Licochalcone A Inhibits BDNF and TrkB Gene Expression and Hypoxic Growth of Human Tumor Cell Lines

 and
and
Abstract
:1. Introduction
2. Results
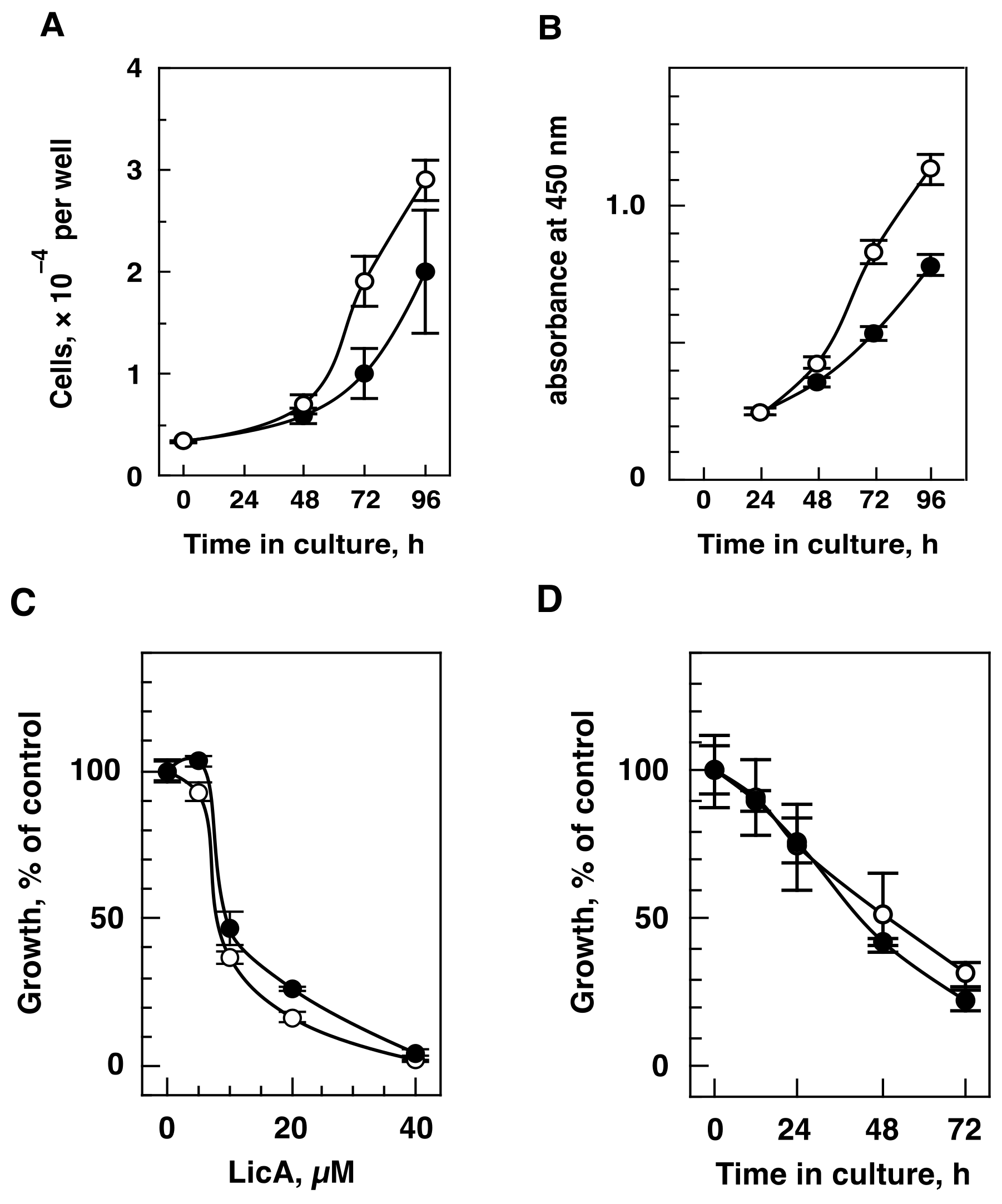
2.1. LicA Inhibits Cell Growth in Hypoxia
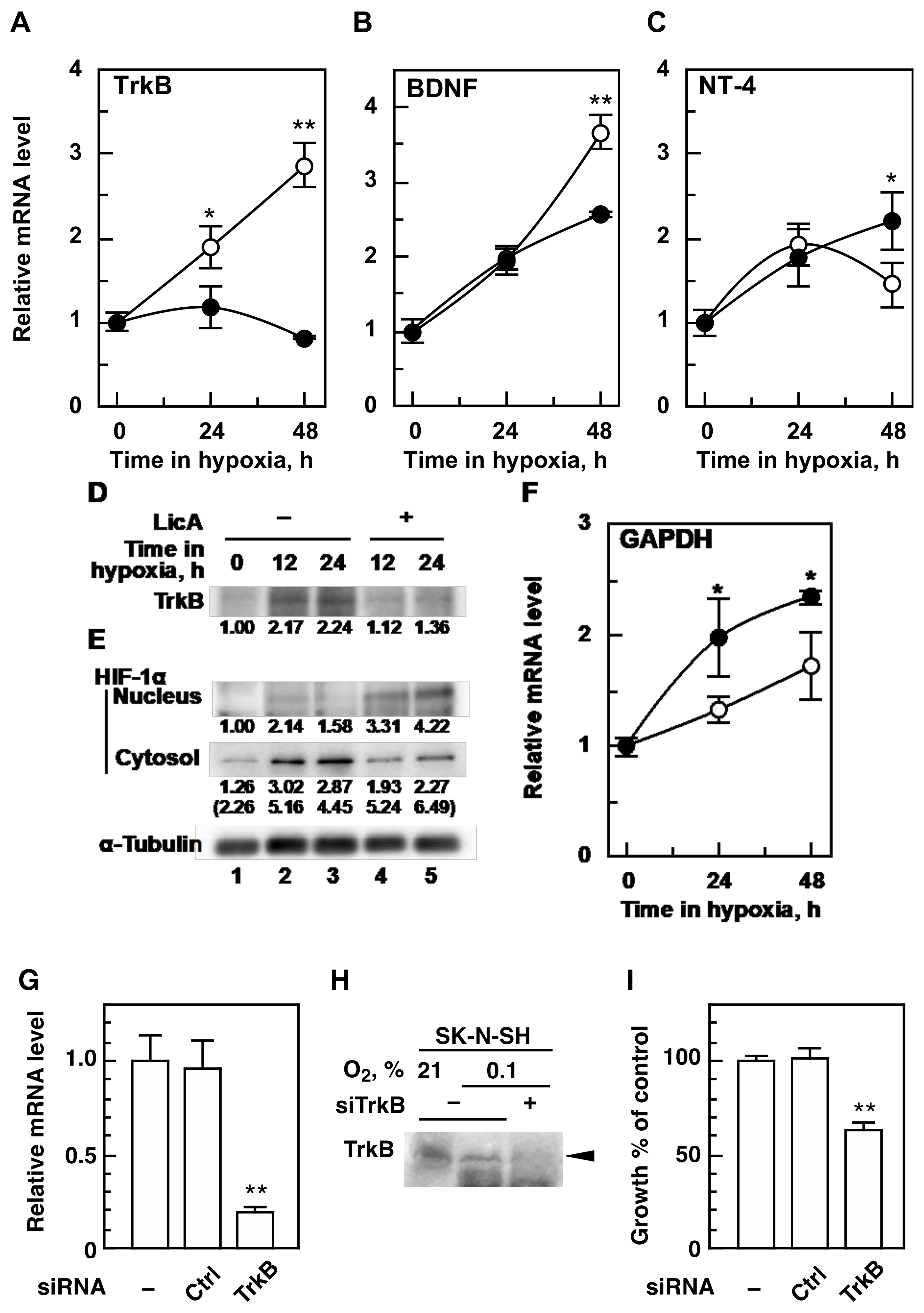
2.2. LicA Inhibits Hypoxic Activation of TrkB and BDNF Expression
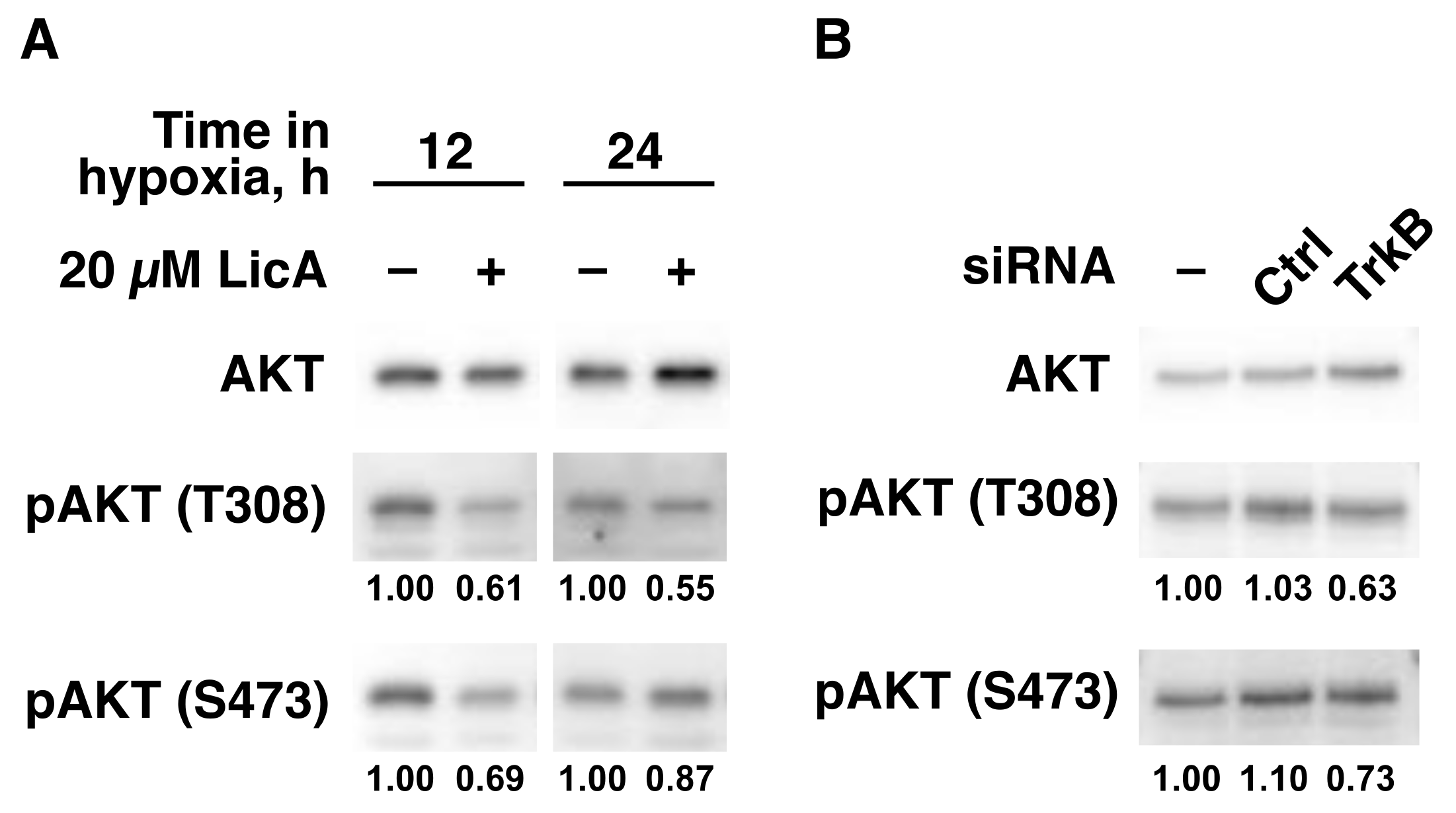
2.3. LicA Inhibits TrkB-Downstream Cell Growth Signaling
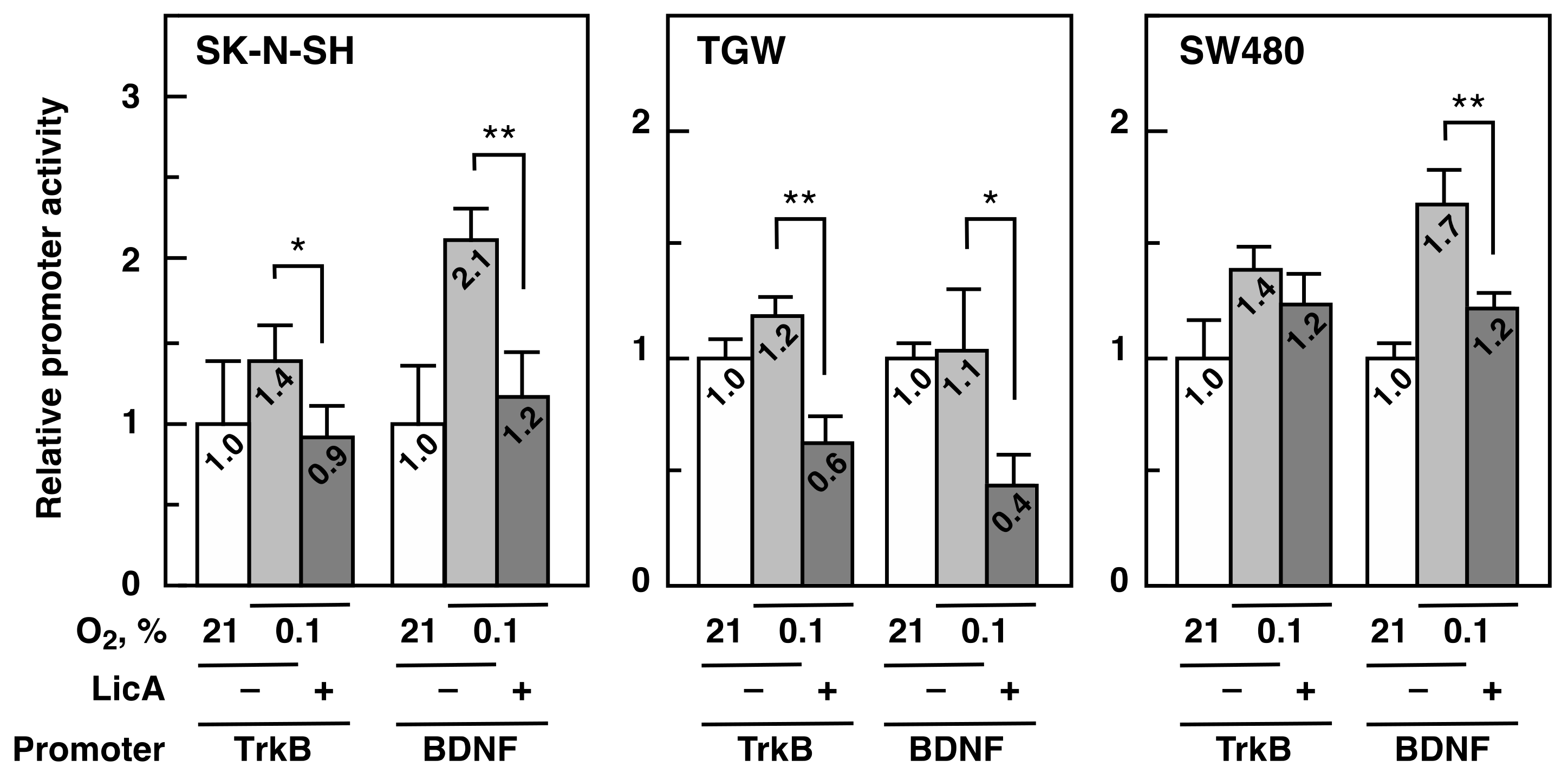
2.4. LicA inhibits TrkB and BDNF Promoter Activity
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cells and Cell Culture
4.3. Cell Growth Monitoring
4.4. Quantitative Reverse-Transcription Polymerase Chain Reaction
4.5. Immunoblotting
4.6. siRNAs and Transfection
4.7. Luciferase Reporter Gene Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AKT | Protein kinase B |
| BDNF | Brain-derived neurotrophic factor |
| CRC | Colorectal cancer |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| EMT | Epithelial-to-mesenchymal transition |
| ERK | Extracellular signal-regulated kinase |
| HIF-1 | Hypoxia-inducible factor-1 |
| HIF-1α | HIF-1 α-subunit |
| IC50 | 50% inhibitory concentration |
| JNK1 | c-Jun N-terminal kinase 1 |
| LicA | Licochalcone A |
| MAPK | Mitogen-activated protein kinase |
| MEK | MAPK/ERK kinase |
| mTOR | Mammalian target of rapamycin |
| NT-4 | Neurotrophin-4 |
| PI3K | Phosphatidylinositol 3-kinase |
| PKC | Protein kinase C |
| PLC- γ | Phospholipase C- γ |
| qRT-PCR | Quantitative reverse-transcription polymerase chain reaction |
| RTK | Receptor tyrosine kinase |
| siRNA | Small interfering RNA |
| TrkB | Tropomyosin-related kinase B |
| WST-8 | 2-(2-methoxy-4-nitrophenyl)-3-4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium |
References
- Ganapathy-Kanniappan, S.; Geschwind, J.-F.H. Tumor glycolysis as a target for cancer therapy: Progress and prospects. Mol. Cancer 2013, 12, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wigerup, C.; Påhlman, S.; Bexell, D. Therapeutic targeting of hypoxia and hypoxia-inducible factors in cancer. Pharmacol. Ther. 2016, 164, 152–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbet, C.; Feron, O. Tumour acidosis: From the passenger to the driver’s seat. Nat. Rev. Cancer 2017, 17, 577–593. [Google Scholar] [CrossRef] [PubMed]
- Redfern, A.D. The Kraken Wakes: Induced EMT as a driver of tumour aggression and poor outcome. Clin. Exp. Metastasis 2018, 35, 285–308. [Google Scholar] [CrossRef] [PubMed]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Chen, X.; Wang, L.; Chen, S. The sweet trap in tumors: Aerobic glycolysis and potential targets for therapy. Oncotarget 2016, 7, 38908–38926. [Google Scholar] [CrossRef] [Green Version]
- Akil, H.; Perraud, A.; Jauberteau, M.-O.; Mathonnet, M. Tropomyosin-related kinase B/brain derived-neurotrophic factor signaling pathway as a potential therapeutic target for colorectal cancer. World J. Gastroenterol. 2016, 22, 490–500. [Google Scholar] [CrossRef]
- Paoli, P.; Giannoni, E.; Chiarugi, P. Anoikis molecular pathways and its role in cancer progression. Biochim. Biophys. Acta 2013, 1833, 3481–3498. [Google Scholar] [CrossRef] [Green Version]
- Aoyama, M.; Asai, K.; Shishikura, T.; Kawamoto, T.; Miyachi, T.; Yokoi, T.; Togari, H.; Wada, Y.; Kato, T.; Nakagawara, A. Human neuroblastomas with unfavorable biologies express high levels of brain-derived neurotrophic factor mRNA and a variety of its variants. Cancer Lett. 2001, 164, 51–60. [Google Scholar] [CrossRef]
- Nakagawara, A.; Azar, C.G.; Scavarda, N.J.; Brodeur, G.M. Expression and function of TRK-B and BDNF in human neuroblastomas. Mol. Cell Biol. 1994, 14, 759–767. [Google Scholar] [CrossRef]
- Nakamura, K.; Martin, K.C.; Jackson, J.K.; Beppu, K.; Woo, C.-W.; Thiele, C.J. Brain-derived neurotrophic factor activation of TrkB induces vascular endothelial growth factor expression via hypoxia-inducible factor-1alpha in neuroblastoma cells. Cancer Res. 2006, 66, 4249–4255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radin, D.P.; Patel, P. BDNF: An Oncogene or Tumor Suppressor? Anticancer Res. 2017, 37, 3983–3990. [Google Scholar] [PubMed] [Green Version]
- Thiele, C.J.; Li, Z.; McKee, A.E. On Trk—The TrkB Signal Transduction Pathway Is an Increasingly Important Target in Cancer Biology. Clin. Cancer Res. 2009, 15, 5962–5967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douma, S.; Van Laar, T.; Zevenhoven, J.; Meuwissen, R.; Van Garderen, E.; Peeper, D.S. Suppression of anoikis and induction of metastasis by the neurotrophic receptor TrkB. Nature 2004, 430, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Smit, M.A.; Geiger, T.R.; Song, J.-Y.; Gitelman, I.; Peeper, D.S. A Twist-Snail axis critical for TrkB-induced epithelial-mesenchymal transition-like transformation, anoikis resistance, and metastasis. Mol. Cell Biol. 2009, 29, 3722–3737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinkevicius, K.W.; Kriegel, C.; Bellaria, K.J.; Lee, J.; Lau, A.N.; Leeman, K.T.; Zhou, P.; Beede, A.M.; Fillmore, C.M.; Caswell, D.; et al. Neurotrophin receptor TrkB promotes lung adenocarcinoma metastasis. Proc. Natl. Acad. Sci. USA 2014, 111, 10299–10304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martens, L.K.; Kirschner, K.M.; Warnecke, C.; Scholz, H. Hypoxia-inducible factor-1 (HIF-1) is a transcriptional activator of the TrkB neurotrophin receptor gene. J. Biol. Chem. 2007, 282, 14379–14388. [Google Scholar] [CrossRef] [Green Version]
- Tsai, J.-P.; Lee, C.-H.; Ying, T.-H.; Lin, C.-L.; Lin, C.-L.; Hsueh, J.-T.; Hsieh, Y.-H. Licochalcone A induces autophagy through PI3K/Akt/mTOR inactivation and autophagy suppression enhances Licochalcone A-induced apoptosis of human cervical cancer cells. Oncotarget 2015, 6, 28851–28866. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.-C.; Tsai, L.-L.; Tsai, J.-P.; Hsieh, S.-C.; Yang, S.-F.; Hsueh, J.-T.; Hsieh, Y.-H. Licochalcone A inhibits the migration and invasion of human lung cancer cells via inactivation of the Akt signaling pathway with downregulation of MMP-1/-3 expression. Tumour Biol. 2014, 35, 12139–12149. [Google Scholar] [CrossRef]
- Hao, W.; Yuan, X.; Yu, L.; Gao, C.; Sun, X.; Wang, D.; Zheng, Q. Licochalcone A-induced human gastric cancer BGC-823 cells apoptosis by regulating ROS-mediated MAPKs and PI3K/AKT signaling pathways. Sci. Rep. 2015, 5, 10336. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, X.; Wang, Y.; Sun, Q.; Chen, M.; Liu, S.; Zou, X. Licochalcone A suppresses hexokinase 2-mediated tumor glycolysis in gastric cancer via downregulation of the Akt signaling pathway. Oncol. Rep. 2018, 39, 1181–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, Z.; Gu, X.; Dong, Y.; Tan, F.; Liu, Z.; Thiele, C.J.; Li, Z. PI3K and MAPK pathways mediate the BDNF/TrkB-increased metastasis in neuroblastoma. Tumour Biol. 2016, 37, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Megison, M.L.; Gillory, L.A.; Beierle, E.A. Cell survival signaling in neuroblastoma. Anticancer Agents Med. Chem. 2013, 13, 563–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desmet, C.J.; Peeper, D.S. The neurotrophic receptor TrkB: A drug target in anti-cancer therapy? Cell Mol. Life Sci. 2006, 63, 755–759. [Google Scholar] [CrossRef]
- Roesler, R.; de Farias, C.B.; Abujamra, A.L.; Brunetto, A.L.; Schwartsmann, G. BDNF/TrkB signaling as an anti-tumor target. Expert Rev. Anticancer Ther. 2011, 11, 1473–1475. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Fornes, O.; Stigliani, A.; Gheorghe, M.; Castro-Mondragon, J.A.; van der Lee, R.; Bessy, A.; Chèneby, J.; Kulkarni, S.R.; Tan, G.; et al. JASPAR 2018: Update of the open-access database of transcription factor binding profiles and its web framework. Nucleic Acids Res. 2018, 46, D260–D266. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Liu, B.; Ji, R.; Jiang, X.; Yan, X.; Xin, Y. Targeting the BDNF/TrkB pathway for the treatment of tumors. Oncol. Lett. 2019, 17, 2031–2039. [Google Scholar] [CrossRef] [Green Version]
- Kato, H.; Okamura, K.; Kurosawa, Y.; Kishikawa, T.; Hashimoto, K. Characterization of DNA rearrangements of N-myc gene amplification in three neuroblastoma cell lines by pulsed-field gel electrophoresis. FEBS Lett. 1989, 250, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, W.C.; Weiss, W.A. Myc proteins as therapeutic targets. Oncogene 2010, 29, 1249–1259. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, D.; Eide, P.W.; Eilertsen, I.A.; Danielsen, S.A.; Eknæs, M.; Hektoen, M.; Lind, G.E.; Lothe, R.A. Epigenetic and genetic features of 24 colon cancer cell lines. Oncogenesis 2013, 2, e71. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, S.; Wang, X.; Yang, Z.; Ou, G. Overexpression of TrkB promotes the progression of colon cancer. APMIS 2010, 118, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Seeger, R.C.; Rayner, S.A.; Banerjee, A.; Chung, H.; Laug, W.E.; Neustein, H.B.; Benedict, W.F. Morphology, growth, chromosomal pattern and fibrinolytic activity of two new human neuroblastoma cell lines. Cancer Res. 1977, 37, 1364–1371. [Google Scholar] [PubMed]
- Iwasaki, I.; Sugiyama, H.; Kanazawa, S.; Hemmi, H. Establishment of cisplatin-resistant variants of human neuroblastoma cell lines, TGW and GOTO, and their drug cross-resistance profiles. Cancer Chemother. Pharmacol. 2002, 49, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Koike, J.; Kugoh, H.; Arita, M.; Ohhira, T.; Kikuchi, Y.; Funahashi, K.; Takamatsu, K.; Boland, C.R.; Koi, M.; et al. Down-regulation of MutS homolog 3 by hypoxia in human colorectal cancer. Biochim. Biophys. Acta 2012, 1823, 889–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, A.B.; Kapatos, G.; O’Farrell, C.; Gonzalez-de-Chavez, F.; Chiu, K.; Farrer, M.J.; Maidment, N.T. N-myc regulates parkin expression. J. Biol. Chem. 2004, 279, 28896–28902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalish, J.A.; Willis, D.J.; Li, C.; Link, J.J.; Deutsch, E.R.; Contreras, M.A.; Quist, W.C.; Logerfo, F.W. Temporal genomics of vein bypass grafting through oligonucleotide microarray analysis. J. Vasc. Surg. 2004, 39, 645–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiyama, H.; Arita, M.; Min, Z.; Zhong, X.; Iwasaki, I.; Hirano, K.; Shimatake, H.; Hemmi, H. A novel dysfunctional p53 mutation in the human neuroblastoma cell line TGW. Tohoku J. Exp. Med. 2003, 201, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Arita, M.; Zhong, X.; Min, Z.; Hemmi, H.; Shimatake, H. Multiple sites required for expression in 5’-flanking region of the hMLH1 gene. Gene 2003, 306, 57–65. [Google Scholar] [CrossRef]
- Laurière, M. A semidry electroblotting system efficiently transfers both high- and low-molecular-weight proteins separated by SDS-PAGE. Anal. Biochem. 1993, 212, 206–211. [Google Scholar] [CrossRef]
- Stoilov, P.; Castren, E.; Stamm, S. Analysis of the Human TrkB Gene Genomic Organization Reveals Novel TrkB Isoforms, Unusual Gene Length, and Splicing Mechanism. Biochem. Biophys. Res. Commun. 2002, 290, 1054–1065. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, M.; Fukuchi, M.; Tabuchi, A.; Kawahara, M.; Tsuneki, H.; Azuma, Y.; Chiba, Y.; Tsuda, M. Robust stimulation of TrkB induces delayed increases in BDNF and Arc mRNA expressions in cultured rat cortical neurons via distinct mechanisms. J. Neurochem. 2007, 103, 626–636. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Origin | Cell Proliferation in | IC50, µM | ||
|---|---|---|---|---|---|
| Normoxia | Hypoxia | ||||
| SK-N-SH | Neuroblastoma | 2.2 ± 0.07 | 1.9 ± 0.08 | 8.4 ± 0.3 | (4) |
| TGW | Neuroblastoma | 1.8 ± 0.07 | 1.4 ± 0.19 | 29.1 ± 1.1 | (4) |
| GOTO | Neuroblastoma | 2.0 ± 0.12 | 1.8 ± 0.09 | 31.1 ± 0.8 | (4) |
| SW480 | CRC | 2.3 ± 0.06 | 1.8 ± 0.06 | 8.8 ± 0.4 | (3) |
| SW620 | CRC | 3.6 ± 0.19 | 1.2 ± 0.06 | 7.0 ± 0.6 | (3) |
| HeLa S3 | Cervix carcinoma | 2.2 ± 0.08 | 1.5 ± 0.06 | 10.7 ± 1.2 | (3) |
| Cell Line | TrkB mRNA | BDNF mRNA | ||
|---|---|---|---|---|
| Untreated | LicA (Inhibition, %) | Untreated | LicA (Inhibition, %) | |
| SK-N-SH | 1.00 ± 0.091 | 0.29 ± 0.007 (71.4) ** | 1.00 ± 0.067 | 0.71 ± 0.011 (28.7) ** |
| TGW | 3.48 ± 0.098 | 1.68 ± 0.119 (51.7) ** | 3.81 ± 0.264 | 1.55 ± 0.284 (59.3) ** |
| GOTO | 16.63 ± 0.429 | 8.74 ± 0.894 (47.4) ** | N.D. | |
| SW480 | 2.49 ± 0.219 | 0.13 ± 0.013 (94.8) ** | 4.79 ± 0.406 | 0.16 ± 0.020 (96.6) ** |
| SW620 | 0.95 ± 0.026 | 0.26 ± 0.017 (72.6) ** | N.D. | |
| HeLa S3 | 0.09 ± 0.005 | 0.02 ± 0.001 (77.7) ** | 10.03 ± 0.250 | 8.66 ± 1.218 (13.7) |
| Target Gene | Sequence (5′ to 3′) | Source | |
|---|---|---|---|
| Forward | Reverse | ||
| TrkB | AAGGTGGCCCAGATGCTGTC | AATGTTATGTCGCTTGATGTGCTGA | HA169993 1 |
| BDNF | AGTTGGGAGCCTGAAATAGTGG | AGGATGCTGGTCCAAGTGGT | This study 2 |
| NT-4 | GCGGAGGAGGTGCTGACA | GGCCAGAAAAGGGGGCAA | This study 2 |
| GAPDH | GAAATCCCATCACCATCTTCCAGG | GAGCCCCAGCCTTCTCCATG | [35] |
| 18S rRNA | CGGCTACCACATCCAAGGAA | GCTGGAATTACCGCGGCT | [36] |
| Gene | Sequence (5′ to 3′) | |
|---|---|---|
| Forward | Reverse | |
| TrkB | F1: TCTAGACACTGTCCATGGAG | R1: ATGGGAAGGATCAAGAAACC |
| F2: ATCCCTGCCTTCATGGAGTG | R2: TGTACACCAGAAGAGTCTAA | |
| F3: CGGAGTTTTACGTGCGTCTG | R3: TTGCGTTCTGAGTGCTCCTAGCA | |
| F4: AACGAGACTCCAACCCATTG | ||
| F5: TCAGACAAGGCTTGCAAATG | ||
| BDNF | F1: GATTAACTGAGCCAGTTCTG | R1: ATTTTTTCACGTTCCCTTCG |
| F2: GCGAACTAGCATGAAATCTC | R2: GCAAACACACGTATAAGCTA | |
| Vector | R: CTTAATGTTTTTGGCATCTTCCA | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arita, M.; Koike, J.; Yoshikawa, N.; Kondo, M.; Hemmi, H. Licochalcone A Inhibits BDNF and TrkB Gene Expression and Hypoxic Growth of Human Tumor Cell Lines. Int. J. Mol. Sci. 2020, 21, 506. https://doi.org/10.3390/ijms21020506
Arita M, Koike J, Yoshikawa N, Kondo M, Hemmi H. Licochalcone A Inhibits BDNF and TrkB Gene Expression and Hypoxic Growth of Human Tumor Cell Lines. International Journal of Molecular Sciences. 2020; 21(2):506. https://doi.org/10.3390/ijms21020506
Chicago/Turabian StyleArita, Michitsune, Junichi Koike, Nobuji Yoshikawa, Motonari Kondo, and Hiromichi Hemmi. 2020. "Licochalcone A Inhibits BDNF and TrkB Gene Expression and Hypoxic Growth of Human Tumor Cell Lines" International Journal of Molecular Sciences 21, no. 2: 506. https://doi.org/10.3390/ijms21020506