A Review of the Action of Magnesium on Several Processes Involved in the Modulation of Hematopoiesis



Abstract
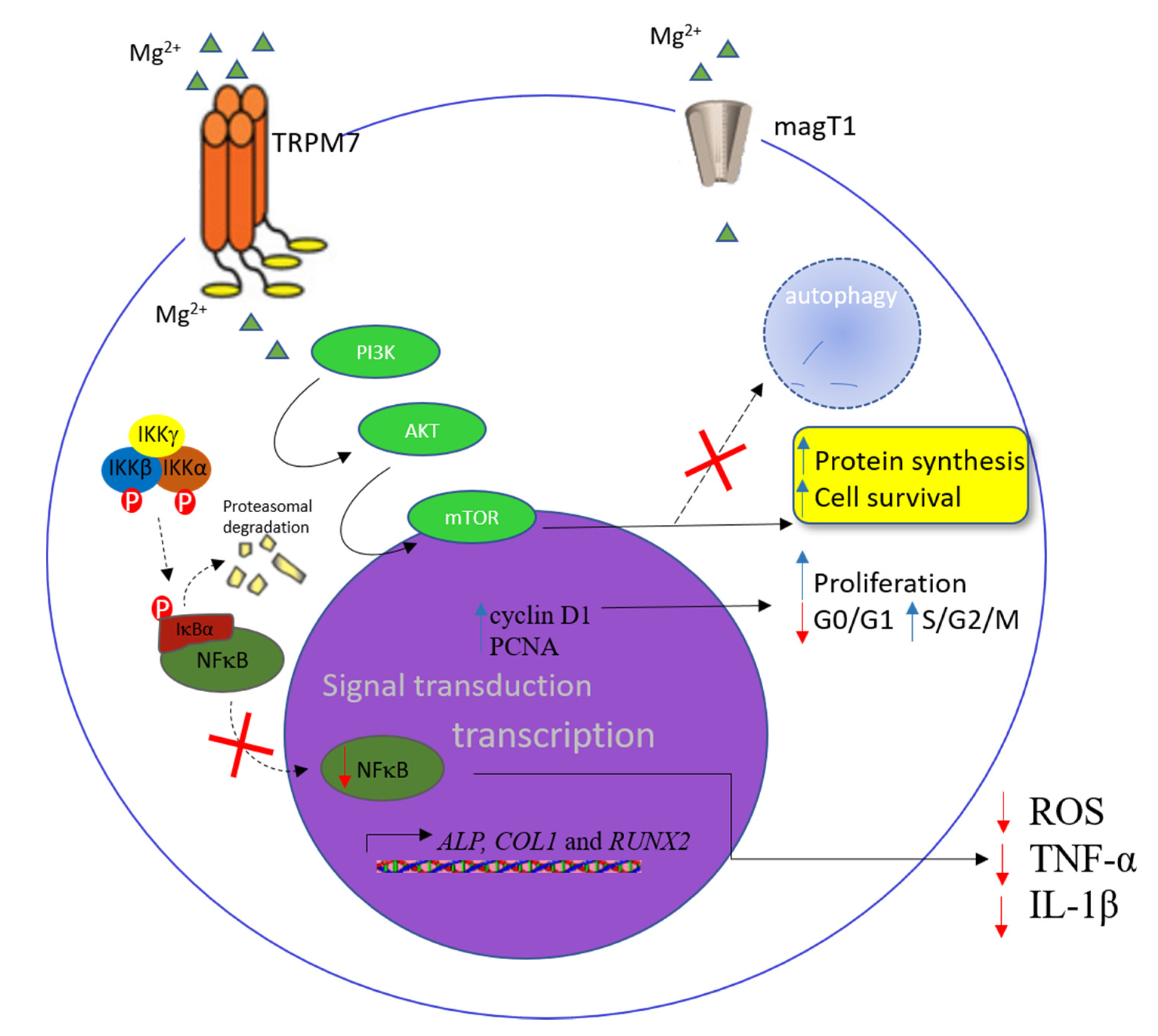
:1. Introduction
2. The Role of Mg2+ in Hematopoiesis
2.1. The Hematopoietic Microenvironment
2.1.1. The Influence of Mg2+ in Mesenchymal Stem Cells, Osteoblasts, and Adipocytes
2.1.2. Mg2+ and Endothelial Cells
2.1.3. Influence of Mg2+ in Macrophages
2.2. Mg2+ and Erythropoiesis
2.3. Mg2+ and Granulopoiesis
2.4. Influence of Mg2+ in Lymphopoiesis
2.5. Influence of Mg2+ in Thrombopoiesis
3. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 1G11 | Murine pulmonary vascular endothelial cell |
| Akt | Protein kinase B |
| ALL | Acute lymphocytic leukemia |
| ALP | Alkaline phosphatase |
| BFU-E | Burst forming unit, erythroid |
| BMD | Bone mineral density |
| BMP2 | Bone morphogenetic protein 2 |
| C/EBPα | CCAAT-enhancer-binding protein α |
| C/EBPβ | CCAAT-enhancer-binding protein β |
| C3H10T1/2 | Murine C3H embryo sarcoma |
| Ca2+ | Calcium |
| c-fos | Member of Fos family of transcription factors |
| CFU-E | Colony-forming unit, erythroid |
| COL10A1 | Collagen type X alpha 1 chain |
| CXCL12 | C–X–C motif chemokine 12 |
| Angpt1 | Angiopoietin 1 |
| CXCL12 | C–X–C motif chemokine 12 |
| CXCR4 | C–X–C chemokine receptor type 4 |
| G-CSF | Granulocyte colony-stimulating factor |
| DC-STAMP | Dendritic cell-specific transmembrane protein |
| DNA | Deoxyribonucleic acid |
| DT40 cell | Chicken B cell line |
| EBV | Epstein–Barr virus |
| ECM | Extracellular matrix |
| EPO | Erythropoietin |
| Fe | Iron |
| FMD | Flow-mediated dilation |
| GATA | GATA-binding protein 1 |
| G-CSF | Granulocyte colony-stimulating factor |
| GM-CSF | Granulocyte macrophage colony-stimulating factor |
| H2O2 | Hydrogen peroxide |
| hFOB1.19 | Human bone osteoblast; SV40 large T antigen transfected |
| HIF1-α | Hypoxia-inducible factor 1α |
| HMGB1 | High-mobility group box 1 |
| hMSCs | Human mesenchymal stem cells |
| HSC | Hematopoietic stem cell |
| HUVEC | Human umbilical vein endothelial cell |
| IgA | Immunoglobulin A |
| IGF2 | Insulin-like growth factor 2 |
| IgG | Immunoglobulin G |
| IgM | Immunoglobulin M |
| IL-1 | Interleukin-1 |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin-8 |
| iNOS | Inducible nitric oxide synthase |
| ITK | Interleukin-2-inducible kinase |
| JAK/STAT | Janus kinase/signal transducer |
| K | Potassium |
| KLF5 | Kruppel-like factor 5 |
| MagT1 | Magnesium transporter 1 |
| Mg2+ | Magnesium |
| mTOR | Mammalian target of rapamycin |
| Na | Sodium |
| NF-ĸB | Nuclear factor kappa B |
| NKG2D | Natural killer group 2 member D |
| NMMIIA | Nonmuscular myosin protein IIA |
| OPG | Osteoprotegerin |
| OST | Oligosaccharyltransferase |
| PCNA | Proliferating cell nuclear antigen |
| CRP | C-reactive protein |
| PGC-1 α | Peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| PGE2 | Prostaglandin E2 |
| PI3-kinase | Phosphoinositide 3-kinase |
| PPARγ2 | Peroxisome proliferator-activated receptor γ2 |
| PTH | Parathyroid hormone |
| PU.1 | Transcription factor PU.1 |
| Rac1 | Ras-related C3 botulinum toxin substrate 1 |
| RANKL | Receptor activator of nuclear factor kappa Β ligand |
| RANTES | Regulated upon activation, normal T cell expressed and secreted |
| RBC | Red blood cells |
| RNA | Ribonucleic acid |
| ROS | Reactive oxygen species |
| RUNX2 | RUNX family transcription factor 2 |
| S1P1 | Sphingosine-1-phosphate receptor 1 |
| S6K | Ribosomal S6 kinase |
| SaOS-2 | Human osteosarcoma cell line |
| SCF | Stem-cell factor |
| sE-selectin | Soluble endothelial selectin |
| sICAM-1 | Soluble intercellular adhesion molecule 1 |
| SIRT1 | Sirtuin-1 |
| STAT | Signal transducer and activator of transcription |
| STT3B | Oligosaccharyltransferase complex catalytic subunit B |
| sVCAM-1 | Soluble vascular cell adhesion molecule 1 |
| TCR | T-cell receptor |
| TNF-α | Tumor necrosis factor α |
| TRPM7 | Transient receptor potential cation channel subfamily M member 7 |
| VCAM1 | Vascular cell adhesion molecule 1 |
| VEGF | Vascular endothelial growth factor |
References
- Nielsen, F.H. Dietary Magnesium and Chronic Disease. Adv. Chronic Kidney Dis. 2018, 25, 230–235. [Google Scholar] [CrossRef]
- Elin, R.J. The Effect of Magnesium Deficiency in Mice on Serum Immunoglobulin Concentrations and Antibody Plaque-Forming Cells. Exp. Boil. Med. 1975, 148, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Wolf, F.I. Magnesium in cell proliferation and differentiation. Front. Biosci. 1999, 4, D607–D617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malpuech-Brugère, C.; Nowacki, W.; Daveau, M.; Gueux, E.; Linard, C.; Rock, E.; Lebreton, J.-P.; Mazur, A.; Rayssiguier, Y. Inflammatory response following acute magnesium deficiency in the rat. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2000, 1501, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Van Orden, R.; Eggett, D.L.; Franz, K.B. Influence of graded magnesium deficiencies on white blood cell counts and lymphocyte subpopulations in rats. Magnes. Res. 2006, 19, 93–101. [Google Scholar] [PubMed]
- Nielsen, F.H. Magnesium deficiency and increased inflammation: Current perspectives. J. Inflamm. Res. 2018, 11, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Desai, B.N.; Navarro, B.; Donovan, A.; Andrews, N.C.; Clapham, D.E. Deletion of Trpm7 Disrupts Embryonic Development and Thymopoiesis Without Altering Mg2+ Homeostasis. Science 2008, 322, 756–760. [Google Scholar] [CrossRef] [Green Version]
- Howe, M.K.; Dowdell, K.; Roy, A.; Niemela, J.E.; Wilson, W.; McElwee, J.J.; Hughes, J.D.; Cohen, J.I. Magnesium Restores Activity to Peripheral Blood Cells in a Patient With Functionally Impaired Interleukin-2-Inducible T Cell Kinase. Front. Immunol. 2019, 10, 2000. [Google Scholar] [CrossRef] [Green Version]
- Yoshizawa, S.; Chaya, A.; Verdelis, K.; Bilodeau, E.A.; Sfeir, C. An in vivo model to assess magnesium alloys and their biological effect on human bone marrow stromal cells. Acta Biomater. 2015, 28, 234–239. [Google Scholar] [CrossRef] [Green Version]
- Luthringer-Feyerabend, B.J.; Willumeit-Römer, R. Effects of magnesium degradation products on mesenchymal stem cell fate and osteoblastogenesis. Gene 2016, 575, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Lima, F.D.S.; Romero, A.B.D.R.; Hastreiter, A.; Nogueira-Pedro, A.; Makiyama, E.; Colli, C.; Fock, R.A. An insight into the role of magnesium in the immunomodulatory properties of mesenchymal stem cells. J. Nutr. Biochem. 2018, 55, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Mao, X.; Ling, J.; Chen, C.; Zhang, W. Role of Magnesium Transporter Subtype 1 (MagT1) in the Osteogenic Differentiation of Rat Bone Marrow Stem Cells. Boil. Trace Element Res. 2015, 171, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Taichman, R.S.; Emerson, S.G. Human osteoblasts support hematopoiesis through the production of granulocyte colony-stimulating factor. J. Exp. Med. 1994, 179, 1677–1682. [Google Scholar] [CrossRef] [PubMed]
- Taichman, R.S.; Reilly, M.; Emerson, S. Human osteoblasts support human hematopoietic progenitor cells in vitro bone marrow cultures. Blood 1996, 87, 518–524. [Google Scholar] [CrossRef] [PubMed]
- McCreary, P.A.; Battifora, H.A.; Hahneman, B.M.; Laing, G.H.; Hass, G.M.; Singh, K. Leukocytosis, Bone Marrow Hyperplasia and Leukemia in Chronic Magnesium Deficiency in the Rat. Blood 1967, 29, 683–690. [Google Scholar] [CrossRef]
- Stritt, S.; Nurden, P.; Favier, R.; Favier, M.; Ferioli, S.; Gotru, S.K.; Van Eeuwijk, J.M.M.; Schulze, H.; Nurden, A.T.; Lambert, M.P.; et al. Defects in TRPM7 channel function deregulate thrombopoiesis through altered cellular Mg2+ homeostasis and cytoskeletal architecture. Nat. Commun. 2016, 7, 11097. [Google Scholar] [CrossRef]
- Krishnamoorthy, M.; Wasim, L.; Buhari, F.H.M.; Zhao, T.; Mahtani, T.; Ho, J.; Kang, S.; Deason-Towne, F.; Perraud, A.-L.; Schmitz, C.; et al. The channel-kinase TRPM7 regulates antigen gathering and internalization in B cells. Sci. Signal. 2018, 11, eaah6692. [Google Scholar] [CrossRef] [Green Version]
- Yee, N.S.; Kazi, A.A.; Yee, R.K. Cellular and Developmental Biology of TRPM7 Channel-Kinase: Implicated Roles in Cancer. Cells 2014, 3, 751–777. [Google Scholar] [CrossRef]
- Zierler, S.; Sumoza-Toledo, A.; Suzuki, S.; Dúill, F. Ó; Ryazanova, L.V.; Penner, R.; Ryazanov, A.G.; Fleig, A. TRPM7 kinase activity regulates murine mast cell degranulation. J. Physiol. 2016, 594, 2957–2970. [Google Scholar] [CrossRef] [Green Version]
- Nadolni, W.; Zierler, S. The Channel-Kinase TRPM7 as Novel Regulator of Immune System Homeostasis. Cells 2018, 7, 109. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Sukumaran, P.; Singh, B.B. Magnesium-Induced Cell Survival Is Dependent on TRPM7 Expression and Function. Mol. Neurobiol. 2019, 57, 528–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, R.; Meng, Z.; Liu, T.; Wang, G.; Qian, G.; Cao, T.; Guan, X.; Dan, H.; Xiao, Y.; Wang, X. Decreased TRPM7 inhibits activities and induces apoptosis of bladder cancer cells via ERK1/2 pathway. Oncotarget 2016, 7, 72941–72960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Z.G.; Rios, F.J.; Montezano, A.C.; Touyz, R.M. TRPM7, Magnesium, and Signaling. Int. J. Mol. Sci. 2019, 20, 1877. [Google Scholar] [CrossRef] [Green Version]
- Ikari, A.; Sawada, H.; Sanada, A.; Tonegawa, C.; Yamazaki, Y.; Sugatani, J. Magnesium deficiency suppresses cell cycle progression mediated by increase in transcriptional activity of p21Cip1 and p27Kip1 in renal epithelial NRK-52E cells. J. Cell. Biochem. 2011, 112, 3563–3572. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, A. Role of magnesium in genomic stability. Mutat. Res. Mol. Mech. Mutagen. 2001, 475, 113–121. [Google Scholar] [CrossRef]
- Covacci, V.; Bruzzese, N.; Sgambato, A.; Di Francesco, A.; Russo, M.A.; Wolf, F.I.; Cittadini, A. Magnesium restriction induces granulocytic differentiation and expression of p27Kip1 in human leukemic HL-60 cells. J. Cell. Biochem. 1998, 70, 313–322. [Google Scholar] [CrossRef]
- Wu, L.; Witte, F.M.; Schilling, A.F.; Willumeit-Römer, R.; Luthringer-Feyerabend, B.J. Effects of extracellular magnesium extract on the proliferation and differentiation of human osteoblasts and osteoclasts in coculture. Acta Biomater. 2015, 27, 294–304. [Google Scholar] [CrossRef]
- Mammoli, F.; Castiglioni, S.; Parenti, S.; Cappadone, C.; Farruggia, G.; Iotti, S.; Davalli, P.; Maier, J.A.; Grande, A.; Frassineti, C. Magnesium Is a Key Regulator of the Balance between Osteoclast and Osteoblast Differentiation in the Presence of Vitamin D3. Int. J. Mol. Sci. 2019, 20, 385. [Google Scholar] [CrossRef] [Green Version]
- Martin, H.; Richert, L.; Berthelot, A. Magnesium deficiency induces apoptosis in primary cultures of rat hepatocytes. J. Nutr. 2003, 133, 2505–2511. [Google Scholar] [CrossRef] [Green Version]
- Ohyama, T. New Aspects of Magnesium Function: A Key Regulator in Nucleosome Self-Assembly, Chromatin Folding and Phase Separation. Int. J. Mol. Sci. 2019, 20, 4232. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Tocados, J.M.; Herencia, C.; Martínez-Moreno, J.M.; De Oca, A.M.; Rodríguez-Ortiz, M.E.; Vergara, N.; Blanco, A.; Steppan, S.; Almadén, Y.; Rodríguez, M.; et al. Magnesium Chloride promotes Osteogenesis through Notch signaling activation and expansion of Mesenchymal Stem Cells. Sci. Rep. 2017, 7, 7839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ma, X.-Y.; Feng, Y.-F.; Ma, Z.-S.; Ma, T.-C.; Zhang, Y.; Li, X.; Wang, L.; Lei, W. Magnesium Ions Promote the Biological Behaviour of Rat Calvarial Osteoblasts by Activating the PI3K/Akt Signalling Pathway. Boil. Trace Element Res. 2017, 179, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zu, H.; Zhao, D.W.; Yang, K.; Tian, S.; Yu, X.; Lu, F.; Liu, B.; Yu, X.; Wang, B.; et al. Ion channel functional protein kinase TRPM7 regulates Mg ions to promote the osteoinduction of human osteoblast via PI3K pathway: In vitro simulation of the bone-repairing effect of Mg-based alloy implant. Acta Biomater. 2017, 63, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Leidi, M.; Dellera, F.; Mariotti, M.; Maier, J.A. High magnesium inhibits human osteoblast differentiation in vitro. Magnes Res 2011, 24, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rude, R.K.; Gruber, H.; Wei, L.; Frausto, A.; Mills, B. Magnesium Deficiency: Effect on Bone and Mineral Metabolism in the Mouse. Calcif. Tissue Int. 2003, 72, 32–41. [Google Scholar] [CrossRef]
- Bernardini, D.; Nasulewic, A.; Mazur, A.; Maier, J.A.M. Magnesium and microvascular endothelial cells: A role in inflammation and angiogenesis. Front. Biosci. 2005, 10, 1177–1182. [Google Scholar] [CrossRef] [Green Version]
- Ferrè, S.; Baldoli, E.; Leidi, M.; Maier, J.A. Magnesium deficiency promotes a pro-atherogenic phenotype in cultured human endothelial cells via activation of NFkB. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2010, 1802, 952–958. [Google Scholar] [CrossRef]
- Baldoli, E.; Maier, J.A. Silencing TRPM7 mimics the effects of magnesium deficiency in human microvascular endothelial cells. Angiogenesis 2011, 15, 47–57. [Google Scholar] [CrossRef]
- Zhu, D.; You, J.; Zhao, N.; Xu, H. Magnesium Regulates Endothelial Barrier Functions through TRPM7, MagT1, and S1P1. Adv. Sci. 2019, 6, 1901166. [Google Scholar] [CrossRef] [Green Version]
- Cunha, A.R.; D’El-Rei, J.; Medeiros, F.; Umbelino, B.; Oigman, W.; Touyz, R.M.; Neves, M.F. Oral magnesium supplementation improves endothelial function and attenuates subclinical atherosclerosis in thiazide-treated hypertensive women. J. Hypertens. 2017, 35, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Joris, P.; Plat, J.; Bakker, S.J.L.; Mensink, R.P. Effects of long-term magnesium supplementation on endothelial function and cardiometabolic risk markers: A randomized controlled trial in overweight/obese adults. Sci. Rep. 2017, 7, 106. [Google Scholar] [CrossRef] [PubMed]
- Shogi, T.; Oono, H.; Nakagawa, M.; Miyamoto, A.; Ishiguro, S.; Nishio, A. Effects of a low extracellular magnesium concentration and endotoxin on IL-1beta and TNF-alpha release from, and mRNA levels in, isolated rat alveolar macrophages. Magnes. Res. 2002, 15, 147–152. [Google Scholar] [PubMed]
- Liu, Z.; Chang, Y.; Zhang, J.; Huang, X.; Jiang, J.; Li, S.; Wang, Z. Magnesium deficiency promotes secretion of high-mobility group box 1 protein from lipopolysaccharide-activated macrophages in vitro. J. Surg. Res. 2013, 180, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, J.; Romani, A.M.; Valentin-Torres, A.M.; Luciano, A.A.; Ramirez Kitchen, C.M.; Funderburg, N.; Mesiano, S.; Bernstein, H.B. Magnesium decreases inflammatory cytokine production: A novel innate immunomodulatory mechanism. J. Immunol. 2012, 188, 6338–6346. [Google Scholar] [CrossRef] [Green Version]
- Elin, R.J.; Utter, A.; Tan, H.K.; Corash, L. Effect of magnesium deficiency on erythrocyte aging in rats. Am. J. Pathol. 1980, 100, 765–778. [Google Scholar]
- Sanchez-Morito, N.; Planells, E.; Aranda, P.; Llopis, J. Influence of magnesium deficiency on the bioavailability and tissue distribution of iron in the rat. J. Nutr. Biochem. 2000, 11, 103–108. [Google Scholar] [CrossRef]
- Shi, Z.; Hu, X.; He, K.; Yuan, B.; Garg, M. Joint association of magnesium and iron intake with anemia among Chinese adults. Nutrition 2008, 24, 977–984. [Google Scholar] [CrossRef]
- Zhan, Y.; Chen, R.; Zheng, W.; Guo, C.; Lu, L.; Ji, X.; Chi, Z.; Yu, J. Association between serum magnesium and anemia: China health and nutrition survey. Biol. Trace Element Res. 2014, 159, 39–45. [Google Scholar] [CrossRef]
- Eltayeb, R.; Rayis, D.A.; Sharif, M.E.; Ahmed, A.B.A.; Elhardello, O.; Adam, I. The prevalence of serum magnesium and iron deficiency anaemia among Sudanese women in early pregnancy: A cross-sectional study. Trans. R. Soc. Trop. Med. Hyg. 2019, 113, 31–35. [Google Scholar] [CrossRef]
- Cinar, V.; Nizamlioglu, M.; Mogulkoc, R.; Baltaci, A.K. Effects of magnesium supplementation on blood parameters of athletes at rest and after exercise. Biol. Trace Elem. Res. 2007, 115, 205–212. [Google Scholar] [CrossRef]
- Hungerford, G.F. Role of histamine in producing the eosinophilia of magnesium deficiency. Proc. Soc. Exp. Biol. Med. 1964, 115, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Kraeuter, S.L.; Schwartz, R. Blood and mast cell histamine levels in magnesium-deficient rats. J. Nutr. 1980, 110, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, L.F. Variations in the mast cell population of skin and bone marrow in magnesium-deprived rats. The influence of sex hormones. J. Nutr. 1977, 107, 2164–2170. [Google Scholar] [CrossRef] [PubMed]
- Bussière, F.I.; Gueux, E.; Rock, E.; Girardeau, J.-P.; Tridon, A.; Mazur, A.; Rayssiguier, Y. Increased phagocytosis and production of reactive oxygen species by neutrophils during magnesium deficiency in rats and inhibition by high magnesium concentration. Br. J. Nutr. 2002, 87, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Zhan, Y.; Yu, J.; Sui, L. Prevalence of Thrombocytopenia and Its Association with Serum Magnesium. Biol. Trace Elem. Res. 2016, 169, 46–51. [Google Scholar] [CrossRef]
- Rishi, M.; Ahmad, A.; Makheja, A.; Karcher, D.; Bloom, S. Effects of reduced dietary magnesium on platelet production and function in hamsters. Lab. Investig. 1990, 63, 717–721. [Google Scholar]
- Maximow, A.A. Relation of Blood cells to connective tissues and endothelium. Physiol. Rev. 1924, 4, 533–563. [Google Scholar] [CrossRef]
- Owen, R.D.; Miller, N.E.; Bailey, C.J.; Stevenson, J.A.F. Immunogenetic consequences of vascular anastomoses between bovine twins. Science 1945, 102, 400–401. [Google Scholar] [CrossRef]
- McCulloch, J.E.T.A. A Direct Measurement of the Radiation Sensitivity of Normal Mouse Bone Marrow Cells. Radiat. Res. 1961, 14, 213. [Google Scholar]
- Papayannopoulou, T.; Scadden, D.T. Stem-cell ecology and stem cells in motion. Blood 2008, 111, 3923–3930. [Google Scholar] [CrossRef]
- Weissman, I.L.; Shizuru, J.A. The origins of the identification and isolation of hematopoietic stem cells, and their capability to induce donor-specific transplantation tolerance and treat autoimmune diseases. Blood 2008, 112, 3543–3553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.C.; Lodish, H.F. Cytokines regulating hematopoietic stem cell function. Curr. Opin. Hematol. 2008, 15, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gao, S.; Xia, J.; Liu, F. Hematopoietic Hierarchy – An Updated Roadmap. Trends Cell Boil. 2018, 28, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Orkin, S.H.; Zon, L.I. Hematopoiesis: An Evolving Paradigm for Stem Cell Biology. Cell 2008, 132, 631–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieger, M.A.; Schroeder, T. Hematopoiesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008250. [Google Scholar] [CrossRef]
- Schofield, R. The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood Cells 1978, 4, 7–25. [Google Scholar]
- Agas, D.; Marchetti, L.; Douni, E.; Sabbieti, M.G. The unbearable lightness of bone marrow homeostasis. Cytokine Growth Factor Rev. 2015, 26, 347–359. [Google Scholar] [CrossRef]
- Scadden, D.T. Nice neighborhood: Emerging concepts of the stem cell niche. Cell 2014, 157, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Kopp, H.-G.; Avecilla, S.T.; Hooper, A.T.; Rafii, S. The Bone Marrow Vascular Niche: Home of HSC Differentiation and Mobilization. Physiology 2005, 20, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Godavarthy, P.S.; Krause, D.S. The bone marrow microenvironment in health and disease at a glance. J. Cell Sci. 2018, 131, jcs201707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naveiras, O.; Nardi, V.; Wenzel, P.L.; Hauschka, P.V.; Fahey, F.; Daley, G.Q. Bone-marrow adipocytes as negative regulators of the haematopoietic microenvironment. Nature 2009, 460, 259–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimura, R.; Li, L. Shifting in Balance between Osteogenesis and Adipogenesis Substantially Influences Hematopoiesis. J. Mol. Cell Boil. 2009, 2, 61–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rude, R.K.; Gruber, H.E. Magnesium deficiency and osteoporosis: Animal and human observations. J. Nutr. Biochem. 2004, 15, 710–716. [Google Scholar] [CrossRef]
- Castiglioni, S.; Cazzaniga, A.; Albisetti, W.; Maier, J.A.M. Magnesium and Osteoporosis: Current State of Knowledge and Future Research Directions. Nutrients 2013, 5, 3022–3033. [Google Scholar] [CrossRef] [Green Version]
- Castiglioni, S.; Romeo, V.; Locatelli, L.; Cazzaniga, A.; Maier, J.A. TRPM7 and MagT1 in the osteogenic differentiation of human mesenchymal stem cells in vitro. Sci. Rep. 2018, 8, 16195. [Google Scholar] [CrossRef]
- Castiglioni, S.; Romeo, V.; Locatelli, L.; Zocchi, M.; Zecchini, S.; Maier, J.A. The simultaneous downregulation of TRPM7 and MagT1 in human mesenchymal stem cells in vitro: Effects on growth and osteogenic differentiation. Biochem. Biophys. Res. Commun. 2019, 513, 159–165. [Google Scholar] [CrossRef]
- Gruber, H.; Rude, R.K.; Wei, L.; Frausto, A.; Mills, B.G.; Norton, H.J. Magnesium deficiency: Effect on bone mineral density in the mouse appendicular skeleton. BMC Musculoskelet. Disord. 2003, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Rude, R.K.; Gruber, H.E.; Wei, L.; Frausto, A. Immunolocalization of RANKL is Increased and OPG Decreased during Dietary Magnesium Deficiency in the Rat. Nutr. Metab. 2005, 2, 24. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Rajaratnam, J.H.; Denton, J.; A Hoyland, J.; Byers, R. Adipocytic proportion of bone marrow is inversely related to bone formation in osteoporosis. J. Clin. Pathol. 2002, 55, 693–698. [Google Scholar] [CrossRef]
- Justesen, J.; Stenderup, K.; Ebbesen, E.; Mosekilde, L.; Steiniche, T.; Kassem, M. Adipocyte tissue volume in bone marrow is increased with aging and in patients with osteoporosis. Biogerontology 2001, 2, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Beekman, K.M.; Veldhuis-Vlug, A.G.; Heijer, M.D.; Maas, M.; Oleksik, A.; Tanck, M.W.; Ott, S.M.; Hof, R.J.V. ‘T; Lips, P.; Bisschop, P.H.; et al. The effect of raloxifene on bone marrow adipose tissue and bone turnover in postmenopausal women with osteoporosis. Bone 2019, 118, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.V.; Sigurdsson, S.; Hue, T.F.; Lang, T.F.; Harris, T.B.; Rosen, C.J.; Vittinghoff, E.; Siggeirsdottir, K.; Sigurdsson, G.; Oskarsdottir, D.; et al. Vertebral Bone Marrow Fat Associated With Lower Trabecular BMD and Prevalent Vertebral Fracture in Older Adults. J. Clin. Endocrinol. Metab. 2013, 98, 2294–2300. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Li, N.; Cheng, H.; Yen, M.-L. Oxidative stress induces imbalance of adipogenic/osteoblastic lineage commitment in mesenchymal stem cells through decreasing SIRT1 functions. J. Cell. Mol. Med. 2017, 22, 786–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrault, I.; Zimowska, W.; Mathieu, J.; Bayle, D.; Rock, E.; Favier, A.; Rayssiguier, Y.; Mazur, A. Changes in gene expression in rat thymocytes identified by cDNA array support the occurrence of oxidative stress in early magnesium deficiency. Biochim. Biophys. Acta 2002, 1586, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Malpuech-Brugère, C.; Nowacki, W.; Gueux, E.; Kuryszko, J.; Rock, E.; Rayssiguier, Y.; Mazur, A. Accelerated thymus involution in magnesium-deficient rats is related to enhanced apoptosis and sensitivity to oxidative stress. Br. J. Nutr. 1999, 81, 405–411. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wu, Z.; Chen, Y.; Qiao, J.; Gao, M.; Yuan, J.; Nie, W.; Guo, Y. Magnesium Deficiency Enhances Hydrogen Peroxide Production and Oxidative Damage in Chick Embryo Hepatocyte In Vitro. BioMetals 2006, 19, 71–81. [Google Scholar] [CrossRef]
- Leidi, M.; Dellera, F.; Mariotti, M.; Banfi, G.; Crapanzano, C.; Albisetti, W.; Maier, J.A. Nitric oxide mediates low magnesium inhibition of osteoblast-like cell proliferation. J. Nutr. Biochem. 2012, 23, 1224–1229. [Google Scholar] [CrossRef]
- Rock, E.; Astier, C.; Lab, C.; Malpuech, C.; Nowacki, W.; Gueux, E.; Mazur, A.; Rayssiguier, Y. Magnesium deficiency in rats induces a rise in plasma nitric oxide. Magnes. Res. 1995, 8, 237–242. [Google Scholar]
- Cunha, M.C.R.; Lima, F.D.S.; Vinolo, M.A.R.; Hastreiter, A.; Curi, R.; Borelli, P.; Fock, R.A. Protein Malnutrition Induces Bone Marrow Mesenchymal Stem Cells Commitment to Adipogenic Differentiation Leading to Hematopoietic Failure. PLoS ONE 2013, 8, e58872. [Google Scholar] [CrossRef] [Green Version]
- Wilson, A.; Fu, H.; Schiffrin, M.; Winkler, C.; Koufany, M.; Jouzeau, J.-Y.; Bonnet, N.; Gilardi, F.; Renevey, F.; Luther, S.A.; et al. Lack of Adipocytes Alters Hematopoiesis in Lipodystrophic Mice. Front. Immunol. 2018, 9, 2573. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.O.; Yu, H.; Yue, R.; Zhao, Z.; Rios, J.J.; Naveiras, O.; Morrison, S.J. Bone marrow adipocytes promote the regeneration of stem cells and haematopoiesis by secreting SCF. Nature 2017, 19, 891–903. [Google Scholar] [CrossRef]
- Xu, Y.; Takahashi, Y.; Wang, Y.; Hama, A.; Nishio, N.; Muramatsu, H.; Tanaka, M.; Yoshida, N.; Villalobos, I.B.; Yagasaki, H.; et al. Downregulation of GATA-2 and overexpression of adipogenic gene-PPARγ in mesenchymal stem cells from patients with aplastic anemia. Exp. Hematol. 2009, 37, 1393–1399. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; He, J.; Koh, S.P.; Zhong, Y.; Liu, Z.; Wang, Z.; Zhang, Y.; Li, Z.; Tam, B.T.; Lin, P.; et al. Reprogrammed marrow adipocytes contribute to myeloma-induced bone disease. Sci. Transl. Med. 2019, 11, eaau9087. [Google Scholar] [CrossRef] [PubMed]
- Nombela-Arrieta, C.; Pivarnik, G.; Winkel, B.; Canty, K.J.; Harley, B.; Mahoney, J.E.; Park, S.-Y.; Lu, J.; Protopopov, A.; Silberstein, L.E. Quantitative imaging of haematopoietic stem and progenitor cell localization and hypoxic status in the bone marrow microenvironment. Nat. Cell Biol. 2013, 15, 533–543. [Google Scholar] [CrossRef]
- Filipowska, J.; Tomaszewski, K.A.; Niedźwiedzki, Ł.; Walocha, J.A.; Niedźwiedzki, T. The role of vasculature in bone development, regeneration and proper systemic functioning. Angiogenesis 2017, 20, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Kiel, M.J.; Yilmaz, Ö.H.; Iwashita, T.; Yilmaz, O.H.; Terhorst, C.; Morrison, S.J. SLAM Family Receptors Distinguish Hematopoietic Stem and Progenitor Cells and Reveal Endothelial Niches for Stem Cells. Cell 2005, 121, 1109–1121. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Saunders, T.L.; Enikolopov, G.; Morrison, S.J. Endothelial and perivascular cells maintain haematopoietic stem cells. Nature 2012, 481, 457–462. [Google Scholar] [CrossRef] [Green Version]
- Butler, J.M.; Nolan, D.J.; Vertes, E.L.; Varnum-Finney, B.; Kobayashi, H.; Hooper, A.T.; Seandel, M.; Shido, K.; White, I.A.; Kobayashi, M.; et al. Endothelial Cells Are Essential for the Self-Renewal and Repopulation of Notch-Dependent Hematopoietic Stem Cells. Cell Stem Cell 2010, 6, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Poulos, M.G.; Guo, P.; Kofler, N.M.; Pinho, S.; Gutkin, M.C.; Tikhonova, A.; Aifantis, I.; Frenette, P.S.; Kitajewski, J.; Rafii, S.; et al. Endothelial Jagged-1 Is Necessary for Homeostatic and Regenerative Hematopoiesis. Cell Rep. 2013, 4, 1022–1034. [Google Scholar] [CrossRef] [Green Version]
- Shahrabi, S.; Rezaeeyan, H.; Ahmadzadeh, A.; Shahjahani, M.; Saki, N. Bone marrow blood vessels: Normal and neoplastic niche. Oncol. Rev. 2016, 10, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itkin, T.; Gur-Cohen, S.; Spencer, J.A.; Schajnovitz, A.; Ramasamy, S.; Kusumbe, A.P.; Ledergor, G.; Jung, Y.; Milo, I.; Poulos, M.G.; et al. Distinct bone marrow blood vessels differentially regulate haematopoiesis. Nature 2016, 532, 323–328. [Google Scholar] [CrossRef]
- Maier, J.A. Endothelial cells and magnesium: Implications in atherosclerosis. Clin. Sci. 2011, 122, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, J.A.; Malpuech-Brugère, C.; Zimowska, W.; Rayssiguier, Y.; Mazur, A. Low magnesium promotes endothelial cell dysfunction: Implications for atherosclerosis, inflammation and thrombosis. Biochim. Biophys. Acta 2004, 1689, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Baldoli, E.; Castiglioni, S.; Maier, J.A. Regulation and Function of TRPM7 in Human Endothelial Cells: TRPM7 as a Potential Novel Regulator of Endothelial Function. PLoS ONE 2013, 8, e59891. [Google Scholar] [CrossRef] [Green Version]
- Mofrad, M.D.; Djafarian, K.; Mozaffari, H.; Shab-Bidar, S. Effect of magnesium supplementation on endothelial function: A systematic review and meta-analysis of randomized controlled trials. Atherosclerosis 2018, 273, 98–105. [Google Scholar] [CrossRef]
- Winkler, I.G.; Sims, N.A.; Pettit, A.R.; Barbier, V.; Nowlan, B.; Helwani, F.; Poulton, I.J.; van Rooijen, N.; Alexander, K.A.; Raggatt, L.J.; et al. Bone marrow macrophages maintain hematopoietic stem cell (HSC) niches and their depletion mobilizes HSCs. Blood 2010, 116, 4815–4828. [Google Scholar] [CrossRef] [Green Version]
- McCabe, A.; MacNamara, K.C. Macrophages: Key regulators of steady-state and demand-adapted hematopoiesis. Exp. Hematol. 2016, 44, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Chow, A.; Lucas, D.; Hidalgo, A.; Méndez-Ferrer, S.; Hashimoto, D.; Scheiermann, C.; Battista, M.; Leboeuf, M.; Prophete, C.; van Rooijen, N.; et al. Bone marrow CD169+ macrophages promote the retention of hematopoietic stem and progenitor cells in the mesenchymal stem cell niche. J. Exp. Med. 2011, 208, 261–271. [Google Scholar] [CrossRef]
- Chasis, J.A.; Mohandas, N. Erythroblastic islands: Niches for erythropoiesis. Blood 2008, 112, 470–478. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Xue, W.; Li, M.; Dong, M.; Wang, J.; Wang, X.; Li, X.; Chen, K.; Zhang, W.; Wu, S.; et al. VCAM-1+ macrophages guide the homing of HSPCs to a vascular niche. Nature 2018, 564, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Cavaillon, J.M. Cytokines and macrophages. Biomed. Pharmacother. 1994, 48, 445–453. [Google Scholar] [CrossRef]
- Metcalf, D. Hematopoietic cytokines. Blood 2008, 111, 485–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libako, P.; Nowacki, W.; Castiglioni, S.; Mazur, A.; Maier, J.A. Extracellular magnesium and calcium blockers modulate macrophage activity. Magnes. Res. 2016, 29, 11–21. [Google Scholar] [CrossRef]
- Tam, M.; Gomez, S.; González-Gross, M.; Marcos, A. Possible roles of magnesium on the immune system. Eur. J. Clin. Nutr. 2003, 57, 1193–1197. [Google Scholar] [CrossRef] [Green Version]
- Weglicki, W.B.; Phillips, T.M. Pathobiology of magnesium deficiency: A cytokine/neurogenic inflammation hypothesis. Am. J. Physiol. Integr. Comp. Physiol. 1992, 263, R734–R737. [Google Scholar] [CrossRef] [Green Version]
- Mazur, A.; Maier, J.A.; Rock, E.; Gueux, E.; Nowacki, W.; Rayssiguier, Y. Magnesium and the inflammatory response: Potential physiopathological implications. Arch. Biochem. Biophys. 2007, 458, 48–56. [Google Scholar] [CrossRef]
- Yokoyama, T.; Oono, H.; Miyamoto, A.; Ishiguro, S.; Nishio, A. Magnesium-deficient medium enhances NO production in alveolar macrophages isolated from rats. Life Sci. 2003, 72, 1247–1257. [Google Scholar] [CrossRef]
- Shigematsu, M.; Tomonaga, S.; Shimokawa, F.; Murakami, M.; Imamura, T.; Matsui, T.; Funaba, M. Regulatory responses of hepatocytes, macrophages and vascular endothelial cells to magnesium deficiency. J. Nutr. Biochem. 2018, 56, 35–47. [Google Scholar] [CrossRef]
- Hu, T.; Xu, H.; Wang, C.; Qin, H.; An, Z. Magnesium enhances the chondrogenic differentiation of mesenchymal stem cells by inhibiting activated macrophage-induced inflammation. Sci. Rep. 2018, 8, 3406. [Google Scholar] [CrossRef]
- Su, N.-Y.; Peng, T.-C.; Tsai, P.-S.; Huang, C.-J. Phosphoinositide 3-kinase/Akt pathway is involved in mediating the anti-inflammation effects of magnesium sulfate. J. Surg. Res. 2013, 185, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, X.; Xu, M.; Yang, F.; Wang, W.; Niu, X. In vitro immunomodulation of magnesium on monocytic cell toward anti-inflammatory macrophages. Regen. Biomater. 2020, 7, 391–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Cao, H.; Zhao, Y.; Cheng, M.; Qin, H.; Cheng, T.; Hu, Y.; Zhang, X.; Liu, X. In vitro and in vivo responses of macrophages to magnesium-doped titanium. Sci. Rep. 2017, 7, 42707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, M.; Passegué, E. TNF-α Coordinates Hematopoietic Stem Cell Survival and Myeloid Regeneration. Cell Stem Cell 2019, 25, 357–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietras, E.M.; Mirantes-Barbeito, C.; Fong, S.L.; Loeffler, D.; Kovtonyuk, L.V.; Zhang, S.; Lakshminarasimhan, R.; Chin, C.P.; Techner, J.-M.; Will, B.; et al. Chronic interleukin-1 exposure drives haematopoietic stem cells towards precocious myeloid differentiation at the expense of self-renewal. Nature 2016, 18, 607–618. [Google Scholar] [CrossRef]
- Barminko, J.; Reinholt, B.; Baron, M.H. Development and differentiation of the erythroid lineage in mammals. Dev. Comp. Immunol. 2016, 58, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Comazzetto, S.; Murphy, M.M.; Berto, S.; Jeffery, E.; Zhao, Z.; Morrison, S.J. Restricted Hematopoietic Progenitors and Erythropoiesis Require SCF from Leptin Receptor+ Niche Cells in the Bone Marrow. Cell Stem Cell 2019, 24, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Mello, F.V.; Land, M.G.P.; Costa, E.S.; Teodósio, C.; Sanchez, M.-L.; Bárcena, P.; Peres, R.T.; Pedreira, C.E.; Alves, L.R.; Orfao, A. Maturation-associated gene expression profiles during normal human bone marrow erythropoiesis. Cell Death Discov. 2019, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Eggold, J.T.; Rankin, E.B. Erythropoiesis, EPO, macrophages, and bone. Bone 2019, 119, 36–41. [Google Scholar] [CrossRef]
- Giger, K.M.; Kalfa, T.A. Phylogenetic and Ontogenetic View of Erythroblastic Islands. Biomed. Res. Int. 2015, 2015, 873628. [Google Scholar] [CrossRef] [Green Version]
- Koury, M.J.; Ponka, P. New insights into erythropoiesis: The roles of folate, vitamin B12, and iron. Annu Rev. Nutr. 2004, 24, 105–131. [Google Scholar] [CrossRef] [PubMed]
- Bang, O.; Orskov, S.L. The magnesium content of the erythrocytes in pernicious and some other anemias. J. Clin. Investig. 1939, 18, 497–500. [Google Scholar] [CrossRef] [Green Version]
- Cohlan, S.Q.; Jansen, V.; Dancis, J.; Piomelli, S. Microcytic anemia with erythroblastosis in offspring of magnesium-deprived rats. Blood 1970, 36, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Piomelli, S.; Jansen, V.; Dancis, J. The hemolytic anemia of magnesium deficiency in adult rats. Blood. 1973, 41, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Cosens, G.; Diamond, I.; Theriault, L.L.; Hurley, L.S. Magnesium deficiency anemia in the rat fetus. Pediatr. Res. 1977, 11, 758–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keen, C.L.; Lowney, P.; Gershwin, M.E.; Hurley, L.S.; Stern, J.S. Dietary magnesium intake influences exercise capacity and hematologic parameters in rats. Metabolism 1987, 36, 788–793. [Google Scholar] [CrossRef]
- De Franceschi, L.; Beuzard, Y.; Jouault, H.; Brugnara, C. Modulation of erythrocyte potassium chloride cotransport, potassium content, and density by dietary magnesium intake in transgenic SAD mouse. Blood 1996, 88, 2738–2744. [Google Scholar] [CrossRef]
- Mak, I.T.; Stafford, R.; Weglicki, W.B. Loss of red blood cell glutathione during Mg deficiency: Prevention by vitamin, E.; D-propranolol, and chloroquine. Am. J. Physiol. 1994, 267 Pt 1, C1366–C1370. [Google Scholar] [CrossRef]
- Mak, I.T.; Komarov, A.M.; Wagner, T.L.; Stafford, R.E.; Dickens, B.F.; Weglicki, W.B. Enhanced NO production during Mg deficiency and its role in mediating red blood cell glutathione loss. Am. J. Physiol. 1996, 271 Pt 1, C385–C390. [Google Scholar] [CrossRef]
- Schauf, B.; Mannschreck, B.; Becker, S.; Dietz, K.; Wallwiener, D.; Aydeniz, B. Evaluation of red blood cell deformability and uterine blood flow in pregnant women with preeclampsia or iugr and reduced uterine blood flow following the intravenous application of magnesium. Hypertens. Pregnancy 2004, 23, 331–343. [Google Scholar] [CrossRef]
- Schauf, B.; Becker, S.; Abele, H.; Klever, T.; Wallwiener, D.; Aydeniz, B. Effect of magnesium on red blood cell deformability in pregnancy. Hypertens. Pregnancy 2005, 24, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Hall, J.; Byles, J.; Shi, Z. Dietary pattern, serum magnesium, ferritin, C-reactive protein and anaemia among older people. Clin Nutr. 2017, 36, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Jelkmann, W. Regulation of erythropoietin production. J. Physiol. 2011, 589 Pt 6, 1251–1258. [Google Scholar] [CrossRef]
- Yu, L.; Song, J.; Lu, X.; Zu, Y.; Li, H.; Wang, S. Association between Serum Magnesium and Erythropoietin Responsiveness in Hemodialysis Patients: A Cross-Sectional Study. Kidney Blood Press Res. 2019, 44, 354–361. [Google Scholar] [CrossRef]
- Torii, S.; Kobayashi, K.; Takahashi, M.; Katahira, K.; Goryo, K.; Matsushita, N.; Yasumoto, K.-I.; Fujii-Kuriyama, Y.; Sogawa, K. Magnesium deficiency causes loss of response to intermittent hypoxia in paraganglion cells. J. Biol. Chem. 2009, 284, 19077–19089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haase, V.H. Regulation of erythropoiesis by hypoxia-inducible factors. Blood Rev. 2013, 27, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, S.; Yamamoto, A.; Tomonaga, S.; Funaba, M.; Matsui, T. Magnesium deficiency induces the emergence of mast cells in the liver of rats. J. Nutr. Sci. Vitaminol. 2013, 59, 560–563. [Google Scholar] [CrossRef] [Green Version]
- Kurantsin-Mills, J.; Cassidy, M.M.; Stafford, R.E.; Weglicki, W.B. Marked alterations in circulating inflammatory cells during cardiomyopathy development in a magnesium-deficient rat model. Br. J. Nutr. 1997, 78, 845–855. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.D.; DeLeo, F.R. Neutrophil apoptosis and the resolution of infection. Immunol. Res. 2009, 43, 25–61. [Google Scholar] [CrossRef]
- Manz, M.G.; Boettcher, S. Emergency granulopoiesis. Nat. Rev. Immunol. 2014, 14, 302–314. [Google Scholar] [CrossRef]
- Panopoulos, A.D.; Zhang, L.; Snow, J.W.; Jones, D.M.; Smith, A.M.; El Kasmi, K.C.; Liu, F.; Goldsmith, M.A.; Link, D.C.; Murray, P.J.; et al. STAT3 governs distinct pathways in emergency granulopoiesis and mature neutrophils. Blood 2006, 108, 3682–3690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yvan-Charvet, L.; Ng, L.G. Granulopoiesis and Neutrophil Homeostasis: A Metabolic, Daily Balancing Act. Trends Immunol. 2019, 40, 598–612. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, K.; Rankin, S.M. CXCR4, the master regulator of neutrophil trafficking in homeostasis and disease. Eur. J. Clin. Investig. 2018, 48 (Suppl. 2), e12949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauseef, W.M.; Borregaard, N. Neutrophils at work. Nat. Immunol. 2014, 15, 602–611. [Google Scholar] [CrossRef]
- Rijkers, G.T.; Griffioen, A.W. Changes in free cytoplasmic magnesium following activation of human lymphocytes. Biochem. J. 1993, 289, 373–377. [Google Scholar] [CrossRef]
- Brandao, K.; Deason-Towne, F.; Perraud, A.L.; Schmitz, C. The role of Mg2+ in immune cells. Immunol. Res. 2013, 55, 261–269. [Google Scholar] [CrossRef]
- Hass, G.M.; Laing, G.H.; Galt, R.M.; McCreary, P.A. Recent Advances: Immunopathology of Magnesium Deficiency in Rats. Induction of Tumors; Incidence, Transmission and Prevention of Lymphoma-Leukemia. Magnes. Bull. 1981, 3, 217–228. [Google Scholar]
- Jasmin, G. Thymic lymphosarcoma in magnesium-deficient rats. Biol. Trace Elem. Res. 1979, 1, 217–228. [Google Scholar] [CrossRef]
- Averdunk, R.; Bippus, P.H.; Günther, T.; Merker, H.J. Development and properties of malignant lymphoma induced by magnesium deficiency in rats. J. Cancer Res. Clin. Oncol. 1982, 104, 63–73. [Google Scholar] [CrossRef]
- BOIS, P. Tumour of the thymus in magnesium-deficient rats. Nature 1964, 204, 1316. [Google Scholar] [CrossRef]
- Sahin, G.; Ertem, U.; Duru, F.; Birgen, D.; Yüksek, N. High prevelance of chronic magnesium deficiency in T cell lymphoblastic leukemia and chronic zinc deficiency in children with acute lymphoblastic leukemia and malignant lymphoma. Leuk. Lymphoma 2000, 39, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Merza, W.M.; majid, A.Y.; Daoud, M.S.; Jawad, A.M. Serum Magnesium Concentration in Patients with Leukemia and Lymphoma. Fac. Med. Baghdad. 2009, 51, 101. [Google Scholar]
- Guo, C.Y.; Halton, J.M.; Barr, R.D.; Atkinson, S.A. Hypomagnesemia associated with chemotherapy in patients treated for acute lymphoblastic leukemia: Possible mechanisms. Oncol. Rep. 2004, 11, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Afridi, H.I.; Kazi, T.G.; Talpur, F.N. Correlation of Calcium and Magnesium Levels in the Biological Samples of Different Types of Acute Leukemia Children. Biol. Trace Elem. Res. 2018, 186, 395–406. [Google Scholar] [CrossRef]
- Feske, S.; Skolnik, E.Y.; Prakriya, M. Ion channels and transporters in lymphocyte function and immunity. Nat. Rev. Immunol. 2012, 12, 532–547. [Google Scholar] [CrossRef] [Green Version]
- Sahni, J.; Scharenberg, A.M. TRPM7 ion channels are required for sustained phosphoinositide 3-kinase signaling in lymphocytes. Cell Metab. 2008, 8, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Krishnamoorthy, M.; Buhari, F.H.M.; Zhao, T.; Brauer, P.M.; Burrows, K.; Cao, E.Y.; Moxley-Paquette, V.; Mortha, A.; Zúñiga-Pflücker, J.C.; Treanor, B. The ion channel TRPM7 is required for B cell lymphopoiesis. Sci. Signal. 2018, 11, eaan2693. [Google Scholar] [CrossRef] [Green Version]
- Li, F.Y.; Chaigne-Delalande, B.; Su, H.; Uzel, G.; Matthews, H.; Lenardo, M.J. XMEN disease: A new primary immunodeficiency affecting Mg2+ regulation of immunity against Epstein-Barr virus. Blood 2014, 123, 2148–2152. [Google Scholar] [CrossRef] [Green Version]
- Ravell, J.C.; Matsuda-Lennikov, M.; Chauvin, S.D.; Zou, J.; Biancalana, M.; Deeb, S.J.; Price, S.; Su, H.C.; Notarangelo, G.; Jiang, P.; et al. Defective glycosylation and multisystem abnormalities characterize the primary immunodeficiency XMEN disease. J. Clin. Investig. 2020, 130, 507–522. [Google Scholar] [CrossRef] [Green Version]
- Blommaert, E.; Péanne, R.; Cherepanova, N.A.; Rymen, D.; Staels, F.; Jaeken, J.; Race, V.; Keldermans, L.; Souche, E.; Corveleyn, A.; et al. Mutations in MAGT1 lead to a glycosylation disorder with a variable phenotype. Proc. Natl. Acad. Sci. USA 2019, 116, 9865–9870. [Google Scholar] [CrossRef] [Green Version]
- Ravell, J.C.; Chauvin, S.D.; He, T.; Lenardo, M. An Update on XMEN Disease. J. Clin. Immunol. 2020, 40, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Matsuda-Lennikov, M.; Biancalana, M.; Zou, J.; Ravell, J.C.; Zheng, L.; Kanellopoulou, C.; Jiang, P.; Notarangelo, G.; Jing, H.; Masutani, E.; et al. Magnesium transporter 1 (MAGT1) deficiency causes selective defects in N-linked glycosylation and expression of immune-response genes. J. Boil. Chem. 2019, 294, 13638–13656. [Google Scholar] [CrossRef] [PubMed]
- Chaigne-Delalande, B.; Li, F.-Y.; O’Connor, G.M.; Lukacs, M.J.; Jiang, P.; Zheng, L.; Shatzer, A.; Biancalana, M.; Pittaluga, S.; Matthews, H.F.; et al. Mg2+ Regulates Cytotoxic Functions of NK and CD8 T Cells in Chronic EBV Infection Through NKG2D. Science 2013, 341, 186–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shechter, M.; Merz, N.B.; Rude, R.K.; Labrador, M.J.P.; Meisel, S.R.; Shah, P.K.; Kaul, S. Low intracellular magnesium levels promote platelet-dependent thrombosis in patients with coronary artery disease. Am. Heart J. 2000, 140, 212–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravn, H.B.; Kristensen, S.D.; Vissinger, H.; Husted, S.E. Magnesium inhibits human platelets. Blood Coagul. Fibrinolysis 1996, 7, 241–244. [Google Scholar] [CrossRef]
- Nadler, J.L.; Malayan, S.; Luong, H.; Shaw, S.; Natarajan, R.D.; Rude, R.K. Intracellular free magnesium deficiency plays a key role in increased platelet reactivity in type II diabetes mellitus. Diabetes Care 1992, 15, 835–841. [Google Scholar] [CrossRef]
- Takaya, J.; Yamato, F.; Higashino, H.; Kobayashi, Y. Relationship of intracellular magnesium of cord blood platelets to birth weight. Metabolism 2004, 53, 1544–1547. [Google Scholar] [CrossRef]
- Oliveira, D.C.; Nogueira-Pedro, A.; Santos, E.W.; Hastreiter, A.; Silva, G.B.; Borelli, P.; Fock, R.A. A review of select minerals influencing the haematopoietic process. Nutr. Res. Rev. 2018, 31, 267–280. [Google Scholar] [CrossRef]

{kind=link}
| Reference | Main Findings | Source |
|---|---|---|
| Mesenchymal Stem Cells and Osteoblasts | ||
| [31] | Increase in Mg2+ enhanced expression of RUNX2, OSTERIX, and OSTEOCALCIN genes, increased the levels cyclin D1 and PCNA proteins, and induced the activation of Notch signaling. | BMSCs |
| [32] | Higher Mg2+ concentration led to upregulation of RUNX2, BMP2, ALP, OPN, and ColI and increased PI3K/Akt signaling pathway. | Osteoblasts from animal model |
| [33] | Higher Mg2+ levels triggered PI3K phosphorylation via TRPM7 and led to migration of osteoblasts. | hFOB1.19 (human osteoblast cells) |
| [34] | Increase in Mg2+ led to changes in the expression of TRPM7 and in homeostasis of other important metal ions for bone tissue, inhibiting ALP activity in osteoblasts. | SaOS-2 cells and human osteoblasts |
| [35] | Decreased osteoblast number. | Bone marrow from mice |
| Endothelial Cells | ||
| [36] | Lower Mg2+ levels impaired proliferation and migration, and increased IL-6, NOS activity, and VCAM expression. | 1G11 cells |
| [37] | Mg2+ deficiency triggered higher NF-ĸB activation, and increased IL-8, RANTES, and GM-CSF. | HUVEC cells |
| [38] | Lower Mg2+ concentration decreased proliferation taxa and downregulation of TRPM7. | HMEC cells |
| [39] | Mg2+ deficiency increased ROS, PGE2, TNFα, and IL-1β levels, triggered changes in mRNA of ICAM-1 and VCAM-1, decreased viability and cell proliferation, triggered changes in the endothelial permeability, and influenced integrity of the endothelial barrier through the S1P1–Rac1 pathway. | ECs from animal model |
| [40] | Mg2+ supplementation improved endothelial function. | Clinical trial |
| [41] | Daily Mg2+ supplement of 350 mg for 24 weeks led to no improvement in soluble vascular cell adhesion molecule (sVCAM)-1, soluble intercellular adhesion molecule (sICAM)-1, and soluble endothelial selectin (sE-selectin), along with no change in FMD markers of endothelial function or inflammatory parameters. | Clinical trial |
| Macrophages | ||
| [42] | Mg2+ deficiency increased TNF-α and IL-1 mRNA levels with the involvement of Ca2+ signaling pathways. | Rat alveolar macrophages |
| [43] | Low Mg2+ concentration increased NF-κB activity and changed HMGB1 expression. | RAW264.7 cells |
| [44] | Increased Mg2+ concentration inhibited the nuclear translocation and phosphorylation of NF-κB and led to a rise in basal IĸBα levels. | Human PBMCs |
| Erythropoiesis | ||
| [45] | Mg2+ deficiency decreased plasma level, and led to faster aging of RBCs, as well as reticulocytosis. | Animal model |
| [46] | Mg2+ deficiency increased Fe absorption and concentration, but reduced the number of RBCs. | Animal model |
| [47] | Inverse association between Mg2+ intake and anemia. | Cross-sectional study |
| [48] | Association between increased serum Mg2+ and decreased risk of anemia in women, dependent on ferritin levels. | Cross-sectional study |
| [49] | Decreased levels of serum ferritin and Mg2+ were associated with anemia in pregnant women. | Cross-sectional study |
| [50] | Mg2+ supplementation raised the hemoglobin levels and counts of erythrocytes. | Clinical trial |
| Granulopoiesis | ||
| [51] | Boost in circulating eosinophils during Mg2+ deficiency. | Experimental model |
| [52] | Mg2+ deficiency impaired mast-cell functions. | Experimental model |
| [53] | Mg2+ deficiency increased the mast cells in the bone marrow. | Experimental model |
| [54] | Increase in the number and activity of PMNs during Mg2+ deficiency. | Experimental model |
| [26] | Lower Mg2+ levels triggered granulocytic differentiation and changes in proteins related to cell-cycle control. | HL-60 cells |
| [15] | Mg2+ deficiency resulted in hypercellular bone marrow, as well as a greater number of granulocytic cell; one animal also developed leukemia with granulocytic infiltrate in several organs. | Experimental model |
| Thrombopoiesis | ||
| [55] | Higher serum Mg2+ levels were associated with increased platelet numbers and lower risk of development of thrombocytopenia. | Cross-sectional study |
| [56] | Mg2+ deficiency impaired the number and shape of megakaryocytes from bone marrow. | Experimental model |
| [16] | Changes in the TRPM7 channel and Mg2+ homeostasis in megakaryocytes triggered macrothrombocytopenia, altering the activity of the NMMIIA and the cytoskeleton, affecting the maturation of platelets in the bone marrow. | Experimental model |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, F.d.S.; Fock, R.A. A Review of the Action of Magnesium on Several Processes Involved in the Modulation of Hematopoiesis. Int. J. Mol. Sci. 2020, 21, 7084. https://doi.org/10.3390/ijms21197084
Lima FdS, Fock RA. A Review of the Action of Magnesium on Several Processes Involved in the Modulation of Hematopoiesis. International Journal of Molecular Sciences. 2020; 21(19):7084. https://doi.org/10.3390/ijms21197084
Chicago/Turabian StyleLima, Fabiana da Silva, and Ricardo Ambrósio Fock. 2020. "A Review of the Action of Magnesium on Several Processes Involved in the Modulation of Hematopoiesis" International Journal of Molecular Sciences 21, no. 19: 7084. https://doi.org/10.3390/ijms21197084