Hippocampal 2-Arachidonoyl Glycerol Signaling Regulates Time-of-Day- and Stress-Dependent Effects on Rat Short-Term Memory



Abstract
:1. Introduction
2. Results
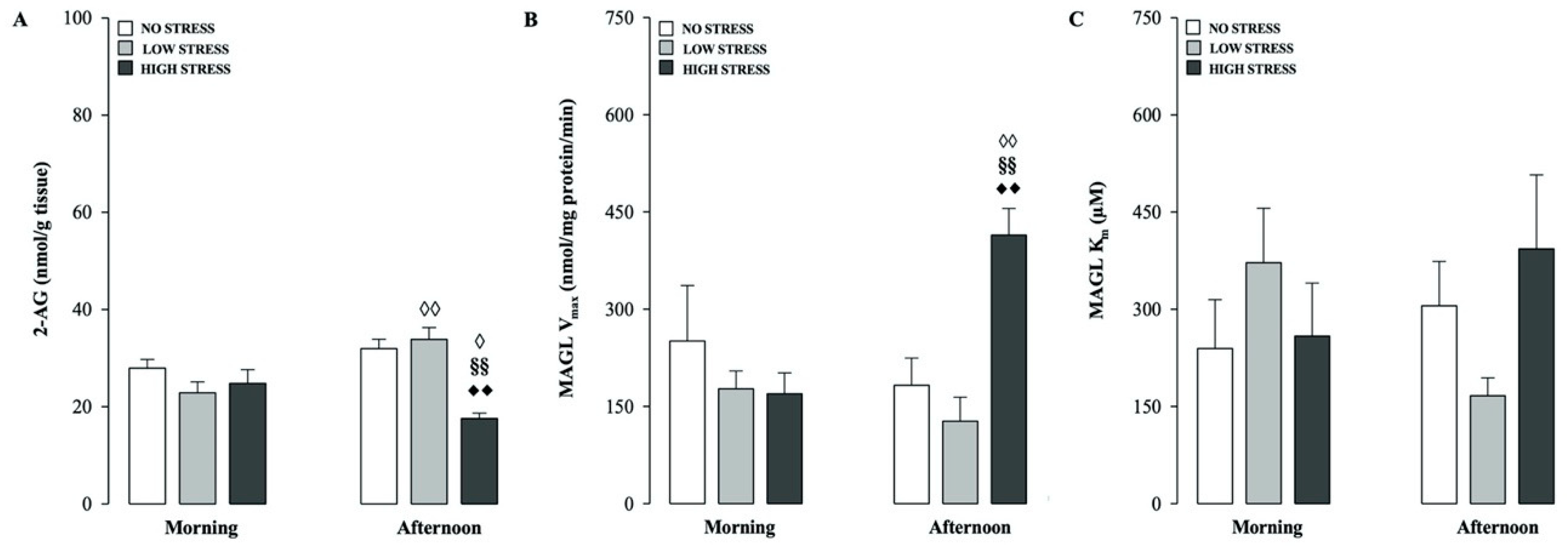
2.1. Effects of Different Stress Intensities on Hippocampal 2-AG Levels and MAGL Hydrolytic Activity in Rats Tested in the Morning or Afternoon
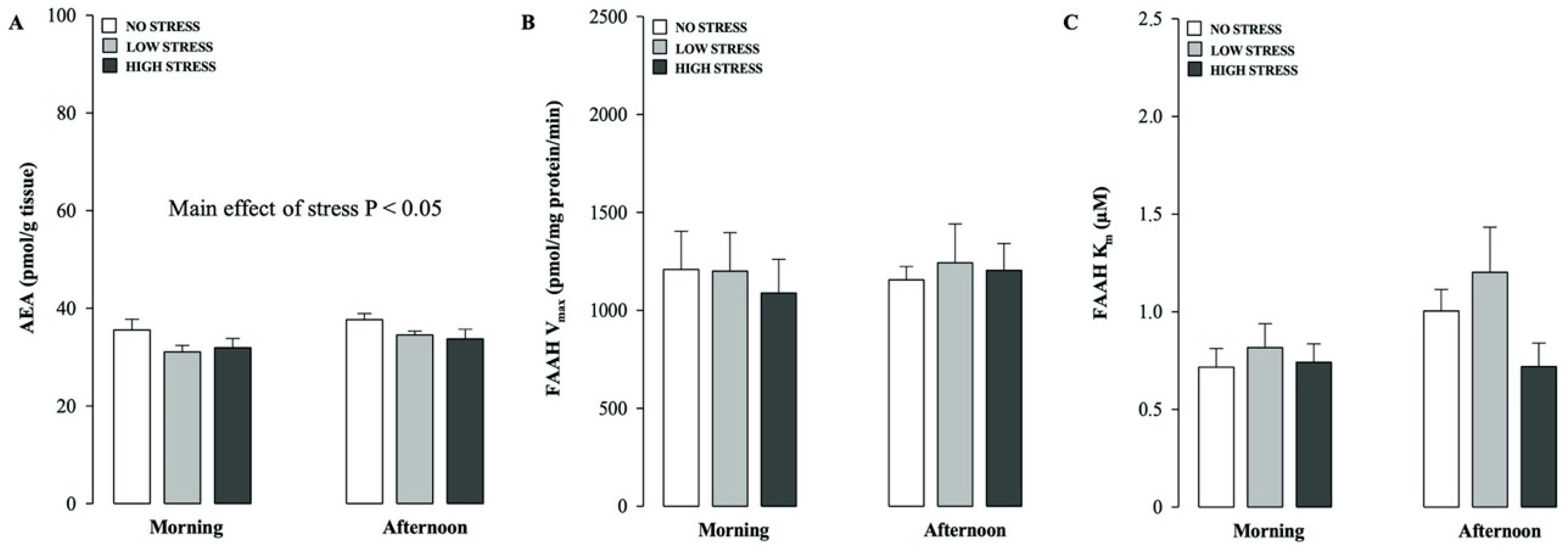
2.2. Effects of Different Stress Intensities on Hippocampal AEA Levels and FAAH Hydrolytic Activity in Rats Tested in the Morning or Afternoon
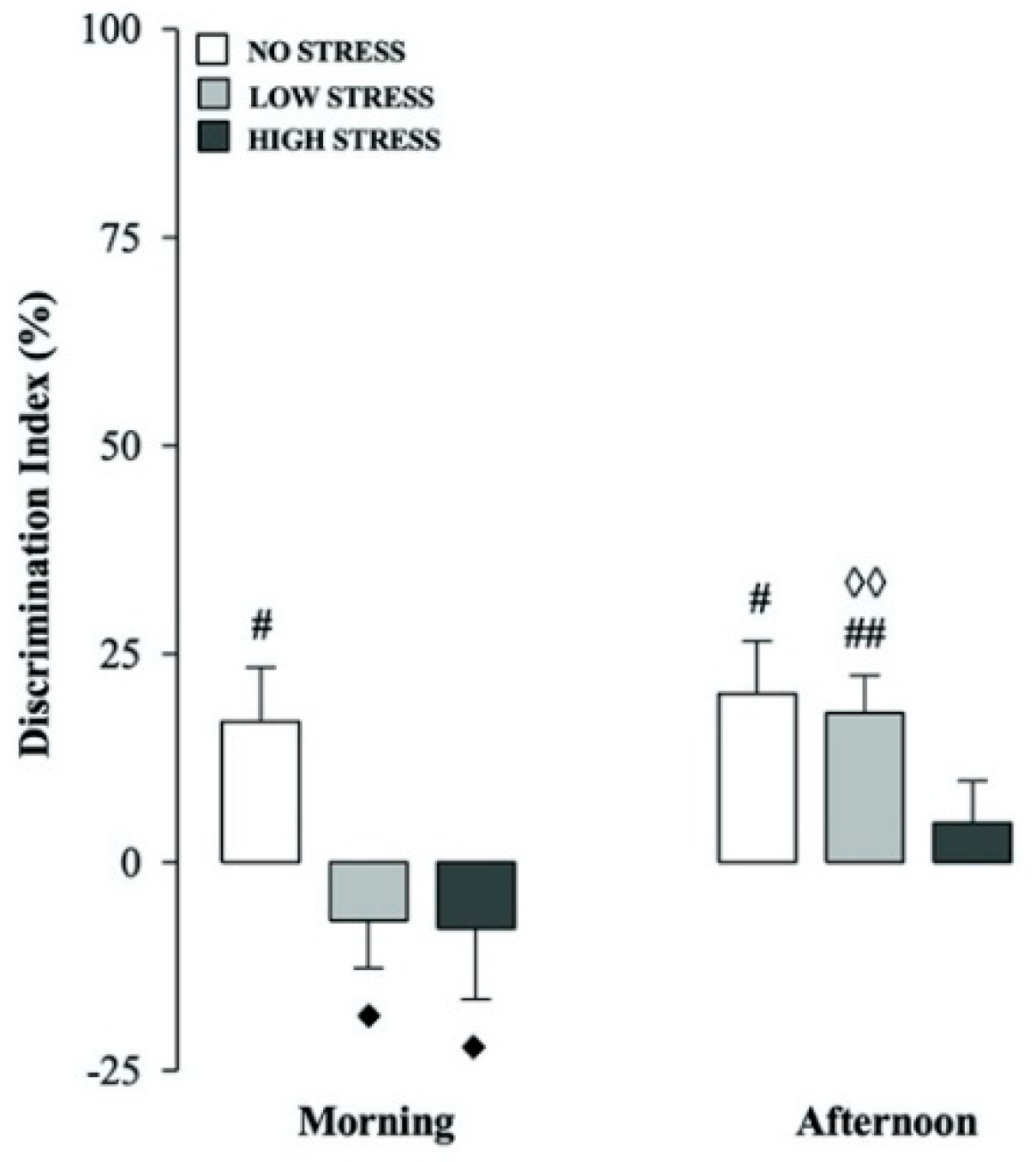
2.3. Stress Intensity and Time-of-Day Effects on Short-Term Recognition Memory
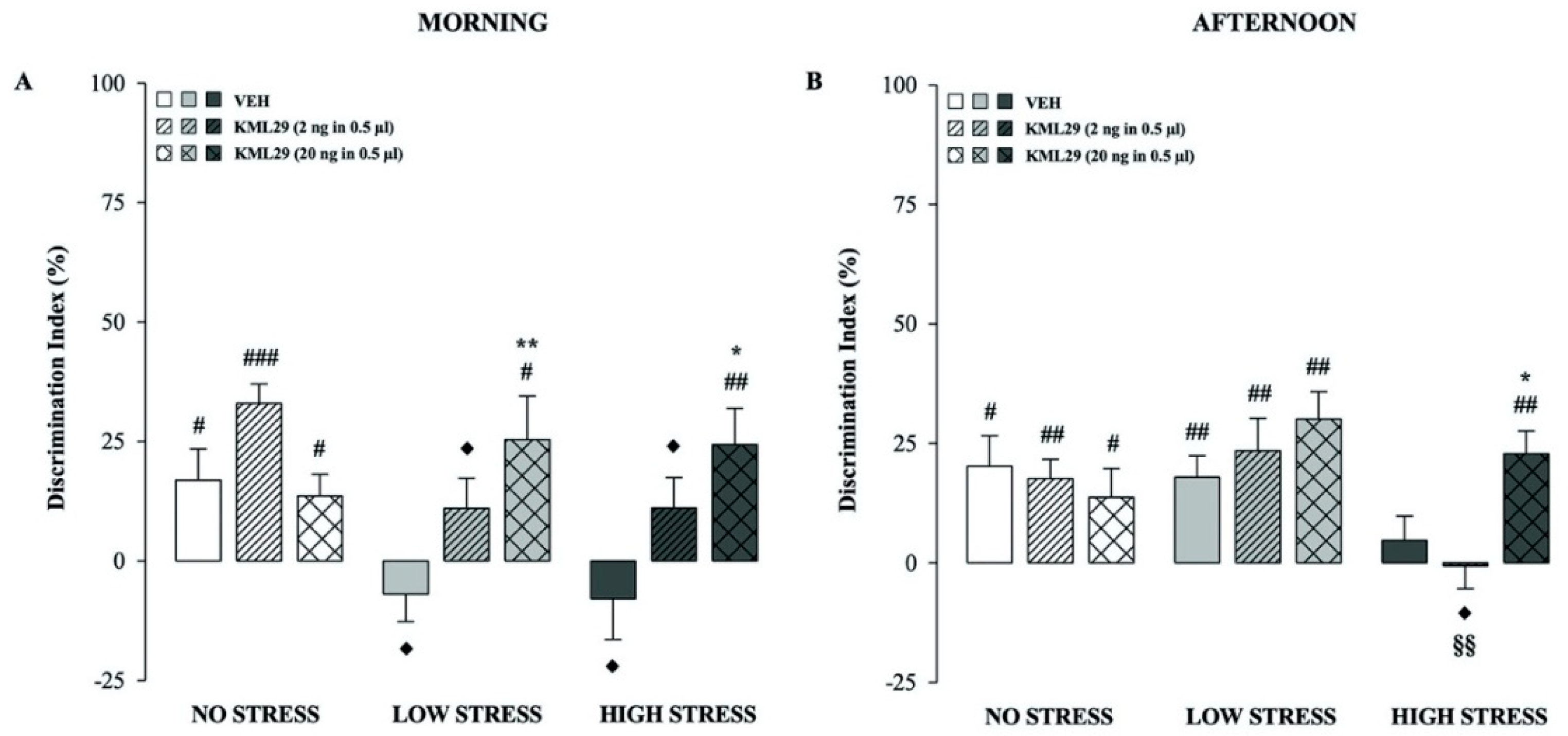
2.4. Effects of the 2-AG Hydrolysis Inhibitor KML29 on Hippocampal Modulation of Short-Term Recognition Memory Performance in the No, Low and High Stress Condition Groups Tested in the Morning
2.5. Effects of the 2-AG Hydrolysis Inhibitor KML29 on Hippocampal Modulation of Short-Term Recognition Memory Performance in the No, Low and High Stress Condition Groups Tested in the Afternoon
3. Discussion
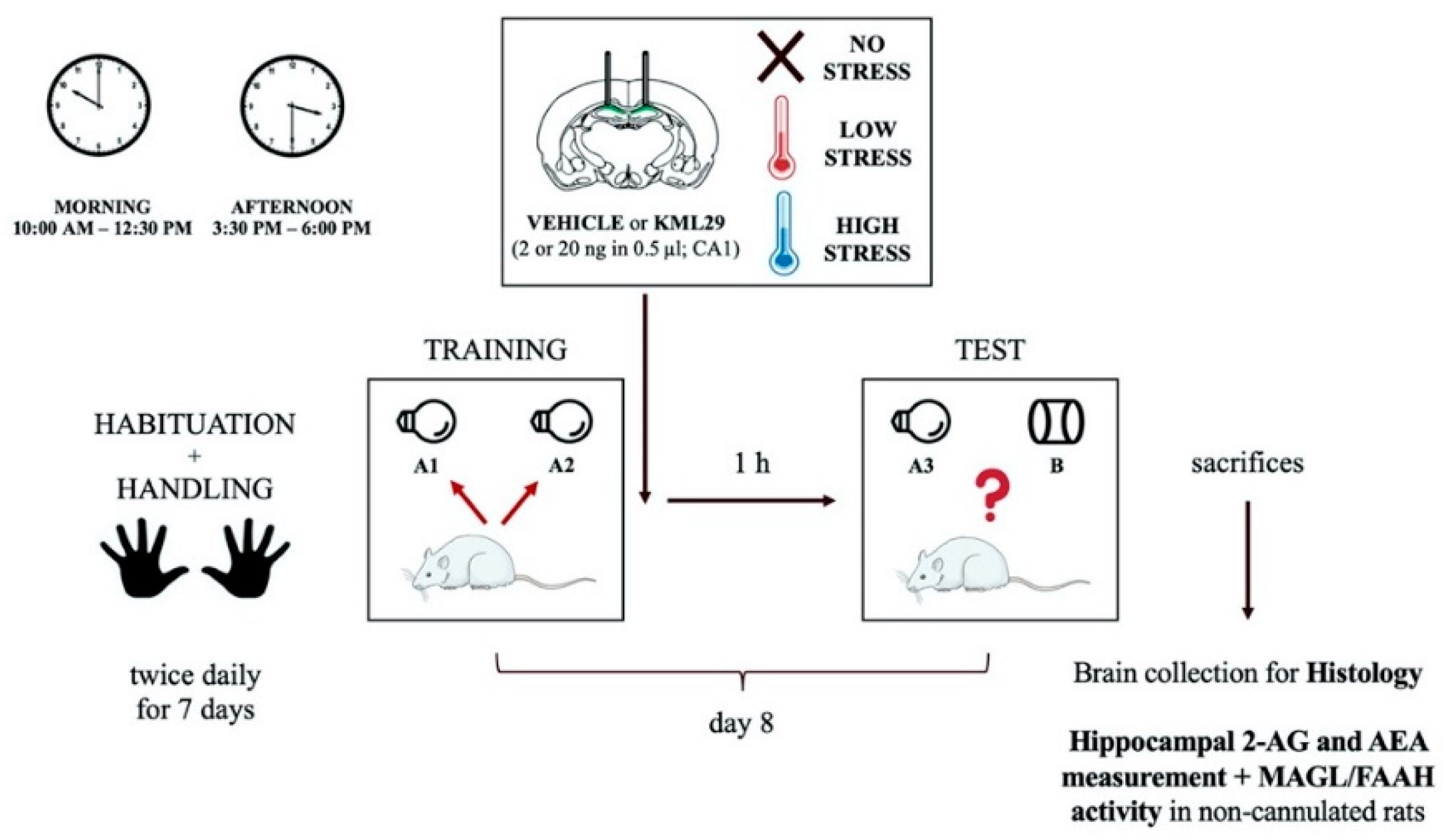
4. Materials and Methods
4.1. Animal Care and Use
4.2. Surgery
4.3. Drug Administration
4.4. Behavioral Procedures
4.5. Endocannabinoid Extraction and Analysis
4.6. Membrane Preparation
4.7. MAGL Activity Assay
4.8. FAAH Activity Assay
4.9. Histology
4.10. Data and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 2-AG | 2-arachidonoyl glycerol |
| AEA | Anandamide |
| ANOVA | Analysis of variance |
| BLA | Basolateral complex of the amygdala |
| CB1 | Cannabinoid type-1 |
| CB2 | Cannabinoid type-2 |
| CRH | Corticotropin-releasing hormone |
| CRHR1 | Corticotropin-releasing hormone receptors 1 |
| FAAH | Fatty acid amide hydrolase |
| HPA | Hypothalamic–pituitary–adrenal axis |
| KML29 | 1,1,1,3,3,3-Hexafluoropropan-2-yl 4-[bis(1,3-benzodioxol-5-yl)-hydroxymethyl]piperidine-1- carboxylate |
| MAGL | Monoacylglycerol lipase |
| mPFC | Medial prefrontal cortex |
| SEM | Standard error of the mean |
References
- Vogel, S.; Schwabe, L. Learning and memory under stress: Implications for the classroom. Npj Sci. Learn. 2016, 1, 16011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, B. Post-traumatic stress disorder: Symptom profiles in men and women. Curr. Med. Res. Opin. 2003, 19, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Bower, G.H.; Sivers, H. Cognitive impact of traumatic events. Dev Psychopathol. 1998, 10, 625–653. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. The Brain on Stress: Toward an Integrative Approach to Brain, Body, and Behavior. Perspect. Psychol. Sci. 2013, 8, 673–675. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S. Stress and Hippocampal Plasticity. Annu. Rev. Neurosci. 1999, 22, 105–122. [Google Scholar] [CrossRef] [Green Version]
- De Kloet, E.R.; Meijer, O.C.; de Nicola, A.F.; de Rijk, R.H.; Joëls, M. Importance of the brain corticosteroid receptor balance in metaplasticity, cognitive performance and neuro-inflammation. Front. Neuroendocrinol. 2018, 49, 124–145. [Google Scholar] [CrossRef]
- Broadbent, N.J.; Gaskin, S.; Squire, L.R.; Clark, R.E. Object recognition memory and the rodent hippocampus. Learn. Mem. 2010, 17, 794–800. [Google Scholar] [CrossRef] [Green Version]
- Hill, M.N.; Patel, S.; Campolongo, P.; Tasker, J.G.; Wotjak, C.T.; Bains, J.S. Functional Interactions between Stress and the Endocannabinoid System: From Synaptic Signaling to Behavioral Output. J. Neurosci. 2010, 30, 14980–14986. [Google Scholar] [CrossRef]
- Colangeli, R.; Pierucci, M.; Benigno, A.; Campiani, G.; Butini, S.; Di Giovanni, G. The FAAH inhibitor URB597 suppresses hippocampal maximal dentate afterdischarges and restores seizure-induced impairment of short and long-term synaptic plasticity. Sci. Rep. 2017, 7, 11152. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, L.A.; Lolait, J.S.; Brownstein, J.M.; Young, C.A.; Bonner, I.T. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990, 346, 561–564. [Google Scholar] [CrossRef]
- Sugiura, T.; Kondo, S.; Sukagawa, A.; Nakane, S.; Shinoda, A.; Itoh, K.; Yamashita, A.; Waku, K. 2-Arachidonoylglycerol: A possible endogenous cannabinoid receptor ligand in brain. Biochem. Biophys. Res. Commun. 1995, 215, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef] [PubMed]
- Kano, M.; Ohno-Shosaku, T.; Hashimotodani, Y.; Uchigashima, M.; Watanabe, M. Endocannabinoid-Mediated Control of Synaptic Transmission. Physiol. Rev. 2009, 89, 309–380. [Google Scholar] [CrossRef] [PubMed]
- Ohno-Shosaku, T.; Kano, M. Endocannabinoid-mediated retrograde modulation of synaptic transmission. Curr. Opin. Neurobiol. 2014, 29, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Dinh, T.P.; Carpenter, D.; Leslie, F.M.; Freund, T.F.; Katona, I.; Sensi, S.L.; Kathuria, S.; Piomelli, D. Brain monoglyceride lipase participating in endocannabinoid inactivation. Proc. Natl. Acad. Sci. USA 2002, 99, 10819–10824. [Google Scholar] [CrossRef] [Green Version]
- Cravatt, B.F.; Demarest, K.; Patricelli, M.P.; Bracey, M.H.; Giang, D.K.; Martin, B.R.; Lichtman, A.H. Supersensitivity to anandamide and enhanced endogenous cannabinoid signaling in mice lacking fatty acid amide hydrolase. Proc. Natl. Acad. Sci. USA 2001, 98, 9371–9376. [Google Scholar] [CrossRef] [Green Version]
- Vaughn, L.K.; Denning, G.; Stuhr, K.L.; de Wit, H.; Hill, M.N.; Hillard, C.J. Endocannabinoid signalling: Has it got rhythm? Br. J. Pharmacol. 2010, 160, 530–543. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.; Roelke, C.T.; Rademacher, D.J.; Cullinan, W.E.; Hillard, C.J. Endocannabinoid Signaling Negatively Modulates Stress-Induced Activation of the Hypothalamic-Pituitary-Adrenal Axis. Endocrinology 2004, 145, 5431–5438. [Google Scholar] [CrossRef]
- Di, S.; Malcher-Lopes, R.; Halmos, K.C.; Tasker, J.G. Nongenomic glucocorticoid inhibition via endocannabinoid release in the hypothalamus: A fast feedback mechanism. J. Neurosci. 2003, 23, 4850–4857. [Google Scholar] [CrossRef] [Green Version]
- Morena, M.; Patel, S.; Bains, J.S.; Hill, M.N. Neurobiological Interactions Between Stress and the Endocannabinoid System. Neuropsychopharmacology 2016, 41, 80–102. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, H.C.; Leggett, J.D.; Wood, S.A.; Castrique, E.S.; Kershaw, Y.M.; Lightman, S.L. Regulation of the hypothalamic-pituitary-adrenal axis circadian rhythm by endocannabinoids is sexually diergic. Endocrinology 2010, 151, 3720–3727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morena, M.; De Castro, V.; Gray, J.M.; Palmery, M.; Trezza, V.; Roozendaal, B.; Hill, M.N.; Compolongo, P. Training-Associated Emotional Arousal Shapes Endocannabinoid Modulation of Spatial Memory Retrieval in Rats. J. Neurosci. 2015, 35, 13962–13974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morena, M.; Leitl, K.D.; Vecchiarelli, H.A.; Gray, J.M.; Campolongo, P.; Hill, M.N. Emotional arousal state influences the ability of amygdalar endocannabinoid signaling to modulate anxiety. Neuropharmacology 2016, 111, S397–S402. [Google Scholar] [CrossRef]
- Morena, M.; Aukema, R.J.; Leitl, K.D.; Rashid, A.J.; Vecchiarelli, H.A.; Josselyn, S.A.; Hil, M.N. Upregulation of Anandamide Hydrolysis in the Basolateral Complex of Amygdala Reduces Fear Memory Expression and Indices of Stress and Anxiety. J. Neurosci. 2019, 39, 1275–1292. [Google Scholar] [CrossRef] [PubMed]
- Santori, A.; Colucci, P.; Mancini, G.F.; Morena, M.; Palmery, M.; Trezza, V.; Puglisi-Allegra, S.; Hill, M.N.; Compolongo, P. Anandamide modulation of circadian- and stress-dependent effects on rat short-term memory. Psychoneuroendocrinology 2019, 108, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.N.; McLaughlin, R.J.; Pan, B.; Fitzgerald, M.L.; Roberts, C.J.; Lee, T.T.; Karatsoreos, I.N.; Mackie, N.; Viau, V.; Pickel, V.M.; et al. Recruitment of prefrontal cortical endocannabinoid signaling by glucocorticoids contributes to termination of the stress response. J. Neurosci. 2011, 31, 10506–10515. [Google Scholar] [CrossRef] [PubMed]
- Antony, J.M.; McDonald, A.C.; Noorbhaksh, F.; Guthrie, N.; Evans, M. Changing the tone of clinical study design in the cannabis industry. Transl. Neurosci. 2020, 11, 4–9. [Google Scholar] [CrossRef] [Green Version]
- Akirav, I. Cannabinoids and glucocorticoids modulate emotional memory after stress. Neurosci. Biobehav. Rev. 2013, 37, 2554–2563. [Google Scholar] [CrossRef]
- Gray, J.M.; Vecchiarelli, H.A.; Morena, M.; Lee, T.T.Y.; Hermanson, D.J.; Kim, A.B.; McLaughlin, R.J.; Hassan, H.I.; Kühne, C.; Wotjak, C.T.; et al. Corticotropin-releasing hormone drives anandamide hydrolysis in the amygdala to promote anxiety. J. Neurosci. 2015, 35, 3879–3892. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, R.J.; Hill, M.N.; Bambico, F.R.; Stuhr, K.L.; Gobbi, G.; Hillard, C.J.; Gorzalka, B.B. Prefrontal cortical anandamide signaling coordinates coping responses to stress through a serotonergic pathway. Eur. Neuropsychopharmacol. 2012, 22, 664–671. [Google Scholar] [CrossRef] [Green Version]
- Evanson, N.K.; Tasker, J.G.; Hill, M.N.; Hillard, C.J.; Herman, J.P. Fast feedback inhibition of the HPA axis by glucocorticoids is mediated by endocannabinoid signaling. Endocrinology 2010, 151, 4811–4819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Hill, M.N.; Zhang, L.; Gorzalka, B.B.; Hillard, C.J.; Alger, B.E. Acute restraint stress enhances hippocampal endocannabinoid function via glucocorticoid receptor activation. J. Psychopharmacol. 2012, 26, 56–70. [Google Scholar] [CrossRef]
- Baggelaar, M.P.; Maccarrone, M.; van der Stelt, M. 2-Arachidonoylglycerol: A signaling lipid with manifold actions in the brain. Prog. Lipid. Res. 2018, 71, 1–17. [Google Scholar] [CrossRef]
- Kim, E.J.; Pellman, B.; Kim, J.J. Stress effects on the hippocampus: A critical review. Learn. Mem. 2015, 22, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Donley, M.P.; Schulkin, J.; Rosen, J.B. Glucocorticoid receptor antagonism in the basolateral amygdala and ventral hippocampus interferes with long-term memory of contextual fear. Behav. Brain Res. 2005, 164, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Campolongo, P.; Morena, M.; Scaccianoce, S.; Trezza, V.; Chiarotti, F.; Schelling, G.; Cuomo, V.; Roozendaal, B. Novelty-induced emotional arousal modulates cannabinoid effects on recognition memory and adrenocortical activity. Neuropsychopharmacology 2013, 38, 1276–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morena, M.; Campolongo, P. The endocannabinoid system: An emotional buffer in the modulation of memory function. Neurobiol. Learn. Mem. 2014, 112, 30–43. [Google Scholar] [CrossRef]
- Atsak, P.; Hauer, D.; Campolongo, P.; Schelling, G.; McGaugh, J.L.; Roozendaal, B. Glucocorticoids interact with the hippocampal endocannabinoid system in impairing retrieval of contextual fear memory. Proc. Natl. Acad. Sci. USA 2012, 109, 3504–3509. [Google Scholar] [CrossRef] [Green Version]
- Spiga, F.; Walker, J.J.; Terry, J.R.; Lightman, S.L. HPA Axis-Rhythms. Compr. Physiol. 2014, 4, 1273–1298. [Google Scholar]
- Menon, J.M.L.; Nolten, C.; Achterberg, E.J.M.; Joosten, R.N.J.M.A.; Dematteis, M.; Feenstra, M.G.P.; Drinkenburg, W.H.; Leeenars, S.H.C. Brain Microdialysate Monoamines in Relation to Circadian Rhythms, Sleep, and Sleep Deprivation—A Systematic Review, Network Meta-analysis, and New Primary Data. J. Circadian Rhythms. 2019, 17, 1. [Google Scholar] [CrossRef] [Green Version]
- Morena, M.; Roozendaal, B.; Trezza, V.; Ratano, P.; Peloso, A.; Hauer, D.; Atsak, P.; Trabace, L.; Cuomo, V.; McCaugh, J.L.; et al. Endogenous cannabinoid release within prefrontal-limbic pathways affects memory consolidation of emotional training. Proc. Natl. Acad. Sci. USA 2014, 111, 18333–18338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campolongo, P.; Roozendaal, B.; Trezza, V.; Hauer, D.; Schelling, G.; McGaugh, J.L.; Cuomo, V. Endocannabinoids in the rat basolateral amygdala enhance memory consolidation and enable glucocorticoid modulation of memory. Proc. Natl. Acad. Sci. USA 2009, 106, 4888–4893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, A.J.; Mott, D.D. Functional neuroanatomy of amygdalohippocampal interconnections and their role in learning and memory. J. Neurosci. Res. 2017, 95, 797–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, M.; Biala, G. The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cogn. Process. 2012, 13, 93–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, M.W. Hippocampal and perirhinal functions in recognition memory. Nat. Rev. Neurosci. 2008, 9, 405. [Google Scholar] [CrossRef]
- Eichenbaum, H.; Yonelinas, A.P.; Ranganath, C. The Medial Temporal Lobe and Recognition Memory. Annu. Rev. Neurosci. 2007, 30, 123–152. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira Alvares, L.; Genro, B.P.; Diehl, F.; Quillfeldt, J.A. Differential role of the hippocampal endocannabinoid system in the memory consolidation and retrieval mechanisms. Neurobiol. Learn. Mem. 2008, 90, 1–9. [Google Scholar] [CrossRef]
- Niederhoffer, N.; Hansen, H.H.; Fernandez-Ruiz, J.J.; Szabo, B. Effects of cannabinoids on adrenaline release from adrenal medullary cells. Br. J. Pharmacol. 2001, 134, 1319–1327. [Google Scholar] [CrossRef]
- Richter, H.; Teixeira, F.M.; Ferreira, S.G.; Kittel, Á.; Köfalvi, A.; Sperlágh, B. Presynaptic α2-adrenoceptors control the inhibitory action of presynaptic CB1 cannabinoid receptors on prefrontocortical norepinephrine release in the rat. Neuropharmacology 2012, 63, 784–797. [Google Scholar] [CrossRef]
- Hill, M.N.; McEwen, B.S. Involvement of the endocannabinoid system in the neurobehavioural effects of stress and glucocorticoids. Prog. Neuropsychopharmacol. Biol. Psychiatry 2010, 34, 791–797. [Google Scholar] [CrossRef] [Green Version]
- Roozendaal, B.; Brunson, K.L.; Holloway, B.L.; McGaugh, J.L.; Baram, T.Z. Involvement of stress-released corticotropin-releasing hormone in the basolateral amygdala in regulating memory consolidation. Proc. Natl. Acad. Sci. USA 2002, 99, 13908–13913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlo Pich, E.; Lorang, M.; Yeganeh, M.; Rodriguez de Fonseca, F.; Raber, J.; Koob, G.F.; Weiss, F. Increase of extracellular corticotropin-releasing factor-like immunoreactivity levels in the amygdala of awake rats during restraint stress and ethanol withdrawal as measured by microdialysis. J. Neurosci. 1995, 15, 5439–5447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, M.N.; Karatsoreos, I.N.; Hillard, C.J.; McEwen, B.S. Rapid elevations in limbic endocannabinoid content by glucocorticoid hormones in vivo. Psychoneuroendocrinology 2010, 35, 1333–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankord, R.; Herman, J.P. Limbic Regulation of Hypothalamo-Pituitary-Adrenocortical Function during Acute and Chronic Stress. Ann. N. Y. Acad. Sci. 2008, 1148, 64–73. [Google Scholar] [CrossRef]
- Qi, M.; Morena, M.; Vecchiarelli, H.A.; Hill, M.N.; Schriemer, D.C. A robust capillary liquid chromatography/tandem mass spectrometry method for quantitation of neuromodulatory endocannabinoids. Rapid. Commun. Mass. Spectrom. 2015, 29, 1889–1897. [Google Scholar] [CrossRef]
- Schneider, A.M.; Simson, P.E. NAN-190 potentiates the impairment of retention produced by swim stress. Pharmacol. Biochem. Behav. 2007, 87, 73–80. [Google Scholar] [CrossRef]
- Hill, M.N.; McLaughlin, R.J.; Morrish, A.C.; Viau, V.; Floresco, S.B.; Hillard, C.J.; Gorzalka, B.B. Suppression of amygdalar endocannabinoid signaling by stress contributes to activation of the hypothalamic-pituitary-adrenal axis. Neuropsychopharmacology 2009, 34, 2733–2745. [Google Scholar]
- Rademacher, D.J.; Meier, S.E.; Shi, L.; Ho, W.-S.V.; Jarrahian, A.; Hillard, C.J. Effects of acute and repeated restraint stress on endocannabinoid content in the amygdala, ventral striatum, and medial prefrontal cortex in mice. Neuropharmacology 2008, 54, 108–116. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Roozendaal, B.; Castello, N.A.; Vedana, G.; Barsegyan, A.; McGaugh, J.L. Noradrenergic activation of the basolateral amygdala modulates consolidation of object recognition memory. Neurobiol. Learn. Mem. 2008, 90, 576–579. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morning | Afternoon | |||||
|---|---|---|---|---|---|---|
| Total Object Exploration Time | Number of Crossings | Number of Rearings | Total Object Exploration Time | Number of Crossings | Number of Rearings | |
| NO STRESS | ||||||
| VEHICLE | 59.8 ± 3.5 | 45.5 ± 1.6 | 44.8 ± 4.2 | 50.4 ± 4.0 | 43.0 ± 3.3 | 32.0 ± 2.2 |
| KML 2 ng | 66.7 ± 3.9 | 52.4 ± 3.4 | 39.4 ± 2.9 | 54.2 ± 3.5 | 48.0 ± 3.4 | 41.4 ± 3.2 |
| KML 20 ng | 61.0 ± 3.5 | 48.8 ± 1.8 | 45.8 ± 2.8 | 58.8 ± 3.8 | 43.5 ± 3.9 | 37.0 ± 2.5 |
| LOW STRESS | ||||||
| VEHICLE | 80.0 ± 11.7 | 51.1 ± 2.8 | 52.0 ± 4.8 | 53.1 ± 3.0 | 42.5 ± 2.8 | 35.2 ± 2.5 |
| KML 2 ng | 67.5 ± 8.1 | 59.2 ± 1.4 | 49.2 ± 3.9 | 56.9 ± 3.5 | 42.0 ± 2.1 | 37.6 ± 3.3 |
| KML 20 ng | 62.7 ± 6.6 | 49.1 ± 3.2 | 47.8 ± 3.7 | 54.2 ± 2.8 | 46.0 ± 2.2 | 37.1 ± 2.2 |
| HIGH STRESS | ||||||
| VEHICLE | 62.8 ± 7.6 | 52.5 ± 4.7 | 46.3 ± 5.0 | 56.9 ± 3.3 | 46.1 ± 2.9 | 40.3 ± 2.8 |
| KML 2 ng | 62.3 ± 7.0 | 51.0 ± 3.6 | 43.1 ± 4.3 | 66.3 ± 5.5 | 48.8 ± 2.9 | 40.9 ± 2.3 |
| KML 20 ng | 63.6 ± 5.8 | 49.5 ± 2.6 | 39.8 ± 2.6 | 53.0 ± 3.1 | 48.2 ± 4.1 | 40.7 ± 3.3 |
| Morning | Afternoon | |||||
|---|---|---|---|---|---|---|
| Total Object Exploration Time | Number of Crossings | Number of Rearings | Total Object Exploration Time | Number of Crossings | Number of Rearings | |
| NO STRESS | ||||||
| VEHICLE | 45.1 ± 3.4 | 20.7 ± 2.7 | 37.4 ± 3.5 | 40.6 ± 4.5 | 26.6 ± 3.9 | 33.6 ± 3.7 |
| KML 2 ng | 48.7 ± 4.6 | 19.2 ± 2.6 | 33.7 ± 3.1 | 29.7 ± 3.4 | 20.1 ± 3.7 | 29.1 ± 4.1 |
| KML 20 ng | 50.6 ± 4.1 | 22.3 ± 2.9 | 34.6 ± 3.4 | 42.3 ± 4.0 | 20.5 ± 3.3 | 32.6 ± 3.5 |
| LOW STRESS | ||||||
| VEHICLE | 25.8 ± 3.1 ** | 13.1 ± 2.8 * | 15.2 ± 2.1 ** | 32.6 ± 3.8 | 12.6 ± 2.1 * | 24.6 ± 3.8 |
| KML 2 ng | 34.7 ± 3.7 | 14.1 ± 2.4 | 20.4 ± 2.8 ** | 33.7 ± 4.2 | 11.6 ± 2.5 | 21.7 ± 3.1 |
| KML 20 ng | 29.3 ± 3.1 * | 12.4 ± 2.8 * | 20.1 ± 3.0 ** | 33.3 ± 4.3 | 13.0 ± 2.5 | 18.3 ± 3.2 * |
| HIGH STRESS | ||||||
| VEHICLE | 36.7 ± 4.4 | 19.0 ± 2.4 | 16.8 ± 3.4 ** | 26.2 ± 4.3 | 15.4 ± 3.5 | 16.0 ± 3.3 ** |
| KML 2 ng | 34.9 ± 4.5 | 20.5 ± 2.8 | 18.4 ± 2.5 ** | 22.9 ± 3.0 | 13.7 ± 2.4 | 19.2 ± 3.3 |
| KML 20 ng | 28.1 ± 4.3 ** | 17.0 ± 2.9 | 14.2 ± 2.6 ** | 27.4 ± 5.2 | 16.2 ± 3.1 | 17.5 ± 3.8 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santori, A.; Morena, M.; Hill, M.N.; Campolongo, P. Hippocampal 2-Arachidonoyl Glycerol Signaling Regulates Time-of-Day- and Stress-Dependent Effects on Rat Short-Term Memory. Int. J. Mol. Sci. 2020, 21, 7316. https://doi.org/10.3390/ijms21197316
Santori A, Morena M, Hill MN, Campolongo P. Hippocampal 2-Arachidonoyl Glycerol Signaling Regulates Time-of-Day- and Stress-Dependent Effects on Rat Short-Term Memory. International Journal of Molecular Sciences. 2020; 21(19):7316. https://doi.org/10.3390/ijms21197316
Chicago/Turabian StyleSantori, Alessia, Maria Morena, Matthew N. Hill, and Patrizia Campolongo. 2020. "Hippocampal 2-Arachidonoyl Glycerol Signaling Regulates Time-of-Day- and Stress-Dependent Effects on Rat Short-Term Memory" International Journal of Molecular Sciences 21, no. 19: 7316. https://doi.org/10.3390/ijms21197316
APA StyleSantori, A., Morena, M., Hill, M. N., & Campolongo, P. (2020). Hippocampal 2-Arachidonoyl Glycerol Signaling Regulates Time-of-Day- and Stress-Dependent Effects on Rat Short-Term Memory. International Journal of Molecular Sciences, 21(19), 7316. https://doi.org/10.3390/ijms21197316