analysis Reveals the Contribution of Thermal and the Specific Effects in Cellular Response to Millimeter Wave Exposure. PLoS ONE 2014, 9, e109435. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, V.; Mariampillai, A.; Bellier, P.V.; Qutob, S.S.; Gajda, G.B.; Lemay, E.; Thansandote, A.; McNamee, J.P. Gene Expression Analysis of a Human Lymphoblastoma Cell Line Exposed In Vitro to an Intermittent 1.9 GHz Pulse-Modulated Radiofrequency Field. Radiat. Res. 2006, 165, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Qutob, S.S.; Chauhan, V.; Bellier, P.V.; Yauk, C.L.; Douglas, G.R.; Williams, A.; Berndt, L.; Gajda, G.; Thansondote, A.; McNamee, J.P. Microarray gene expression profiling of a human glioblastoma cell line exposed in vitro to 1.9 GHz pulse modulated radiofrequency fields. Radiat. Res. 2006, 165, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Kiyokawa, T.; Narita, E.; Suzuki, Y.; Taki, M.; Miyakoshi, J. Analysis of gene expression in a human-derived glial cell line exposed to 2.45 GHz continuous radiofrequency electromagnetic fields. J. Radiat. Res. 2011, 52, 185–192. [Google Scholar] [CrossRef]
- Whitehead, T.D.; Moros, E.G.; Brownstein, B.H.; Roti Roti, J.L. Gene expression does not change significantly in C3H 10TK cells after exposure to 847.74 CDMA or 835.62 FDMA radiofrequency radiation. Radiat. Res. 2006, 165, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Hirose, H.; Sakuma, N.; Kaji, N.; Nakayama, K.; Inoue, K.; Sekijima, M.; Nojima, T.; Miyakoshi, J. Mobile phone base station-emitted radiation does not induce phosphorylation of hsp27. Bioelectromagnetics 2007, 28, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Chen, G.; Weng, Y.; Wang, L.; Chiang, H.; Lu, D.; Xu, Z. Effects of global system for mobile communications 1800 MHz radiofrequency electromagnetic fields on gene and protein expression in MCF-7 cells. Proteomics 2006, 6, 4732–4738. [Google Scholar] [CrossRef] [PubMed]
- Dasdag, S.; Akdag, M.Z.; Erdal, M.E.; Erdal, N.; Ay, O.I.; Ay, M.E.; Yilmaz, S.G.; Tasdelen, B.; Yegin, K. Effects of 2.4 GHz Radiofrequency Radiation emitted from WI-FI equipment on microRNA expression in brain tissue. Int. J. Radiat. Biol. 2015, 91, 555–561. [Google Scholar] [CrossRef]
- McNamee, J.P.; Bellier, P.V.; Konkle, A.T.M.; Thomas, R.; Siiwat Wasoontarajaroen, S.; Lemay, E.; Gajda, G.B. Analysis of gene expression in mouse brain regions after exposure to 1.9 GHz radiofrequency fields. Int. J. Radiat. Biol. 2016, 92, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lu, Y.; Yi, J.; Li, Z.; Gao, D.; Yu, Z.; Wu, T.; Zhang, C. A Genome-Wide mRNA Expression Profile in Caenorhabditis elegans under Prolonged Exposure to 1750 MHz Radiofrequency Fields. PLoS ONE 2016, 11, e0147273. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.W.; Yang, C.J.; Lian, H.Y.; Cai, P. Exposure of ELF-EMF and RF-EMF Increase the Rate of Glucose Transport and TCA Cycle in Budding Yeast. Front. Microbiol. 2016, 7, 1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Said-Salman, I.H.; Jebaii, F.A.; Yusef, H.H.; Moustafa, M.E. Global gene expression analysis of Escherichia coli K-12 DH5α after exposure to 2.4 GHz wireless fidelity radiation. Sci. Rep. 2019, 9, 14425. [Google Scholar] [CrossRef] [PubMed]
- Wireless LAN Medium Access Control (MAC) and Physical Layer (PHY) Specification: Higher-Speed Physical Layer Extension to 54 Mbps; IEEE Std. 802.11g; ISEE: Washington, DC, USA, 2003.
- Vijayalaxmi; Obe, G. Controversial cytogenetic observations in mammalian somatic cells exposed to radiofrequency radiation. Radiat. Res. 2004, 162, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Simkó, M.; Remondini, D.; Zeni, O.; Scarfi, M.R. Quality Matters: Systematic Analysis of Endpoints Related to “Cellular Life” in Vitro Data of Radiofrequency Electromagnetic Field Exposure. Int. J. Environ. Res. Public Health 2016, 13, 701. [Google Scholar] [CrossRef] [PubMed]
- Hansteen, I.L.; Clausen, K.O.; Haugan, V.; Svendsen, M.; Svendsen, M.V.; Eriksen, J.G.; Skiaker, R.; Hauger, E.; Lågeide, L.; Vistnes, A.I.; et al. Cytogenetic Effects of Exposure to 2.3 GHz Radiofrequency Radiation on Human Lymphocytes In Vitro. Anticancer Res. 2009, 29, 4323–4330. [Google Scholar]
- Takashima, Y.; Hirose, H.; Koyama, S.; Suzuki, Y.; Taki, M.; Miyakoshi, J. Effects of continuous and intermittent exposure to RF fields with a wide range of SARs on cell growth, survival, and cell cycle distribution. Bioelectromagnetics 2006, 27, 392–400. [Google Scholar] [CrossRef]
- Sekijima, M.; Takeda, H.; Yasunaga, K.; Sakuma, N.; Hirose, H.; Nojima, T.; Miyakoshi, J. 2-GHz band CW and W-CDMA modulated radiofrequency fields have no significant effect on cell proliferation and gene expression profile in human cells. J. Radiat. Res. 2010, 51, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Koyama, S.; Isozumi, Y.; Suzuki, Y.; Taki, M.; Miyakoshi, J. Effects of 2.45-GHz Electromagnetic Fields with a Wide Range of SARs on Micronucleus Formation in CHO-K1 Cells. Sci. World J. 2004, 4, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, A.; Alves, R.; Ramalho, A. Cytogenetic analysis of the effects of 2.5 and 10.5 GHz microwaves on human lymphocytes. Genet. Mol. Biol. 2004, 27, 460–466. [Google Scholar] [CrossRef] [Green Version]
- Vijayalaxmi. Cytogenetic Studies in Human Blood Lymphocytes Exposed In Vitro to 2.45 GHz or 8.2 GHz Radiofrequency Radiation. Radiat. Res. 2006, 166, 532–538. [Google Scholar] [CrossRef]
- Zeni, O.; Schiavoni, A.; Perrotta, A.; Forigo, D.; Deplano, M.; Scarfi, M.R. Evaluation of genotoxic effects in human leukocytes after in vitro exposure to 1950 MHz UMTS radiofrequency field. Bioelectromagnetics 2008, 29, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, N.; Komatsubara, Y.; Takeda, H. DNA strand breaks are not induced in human cells exposed to 2.1425 GHz band CW and W-CDMA modulated radiofrequency fields allocated to mobile radio base stations. Bioelectromagnetics 2006, 27, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, G.; Fu, X.; Xue, J.; Ji, S.; Zhang, Z.; Zhang, Y.; Li, A. Exposure to 3G mobile phone signals does not affect the biological features of brain tumor cells. BMC Public Health 2015, 15, 764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamkowski, A.; Kreitlow, M.; Radunz, J.; Willenbockel, M.; Sabath, F.; Schuhn, W.; Stiemer, M.; Fichte, L.O.; Dudzinski, M.; Böhmelt, S.; et al. Gene Expression Analysis in Human Peripheral Blood Cells after 900 MHz RF-EMF Short-Term Exposure. Radiat. Res. 2018, 189, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Hermanns, P.; Reicherter, K.; Lee, B. RMRP (RNA component of mitochondrial RNA processing endoribonuclease). Atlas Genet. Cytogenet. Oncol. Haematol. 2008, 12, 328–333. [Google Scholar] [CrossRef]
- Maida, Y.; Mami, Y.; Furuuchi, M.; Lassmann, T.; Possemato, R.; Okamoto, N.; Kasim, V.; Hayashizaki, Y.; Hahn, W.C.; Masutomi, K. An RNA dependent RNA polymerase formed by hTERT and the RNase MRP RNA. Nature 2009, 461, 230–235. [Google Scholar] [CrossRef]
- Davis, T.A.; Loos, B.; Engelbrecht, A.M. AHNAK: The giant jack of all trades. Cell. Signal. 2014, 26, 2683–2693. [Google Scholar] [CrossRef]
- Marjanović, A.M.; Pavičić, I.; Trošić, I. Biological Indicators in Response to Radiofrequency/Microwave Exposure. Arh. Hig. Rada. Toksikol. 2012, 63, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Z.; Jiang, Y.; Wang, L.; Wang, L.; Jiang, J.; Zhang, J. Mutations in KIAA1109, CACNA1C, BSN, AKAP13, CELSR2, and HELZ2 Are Associated with the Prognosis in Endometrial Cancer. Front. Genet. 2019, 10, 909. [Google Scholar] [CrossRef]
- Veltri, A.; Lang, C.; Lien, W.H. Concise Review: Wnt Signaling Pathways in Skin Development and Epidermal Stem Cells. Stem Cells 2018, 36, 22–35. [Google Scholar] [CrossRef] [Green Version]
- Klaus, A.; Birchmeier, W. Wnt signalling and its impact on development and cancer. Nat. Rev. Cancer 2008, 8, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Komiya, Y.; Habas, R. Wnt signal transduction pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durna Dastan, S.; Soylu, S.; Pence, H.H.; Uyanik, B.; Duman, M.; Kurt, V.; Dastan, T.; Zilan, V.; Turan, V. Hazardous Genomic Bioeffects of Home Wi-Fi Systems. NeuroQuantology 2018, 16. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Johnson, D.; Dunbar, K.; Dong, H.; Ge, X.; Kim, Y.C.; Wing, C.; Jayathilaka, N.; Emmanuel, N.; Zhou, C.Q. 2.45 GHz radiofrequency fields alter gene expression in cultured human cells. FEBS Lett. 2005, 579, 4829–4836. [Google Scholar] [CrossRef] [Green Version]
- Pacini, S.; Ruggiero, M.; Sardi, I.; Aterini, S.; Gulisano, F.; Gulisano, M. Exposure to global system for mobile communication (GSM) cellular phone radiofrequency alters gene expression, proliferation, and morphology of human skin fibroblasts. Oncol. Res. 2002, 13, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Paffi, A.; Liberti, M.; Lopresto, V.; Merla, C.; Lodato, R.; Lovisolo, G.A.; Apollonio, F. A Wire Patch Cell Exposure System for in vitro Experiments at Wi-Fi Frequencies. IEEE Trans. MTT 2010, 58, 4086–4093. [Google Scholar] [CrossRef]
- Molfetta, S.; Apollonio, F.; Liberti, M.; D’Inzeo, G. Low cost exposure set-up for RF biological experiments. In Abstract Book 6th EBEA; EBEA: Budapest, Hungary, 2003. [Google Scholar]
- Platano, D.; Mesirca, P.; Paffi, A.; Pellegrino, M.; Liberti, M.; Apollonio, F.; Bersani, F.; Aicardi, V. Acute exposure to 900 MHz CW and GSM-modulated radiofrequencies does not affect Ba2+ currents through voltage-gated calcium channels in rat cortical neurons. Bioelectromagnetics 2007, 28, 598–606. [Google Scholar] [CrossRef]
- Benvenuto, M.; Mattera, R.; Sticca, J.I.; Rossi, P.; Cipriani, C.; Giganti, M.G.; Volpi, A.; Modesti, A.; Masuelli, L.; Bei, R. Effect of the BH3 Mimetic Polyphenol (−)-Gossypol (AT-101) on the in vitro and in vivo Growth of Malignant Mesothelioma. Front. Pharmacol. 2018, 9, 1269. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, A.E.; Jones, T.R.; Lamprecht, M.R.; Clarke, C.; Kang, I.H.; Friman, O.; Guertin, D.A.; Chang, J.H.; Lindquist, R.A.; Moffat, J.; et al. Cellprofiler: Image analysis software for identifying and quantifying cell phenotypes. Genome Biol. 2006, 7, R100. [Google Scholar] [CrossRef] [Green Version]
- Masuelli, L.; Granato, M.; Benvenuto, M.; Mattera, R.; Bernardini, R.; Mattei, M.; d’Amati, G.; D’Orazi, G.; Faggioni, A.; Bei, R.; et al. Chloroquine supplementation increases the cytotoxic effect of curcumin against Her2/neu overexpressing breast cancer cells in vitro and in vivo in nude mice while counteracts it in immune competent mice. Oncoimmunology 2017, 6, e1356151. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.D.; Campbell, M.J.; Kejariwal, A.; Mi, H.; Karlak, B.; Daverman, R.; Diemer, K.; Muruganujan, A.; Narechania, A. PANTHER: A library of protein families and subfamilies indexed by function. Genome Res. 2003, 13, 2129–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2.45 GHz | Time Point | Sample | Sub-G1 1 | G0/G1 | S | G2/M | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | p | Mean | p | Mean | p | Mean | P | |||
| CW | 2 h | Sham | 1.92 ± 0.42 | 50.92 ± 12.97 | 6.85 ± 1.77 | 40.61 ± 11.66 | ||||
| Exposed | 1.44 ± 0.76 | NS | 50.12 ± 12.95 | NS | 7.97 ± 2.18 | NS | 40.81 ± 10.13 | NS | ||
| 24 h | Sham | 0.80 ± 0.66 | 86.03 ± 2.33 | 3.46 ± 0.49 | 9.85 ± 2.35 | |||||
| Exposed | 0.82 ± 0.70 | NS | 86.31 ± 2.61 | NS | 3.53 ± 0.59 | NS | 9.50 ± 2.81 | NS | ||
| PW | 2 h | Sham | 0.71 ± 0.13 | 72.84 ± 1.12 | 4.06 ± 0.61 | 22.62 ± 1.58 | ||||
| Exposed | 0.84 ± 0.13 | NS | 75.00 ± 0.97 | NS | 4.17 ± 0.51 | NS | 20.23 ± 1.43 | NS | ||
| 24 h | Sham | 0.49 ± 0.16 | 84.10 ± 0.54 | 2.00 ± 0.17 | 13.53 ± 0.41 | |||||
| Exposed | 0.44 ± 0.18 | NS | 82.43 ± 1.47 | NS | 2.24 ± 0.34 | NS | 15.03 ± 1.02 | NS | ||
| 2.45 GHz CW | Gene ID | Gene Name | Description | FC | Regulation |
|---|---|---|---|---|---|
| 0 min after exposure | ACOT4 | Acyl-coenzyme A thioesterase 4 | Signaling receptor binding and palmitoyl-CoA hydrolase activity | 1.6 | up |
| DLX5 | Homeobox protein DLX-5 | Transcriptional factor involved in bone development | 2 | up | |
| FAM72D | Protein FAM72D | Unknown function | 2 | up | |
| LRP2BP | LRP2-binding protein | Protein binding | 2.5 | down | |
| NIPSNAP3B | Protein NipSnap homolog 3B | Rutative roles in vesicular trafficking | 2.2 | up | |
| SUN3 | SUN domain-containing protein 3 | Protein binding | 2.3 | up | |
| 2 h after exposure | HES4 | Transcription factor HES-4 | Basic helix-loop-helix transcription factor | 1.6 | down |
| HMCN1 | Hemicentin-1 | Receptor binding | 1.8 | up | |
| ALMS1 | Alstrom syndrome protein 1 | Cytoskeletal protein binding | 1.5 | up | |
| ANKRD36C | Ankyrin repeat domain-containing protein 36 | Protein Coding gene involved in ion channel inhibitor activity | 1.6 | down | |
| GRIP2 | Glutamate receptor-interacting protein 2 | Multi-PDZ domain scaffolding proteins required for dendrite development | 1.5 | down | |
| BSN | Protein bassoon | Scaffolding protein involved in organizing the presynaptic cytoskeleton | 1.6 | up | |
| FAM53A | Protein FAM53A | Encodes a secreted peptide hormone and member of the EGF family of proteins | 1.5 | down | |
| EREG | Epiregulina | Transient receptor potential cation channel | 1.5 | down | |
| SLC9B1 | Sodium/hydrogen exchanger 9B1 | Transmembrane transporter activity | 1.7 | down | |
| LUCAT1 | lung cancer associated transcript 1 | Non-coding RNA | 1.6 | down | |
| EGR1 | Early growth response protein 1 | Transcriptional regulator | 1.7 | up | |
| AC005618.6 | Protocadherin gamma-B3 | Cell adhesion, cell-cell signaling | 1.8 | up | |
| HIST1H2AD | Histone H2A type 1-D | Histone | 2 | down | |
| HIST1H2BG | Histone H2B type 1-C/E/F/G/I | Histone | 1.8 | down | |
| IER3 | Radiation-inducible immediate-early gene IEX-1 | Cell proliferation and survival | 2.2 | down | |
| PRSS35 | Inactive serine protease 35 | Serin protease activity | 1.7 | down | |
| SAMD3 | Sterile alpha motif domain-containing protein 3 | Protein binding | 1.8 | up | |
| GPER1 | G-protein coupled estrogen receptor 1 | Protein binding | 1.5 | down | |
| RIMS2 | Regulating synaptic membrane exocytosis protein 2 | Protein binding | 1.6 | up | |
| ZNF462 | Zinc finger protein 462 | Protein binding | 1.7 | up | |
| SYNPO2L | Synaptopodin 2-like protein | Protein binding | 1.8 | up | |
| CNNM1 | Metal transporter CNNM1 | Protein binding | 1.5 | down | |
| C11orf96 | Uncharacterized protein C11orf96 | 1.7 | down | ||
| BEST1 | Bestrophin-1 | Ion binding | 1.6 | up | |
| STYK1 | Tyrosine-protein kinase STYK1 | Receptor binding | 1.5 | up | |
| RPPH1 | Ribonuclease P RNA Component H1 | long non-coding RNA | 3.9 | down | |
| RN7SL1 | RNA, 7SL, cytoplasmic 1 | small cytoplasmic RNA | 2.7 | down | |
| RN7SL2 | RN7SL2 | small cytoplasmic RNA | 4.1 | down | |
| RHOJ | Rho-related GTP-binding protein RhoJ | small GTPase | 1.5 | down | |
| SYNE2 | Nesprin-2 | Actin binding | 2 | up | |
| AHNAK2 | Protein AHNAK2 | RNA binding | 1.5 | up | |
| FBXL22 | F-box and leucine-rich protein 22 | Protein ubiquitination | 1.6 | up | |
| SLC43A2 | Large neutral amino acids transporter small subunit 4 | Transmembrane transporter activity | 1.7 | up | |
| MYCBPAP | MYCBP-associated protein | Cell differenziation | 2 | up | |
| ZNF433 | Zinc finger protein 433 | DNA-binding (transcription) | 1.3 | up | |
| ZNF233 | Zinc finger protein 233 | Nucleic acid binding, regulation of transcription | 1.8 | up | |
| RP1-198K11.5 | Non-coding RNA | 1.5 | down | ||
| MXRA5 | Matrix-remodeling-associated protein 5 | Receptor binding | 1.8 | up | |
| RMRP | RNA Component Of Mitochondrial RNA Processing Endoribonuclease | Non-coding RNA | 8.8 | down | |
| 24 h after exposure | ANKRD36C | Ankyrin repeat domain-containing protein 36 | Protein Coding gene, ion channel inhibitor activity | 1.6 | down |
| MIR145 | microRNA 145 | Non-coding RNA | 1.4 | up | |
| KIAA0895 | Uncharacterized protein KIAA0895 | 1.8 | up | ||
| LCNL1 | Lipocalin-like 1 protein | Binding, isomerase activity | 2.3 | up | |
| RASGEF1A | Ras-GEF domain-containing family member 1A | protein binding, small GTPase regulator activity | 1.5 | up | |
| RN7SL1 | RNA, 7SL, cytoplasmic 1 | small cytoplasmic RNA | 1.8 | down | |
| RN7SL2 | RNA, 7SL, cytoplasmic 2 | small cytoplasmic RNA | 1.8 | down | |
| RGS11 | Regulator of G-protein signaling 11 | Regulator of G protein signaling | 1.7 | up |
| 2.45 GHz PW | Gene ID | Gene Name | Description | FC | Regulation |
|---|---|---|---|---|---|
| 0 min after exposure | KIAA1324 | UPF0577 protein KIAA1324 | RNA binding | 1.6 | up |
| KIAA1211 | Uncharacterized protein KIAA1211 | Unknown function | 2.3 | up | |
| CXCL3 | C-X-C motif chemokine 3 | Chemokine | 1.6 | down | |
| EGR3 | Early growth response protein 3 | Transcriptional regulator | 1.5 | down | |
| SLC16A13 | Monocarboxylate transporter 13 | Transmembrane transporter | 1.5 | up | |
| 2 h after exposure | TMEM240 | Transmembrane protein 240 | Transmembrane-domain containing protein | 1.54 | up |
| TNFRSF25 | Tumor necrosis factor receptor superfamily member 25 | Signaling receptor activity | 1.7 | up | |
| BEST4 | Bestrophin-4 | Anion channel | 1.8 | up | |
| RNF175 | RING finger protein 175 | Ubiquitin- protein ligase | 1.7 | up | |
| KLKB1 | Plasma kallikrein | Serin-protease | 1.5 | down | |
| PTGER4 | Prostaglandin E2 receptor EP4 subtype | G-protein coupled receptor | 2 | down | |
| MDFI | MyoD family inhibitor | Transcription factor binding | 2 | up | |
| PPP1R9A | Neurabin-1 | Actin binding | 1.9 | down | |
| RMRP | RNA Component Of Mitochondrial RNA Processing Endoribonuclease | Non-coding RNA | 5.7 | down | |
| PRUNE2 | Protein prune homolog 2 | Pyrophosphatase activity | 1.6 | down | |
| ENO4 | Enolase 4 | Lyase activity | 1.9 | down | |
| KCNQ1OT1 | KCNQ1 opposite strand/antisense transcript 1 | non-coding RNA | 1.7 | down | |
| OLR1 | Oxidized low-density lipoprotein receptor 1 | Lipoprotein receptor | 2.1 | down | |
| HOXC11 | Homeobox protein Hox-C11 | Transcription factor | 1.7 | up | |
| PTPRQ | Receptor-type tyrosine-protein phosphatase R | Protein phosphatase | 1.8 | down | |
| PAPLN | Papilin | Peptidase activity | 1.7 | down | |
| TMEM121 | Transmembrane protein 121 | 2 | up | ||
| ATF7IP2 | Activating transcription factor 7-interacting protein 2 | ATPase activity | 2.4 | down | |
| CNBD2 | Cyclic nucleotide-binding domain-containing protein 2 | cAMP binding | 1.7 | up | |
| AHNAK2 | Protein AHNAK2 | RNA binding | 1.8 | down | |
| LIF | Leukemia inhibitory factor | Cytokine activity | 1.8 | down | |
| PDZD4 | PDZ domain-containing protein 4 | Ubiquitin protein ligase activity | 2.1 | up | |
| 24 h after exposure | PPP1R1C | Protein phosphatase 1 regulatory subunit 1C | Signaling molecule, phosphatase inhibitor | 2.6 | up |
| ADAMTS13 | A disintegrin and metalloproteinase with thrombospondin motifs 13 | Metallopeptidase activity | 1.5 | up | |
| PANO | Proapoptotic Nucleolar Protein 1 | Apoptosis-inducing protein | 1.9 | up | |
| NEAT1 | nuclear paraspeckle assembly transcript 1 | Non-coding RNA | 1.5 | up | |
| VAMP1 | Vesicle-associated membrane protein 1 | Transport | 1.5 | up | |
| GOLGA8B | Golgin subfamily A member 8B | Membrane traffic protein | 1.5 | up | |
| GUSBP11 | Putative inactive beta-glucuronidase protein GUSBP11 | Hydrolase activity | 1.5 | up |
| 2.45 GHz | Gene | RNA-seq | RT-PCR | ||||
|---|---|---|---|---|---|---|---|
| FC | Regulation | p-Value | FC | Regulation | p-Value | ||
| CW | RMRP | 8.8 | ↓ | ≤0.05 | 0.73 | ↓ | NS |
| ALMS1 | 2.06 | ↑ | ≤0.05 | 1.14 | ↑ | NS | |
| BSN | 2.37 | ↑ | ≤0.05 | 1.52 | ↑ | ≤0.001 | |
| SYNE2 | 2.06 | ↑ | ≤0.05 | 2.1 | ↑ | NS | |
| AHNAK2 | 2.18 | ↑ | ≤0.05 | 1.0 | - | NS | |
| PW | RMRP | 5.7 | ↓ | ≤0.05 | 0.96 | ↓ | NS |
| AHNAK2 | 1.5 | ↓ | ≤0.05 | 1.99 | ↑ | NS | |
| Gene Name | Description | Assay ID |
|---|---|---|
| RMRP | RNA Component of Mitochondrial RNA Processing Endoribonuclease | Hs03298751_s1 |
| ALMS1 | Cytoskeletal protein binding | Hs00367316_m1 |
| BSN | Scaffolding protein involved in organizing the presynaptic cytoskeleton | Hs01109152_m1 |
| SYNE2 | Actin binding | Hs00794881_m1 |
| AHNAK2 | RNA binding | Hs00292832_m1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Regalbuto, E.; Anselmo, A.; De Sanctis, S.; Franchini, V.; Lista, F.; Benvenuto, M.; Bei, R.; Masuelli, L.; D’Inzeo, G.; Paffi, A.; et al. Human Fibroblasts In Vitro Exposed to 2.45 GHz Continuous and Pulsed Wave Signals: Evaluation of Biological Effects with a Multimethodological Approach. Int. J. Mol. Sci. 2020, 21, 7069. https://doi.org/10.3390/ijms21197069
Regalbuto E, Anselmo A, De Sanctis S, Franchini V, Lista F, Benvenuto M, Bei R, Masuelli L, D’Inzeo G, Paffi A, et al. Human Fibroblasts In Vitro Exposed to 2.45 GHz Continuous and Pulsed Wave Signals: Evaluation of Biological Effects with a Multimethodological Approach. International Journal of Molecular Sciences. 2020; 21(19):7069. https://doi.org/10.3390/ijms21197069
Chicago/Turabian StyleRegalbuto, Elisa, Anna Anselmo, Stefania De Sanctis, Valeria Franchini, Florigio Lista, Monica Benvenuto, Roberto Bei, Laura Masuelli, Guglielmo D’Inzeo, Alessandra Paffi, and et al. 2020. "Human Fibroblasts In Vitro Exposed to 2.45 GHz Continuous and Pulsed Wave Signals: Evaluation of Biological Effects with a Multimethodological Approach" International Journal of Molecular Sciences 21, no. 19: 7069. https://doi.org/10.3390/ijms21197069
APA StyleRegalbuto, E., Anselmo, A., De Sanctis, S., Franchini, V., Lista, F., Benvenuto, M., Bei, R., Masuelli, L., D’Inzeo, G., Paffi, A., Trodella, E., & Sgura, A. (2020). Human Fibroblasts In Vitro Exposed to 2.45 GHz Continuous and Pulsed Wave Signals: Evaluation of Biological Effects with a Multimethodological Approach. International Journal of Molecular Sciences, 21(19), 7069. https://doi.org/10.3390/ijms21197069