Bone Marrow and Adipose Tissue Adenosine Receptors Effect on Osteogenesis and Adipogenesis


Abstract
1. Introduction
2. Adenosine Receptors Are Expressed on Bone Marrow-Derived Mesenchymal Stem Cells and Preadipocytes
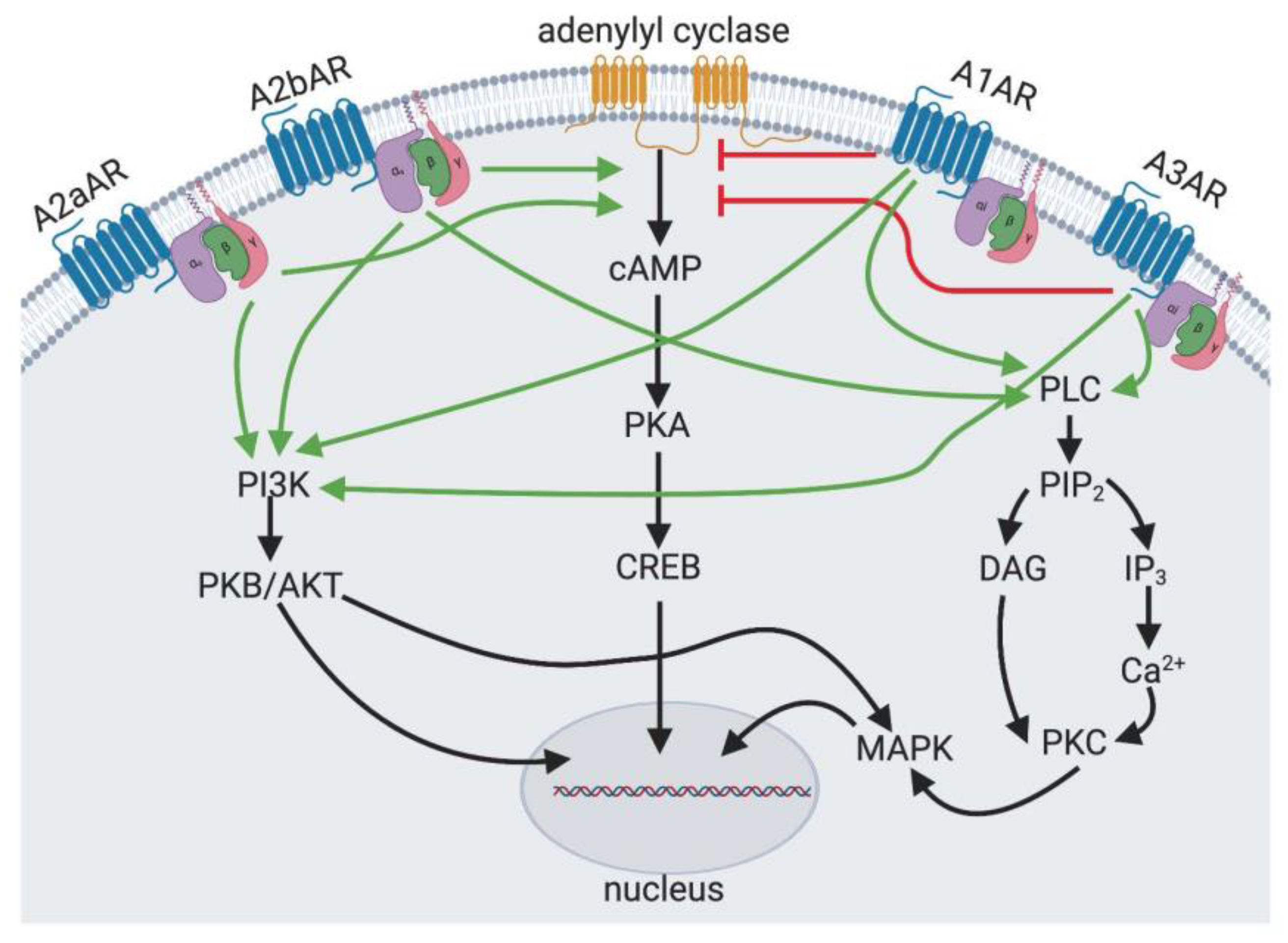
3. Regulation of Adipogenesis and Osteogenesis by Bone Marrow Cell Adenosine Receptors
4. Adenosine Receptors in Glucose Homeostasis and Obesity
5. Discussion: Implications for Therapeutic Benefit
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ATP | Adenosine triphosphate |
| ADP | Adenosine diphosphate |
| AMP | Adenosine monophosphate |
| A1AR | A1 adenosine receptor |
| A2aAR | A2a adenosine receptor |
| A2bAR | A2b adenosine receptor |
| A3AR | A3 adenosine receptor |
| cAMP | Cyclic adenosine monophosphate |
| DAG | Diacyglycerol |
| PKC | Protein kinase C |
| PLC | Phospholipase C |
| PI3K | Phosphoinositide 3-kinase |
| MAPK | Mitogen-activated protein kinase |
| MSC | Mesenchymal stem cell |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| CEBP | CCAAT enhancer binding protein |
| KLF4 | Krüppel-like factor 4 |
| Runx2 | Runt-related transcription factor 2 |
| TNFα | Tumor necrosis factor alpha |
| CPA | N6-Cyclopentyladenosine |
| PIA | Phenylisopropyladenosine |
| CCPA | 2-Chloro-N6-Cyclopentyladenosine |
| DPCPX | 8-Cyclopentyl-1,3-dipropylxanthine |
| CGS 21680 | 3-(4-(2-((6-amino-9-((2R,3R,4S,5S)-5-(ethylcarbomoyl)-3,4-dihydroxytetrahydrofuran-2-yl)-9H-purin-2-yl)amino)ethyl)phenyl)propanoic acid |
| SCH442416 | 2-(2-Furanyl)-7-[3-(4-methoxyphenyl)propyl]-7H-pyrazolo[4,3-e][1,2,4]triazolo[1,5-c]pyrimidin-5-amine |
| BAY 60-6583 | 2-[[6-Amino-3,5-dicyano-4-[4-(cyclopropylmethoxy)penyl]-2-pyridinyl]thio]-acetamide |
| MRS-1706 | N-(4-Acetylphenyl)-2-[4-(2,3,6,7-tetrahydro-2,6-dioxo-1,3-dipropyl-1H-purin-8-yl)phenoxy]acetamide |
| MRS-1754 | N-4(4-Cyanophenyl)-2-[4-(2,3,6,7-tetrahydro-2,6-dioxo-1,3-dipropyl-1H-purin-8-yl)phenoxy]acetamide |
| ATL-801 | (N-[5-(1-cyclopropyl-2,6-dioxo-3-propul-2,3,6,7-tetrahydro-1H-purin-8-yl)-pyridin-2-yl]-N-ethyl-nicotinamide |
| IB-MECA | N6-(3-Iodobenzyl)adenosine-5′-N-methyluronamide |
| C1-IB-MECA | 2-Chloro-N6-(3-Iodobenzyl)adenosine-5′-N-methyluronamide |
| MRS5698 | (1S,2R,3S,4R,5S)-4-[6-[[(3-Chlorophenyl)methyl]amino]-2-[2-(3,4-difluorophenyl)ethynyl]-9H-purin-9-yl]-2,3-dihydroxy-N-methylbicyclo[3.1.0]hexane-1-carboxamide |
| MRS1523 | 3-Propyl-6-ethyl-5-[(ethylthio)carbonyl]-2 phenyl-4-propyl-3-pyridine carboxylate |
| MRS1220 | N-[9-Chloro-2-(2-furanyl)[1,2,4]-triazol[1,5-c]quinazolin-5-yl]benzene acetamide |
| NECA | 5′-N-Ethylcarboxamidoadenosine |
| SDZWAG994 | N-Cyclohexyl-2′-O-methyladenosine |
| ARA | ([1S,2R,3R,5R]-3-methoxymethyl-5-[6-(1-[5-trifluoromethyl-pyridin-2-yl]pyrrolidine-3-[S]-ylamino)-purin-9-yl]cyclopentane-1,2-diol) |
| RPR749 | C22H26F3N7O3 |
| GS-9667 | (2S,3S,4R,5R)-2-(((2-fluorophenyl)thio)methyl)-5-(6-(((1R,2R)-2-hydroxycyclopentyl)amino)-9H-purin-9-yl)tetrahydrofuran-3,4-diol |
References
- Fredholm, B.B. Release of Adenosine-like Material from Isolated Perfused Dog Adipose Tissue Following Sympathetic Nerve Stimulation and its Inhibition by Adrenergic α-Receptor Blockade. Acta Physiol. Scand. 1976, 96, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, U.; Ebert, R.; Erbler, H.C. Adenosine release from isolated fat cells and its significance for the effects of hormones on cyclic 3′,5′-AMP levels and lipolysis. Naunyn Schmiedebergs Arch. Pharmacol. 1973, 276, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B. Adenosine—a physiological or pathophysiological agent? J. Mol. Med. 2013, 92, 201–206. [Google Scholar] [CrossRef]
- Borea, P.A.; Gessi, S.; Merighi, S.; Varani, K. Adenosine as a Multi-Signalling Guardian Angel in Human Diseases: When, Where and How Does it Exert its Protective Effects? Trends Pharmacol. Sci. 2016, 37, 419–434. [Google Scholar] [CrossRef] [PubMed]
- Peleli, M.; Fredholm, B.B.; Sobrevia, L.; Carlstrom, M. Pharmacological targeting of adenosine receptor signaling. Mol. Asp. Med. 2017, 55, 4–8. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, Y.; Nguyen, H.G.; Koupenova, M.; Chauhan, A.K.; Makitalo, M.; Jones, M.R.; Hilaire, C.S.; Seldin, D.C.; Toselli, P.; et al. The A2B adenosine receptor protects against inflammation and excessive vascular adhesion. J. Clin. Investig. 2006, 116, 1913–1923. [Google Scholar] [CrossRef]
- Dixon, A.K.; Gubitz, A.K.; Sirinathsinghji, D.J.; Richardson, P.J.; Freeman, T.C. Tissue distribution of adenosine receptor mRNAs in the rat. Br. J. Pharmacol. 1996, 118, 1461–1468. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Arslan, G.; Halldner, L.; Kull, B.; Schulte, G.; Wasserman, W. Structure and function of adenosine receptors and their genes. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2000, 362, 364–374. [Google Scholar] [CrossRef]
- Poth, J.M.; Brodsky, K.; Ehrentraut, H.; Grenz, A.; Eltzschig, H.K. Transcriptional control of adenosine signaling by hypoxia-inducible transcription factors during ischemic or inflammatory disease. J. Mol. Med. 2012, 91, 183–193. [Google Scholar] [CrossRef]
- Hilaire, C.S.; Carroll, S.H.; Chen, H.; Ravid, K. Mechanisms of induction of adenosine receptor genes and its functional significance. J. Cell. Physiol. 2009, 218, 35–44. [Google Scholar] [CrossRef]
- Karmazyn, M.; A Cook, M. Adenosine A1 receptor activation attenuates cardiac injury produced by hydrogen peroxide. Circ. Res. 1992, 71, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, N.R.; Kumar, G.K. Oxidative stress in the systemic and cellular responses to intermittent hypoxia. Biol. Chem. 2004, 385, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Madi, L.; Cohen, S.; Ochayin, A.; Bar-Yehuda, S.; Barer, F.; Fishman, P. Overexpression of A3 adenosine receptor in peripheral blood mononuclear cells in rheumatoid arthritis: Involvement of nuclear factor-kappaB in mediating receptor level. J. Rheumatol. 2007, 34, 20–26. [Google Scholar]
- Madi, L.; Ochaion, A.; Rath-Wolfson, L.; Bar-Yehuda, S.; Erlanger, A.; Ohana, G.; Harish, A.; Merimski, O.; Barer, F.; Fishman, P. The A3 Adenosine Receptor Is Highly Expressed in Tumor versus Normal Cells. Clin. Cancer Res. 2004, 10, 4472–4479. [Google Scholar] [CrossRef] [PubMed]
- Ochaion, A.; Bar-Yehuda, S.; Cohn, S.; Del Valle, L.; Perez-Liz, G.; Madi, L.; Barer, F.; Farbstein, M.; Fishman-Furman, S.; Reitblat, T.; et al. Methotrexate enhances the anti-inflammatory effect of CF101 via up-regulation of the A3 adenosine receptor expression. Arthritis Res. Ther. 2006, 8, R169. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Misra, R.; Aggarwal, A. Baseline adenosine receptor mRNA expression in blood as predictor of response to methotrexate therapy in patients with rheumatoid arthritis. Rheumatol. Int. 2019, 39, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Morello, S.; Ito, K.; Yamamura, S.; Lee, K.-Y.; Jazrawi, E.; DeSouza, P.; Barnes, P.; Cicala, C.; Adcock, I.M. IL-1 beta and TNF-alpha regulation of the adenosine receptor (A2A) expression: Differential requirement for NF-kappa B binding to the proximal promoter. J. Immunol. 2006, 177, 7173–7183. [Google Scholar] [CrossRef]
- Murphree, L.J.; Sullivan, G.W.; Marshall, M.A.; Linden, J. Lipopolysaccharide rapidly modifies adenosine receptor transcripts in murine and human macrophages: Role of NF-κB in A2Aadenosine receptor induction. Biochem. J. 2005, 391, 575–580. [Google Scholar] [CrossRef]
- Ahmad, A.; Ahmad, S.; Glover, L.E.; Miller, S.M.; Shannon, J.M.; Guo, X.; Franklin, W.A.; Bridges, J.P.; Schaack, J.B.; Colgan, S.P.; et al. Adenosine A2A receptor is a unique angiogenic target of HIF-2 in pulmonary endothelial cells. Proc. Natl. Acad. Sci. USA 2009, 106, 10684–10689. [Google Scholar] [CrossRef]
- Kobayashi, S.; Beitner-Johnson, D.; Conforti, L.; Millhorn, D.E. Chronic hypoxia reduces adenosine A2A receptor-mediated inhibition of calcium current in rat PC12 cells via downregulation of protein kinase A. J. Physiol. 1998, 512, 351–363. [Google Scholar] [CrossRef]
- Di Bonaventura, M.V.E.; Pucci, M.; Giusepponi, M.E.; Romano, A.; Lambertucci, C.; Volpini, R.; Di Bonaventura, E.M.; Gaetani, S.; Maccarrone, M.; D’Addario, C.; et al. Regulation of adenosine A2A receptor gene expression in a model of binge eating in the amygdaloid complex of female rats. J. Psychopharmacol. 2019, 33, 1550–1561. [Google Scholar] [CrossRef]
- Kong, T.; Westerman, K.A.; Faigle, M.; Eltzschig, H.K.; Colgan, S.P. HIF-dependent induction of adenosine A2B receptor in hypoxia. FASEB J. 2006, 20, 2242–2250. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Lu, H.; Samanta, D.; Salman, S.; Lu, Y.; Semenza, G.L. Hypoxia-inducible factor 1-dependent expression of adenosine receptor 2B promotes breast cancer stem cell enrichment. Proc. Natl. Acad. Sci. USA 2018, 115, E9640–E9648. [Google Scholar] [CrossRef] [PubMed]
- Sitkovsky, M.V.; Ohta, A. The ‘danger’ sensors that STOP the immune response: The A2 adenosine receptors? Trends Immunol. 2005, 26, 299–304. [Google Scholar] [CrossRef]
- Yang, M.; Ma, C.; Liu, S.; Shao, Q.; Gao, W.; Song, B.; Sun, J.; Xie, Q.; Zhang, Y.; Feng, A.; et al. HIF-dependent induction of adenosine receptor A2b skews human dendritic cells to a Th2-stimulating phenotype under hypoxia. Immunol. Cell Biol. 2009, 88, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Eckle, T.; Kewley, E.M.; Brodsky, K.S.; Tak, E.; Bonney, S.; Gobel, M.; Anderson, D.; Glover, L.E.; Riegel, A.K.; Colgan, S.P.; et al. Identification of Hypoxia-Inducible Factor HIF-1A as Transcriptional Regulator of the A2B Adenosine Receptor during Acute Lung Injury. J. Immunol. 2014, 192, 1249–1256. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.H.; Lee, J.; Kim, J.; Jo, Y.H.; Kirchner, V.A.; Kim, N.; Kwak, B.J.; Hwang, S.; Song, G.; Lee, S.; et al. HIF-1α regulates A2B adenosine receptor expression in liver cancer cells. Exp. Ther. Med. 2019, 18, 4231–4240. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.H.; Wigner, N.A.; Kulkarni, N.; Johnston-Cox, H.; Gerstenfeld, L.C.; Ravid, K. A2B adenosine receptor promotes mesenchymal stem cell differentiation to osteoblasts and bone formation in vivo. J. Biol. Chem. 2012, 287, 15718–15727. [Google Scholar] [CrossRef] [PubMed]
- Børglum, J.; Vassaux, G.; Richelsen, B.; Gaillard, D.; Darimont, C.; Ailhaud, G.; Négrel, R.; Richelsen, B. Changes in adenosine A1- and A2-receptor expression during adipose cell differentiation. Mol. Cell. Endocrinol. 1996, 117, 17–25. [Google Scholar] [CrossRef]
- Gharibi, B.; Abraham, A.A.; Ham, J.; Evans, B.A.J. Adenosine receptor subtype expression and activation influence the differentiation of mesenchymal stem cells to osteoblasts and adipocytes. J. Bone Miner. Res. 2011, 26, 2112–2124. [Google Scholar] [CrossRef]
- Gnad, T.; Scheibler, S.; Von Kügelgen, I.; Scheele, C.; Kilić, A.; Glöde, A.; Hoffmann, L.S.; Reverte-Salisa, L.; Horn, P.; Mutlu, S.; et al. Adenosine activates brown adipose tissue and recruits beige adipocytes via A2A receptors. Nat. Cell Biol. 2014, 516, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Johansson, S.M.; Salehi, A.; Sandström, M.E.; Westerblad, H.; Lundquist, I.; Carlsson, P.-O.; Fredholm, B.B.; Katz, A. A1 receptor deficiency causes increased insulin and glucagon secretion in mice. Biochem. Pharmacol. 2007, 74, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Schoelch, C.; Kuhlmann, J.; Gossel, M.; Mueller, G.; Neumann-Haefelin, C.; Belz, U.; Kalisch, J.; Biemer-Daub, G.; Kramer, W.; Juretschke, H.-P.; et al. Characterization of adenosine-A1 receptor-mediated antilipolysis in rats by tissue microdialysis, 1H-spectroscopy, and glucose clamp studies. Diabetes 2004, 53, 1920–1926. [Google Scholar] [CrossRef] [PubMed]
- Eisenstein, A.; Carroll, S.H.; Johnston-Cox, H.; Farb, M.; Gokce, N.; Ravid, K. An Adenosine Receptor-Krüppel-like Factor 4 Protein Axis Inhibits Adipogenesis. J. Biol. Chem. 2014, 289, 21071–21081. [Google Scholar] [CrossRef] [PubMed]
- Ravid, K.; Lowenstein, J.M. Changes in adenosine receptors during differentiation of 3T3-F442A cells to adipocytes. Biochem. J. 1988, 249, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Vassaux, G.; Gaillard, D.; Mari, B.; Ailhaud, G.; Négrel, R. Differential Expression of Adenosine A1 and Adenosine A2 Receptors in Preadipocytes and Adipocytes. Biochem. Biophys. Res. Commun. 1993, 193, 1123–1130. [Google Scholar] [CrossRef]
- Johnston-Cox, H.; Koupenova, M.; Yang, D.; Corkey, B.; Gokce, N.; Farb, M.G.; Lebrasseur, N.K.; Ravid, K. The A2b Adenosine Receptor Modulates Glucose Homeostasis and Obesity. PLoS ONE 2012, 7, e40584. [Google Scholar] [CrossRef]
- Koupenova, M.; Johnston-Cox, H.; Vezeridis, A.; Gavras, H.; Yang, D.; Zannis, V.I.; Ravid, K. A2b adenosine receptor regulates hyperlipidemia and atherosclerosis. Circulation 2011, 125, 354–363. [Google Scholar] [CrossRef]
- Kaartinen, J.M.; Hreniuk, S.P.; Martin, L.F.; Ranta, S.; LaNoue, K.F.; Ohisalo, J.J. Attenuated adenosine-sensitivity and decreased adenosine-receptor number in adipocyte plasma membranes in human obesity. Biochem. J. 1991, 279, 17–22. [Google Scholar] [CrossRef]
- Hajjawi, M.O.R.; Patel, J.J.; Corcelli, M.; Arnett, T.; Orriss, I.R. Lack of effect of adenosine on the function of rodent osteoblasts and osteoclasts in vitro. Purinergic Signal. 2016, 12, 247–258. [Google Scholar] [CrossRef]
- Barroso, I.; Gurnell, M.; Crowley, V.E.F.; Agostini, M.; Schwabe, J.W.; Soos, M.A.; Maslen, G.L.; Williams, T.D.M.; Lewis, H.; Schafer, A.J. Dominant negative mutations in human PPARgamma associated with severe insulin resistance, diabetes mellitus and hypertension. Nature 1999, 402, 880–883. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Mullican, S.E.; DiSpirito, J.R.; Peed, L.C.; Lazar, M.A. Lipoatrophy and severe metabolic disturbance in mice with fat-specific deletion of PPARgamma. Proc. Natl. Acad. Sci. USA 2013, 110, 18656–18661. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; Hu, E.; Spiegelman, B.M. Stimulation of adipogenesis in fibroblasts by PPAR gamma 2, a lipid-activated transcription factor. Cell 1994, 79, 1147–1156. [Google Scholar] [CrossRef]
- Cao, Z.; Umek, R.M.; McKnight, S.L. Regulated expression of three C/EBP isoforms during adipose conversion of 3T3-L1 cells. Genes Dev. 1991, 5, 1538–1552. [Google Scholar] [CrossRef] [PubMed]
- Steger, D.J.; Grant, G.R.; Schupp, M.; Tomaru, T.; Lefterova, M.I.; Schug, J.; Manduchi, E.; Stoeckert, C.J.; Lazar, M.A. Propagation of adipogenic signals through an epigenomic transition state. Genes Dev. 2010, 24, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Yoshida, N.; Kishimoto, T.; Akira, S. Defective adipocyte differentiation in mice lacking the C/EBPbeta and/or C/EBPdelta gene. EMBO J. 1997, 16, 7432–7443. [Google Scholar] [CrossRef]
- Wu, Z.; Bucher, N.L.; Farmer, S.R. Induction of peroxisome proliferator-activated receptor gamma during the conversion of 3T3 fibroblasts into adipocytes is mediated by C/EBPbeta, C/EBPdelta, and glucocorticoids. Mol. Cell. Biol. 1996, 16, 4128–4136. [Google Scholar] [CrossRef]
- Gharibi, B.; Abraham, A.A.; Ham, J.; Evans, B.A.J. Contrasting effects of A1 and A2b adenosine receptors on adipogenesis. Int. J. Obes. 2011, 36, 397–406. [Google Scholar] [CrossRef]
- Hinz, S.; Lacher, S.K.; Seibt, B.F.; Müller, C.E. BAY60-6583 Acts as a Partial Agonist at Adenosine A2B Receptors. J. Pharmacol. Exp. Ther. 2014, 349, 427–436. [Google Scholar] [CrossRef]
- Katebi, M.; Soleimani, M.; Cronstein, B.N. Adenosine A2Areceptors play an active role in mouse bone marrow-derived mesenchymal stem cell development. J. Leukoc. Biol. 2008, 85, 438–444. [Google Scholar] [CrossRef]
- Corciulo, C.; Wilder, T.; Cronstein, B.N. Adenosine A2B receptors play an important role in bone homeostasis. Purinergic Signal. 2016, 12, 537–547. [Google Scholar] [CrossRef]
- Shih, Y.-R.V.; Liu, M.; Kwon, S.K.; Iida, M.; Gong, Y.; Sangaj, N.; Varghese, S. Dysregulation of ectonucleotidase-mediated extracellular adenosine during postmenopausal bone loss. Sci. Adv. 2019, 5, eaax1387. [Google Scholar] [CrossRef] [PubMed]
- Kara, F.M.; Chitu, V.; Sloane, J.; Axelrod, M.; Fredholm, B.B.; Stanley, E.R.; Cronstein, B.N. Adenosine A 1 receptors (A 1 Rs) play a critical role in osteoclast formation and function. FASEB J. 2010, 24, 2325–2333. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Cronstein, B.N. Adenosine A1 receptor regulates osteoclast formation by altering TRAF6/TAK1 signaling. Purinergic Signal 2012, 8, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Kara, F.M.; Doty, S.B.; Boskey, A.L.; Goldring, S.; Zaidi, M.; Fredholm, B.B.; Cronstein, B.N. Adenosine A1receptors regulate bone resorption in mice: Adenosine A1receptor blockade or deletion increases bone density and prevents ovariectomy-induced bone loss in adenosine A1receptor-knockout mice. Arthritis Rheum. 2010, 62, 534–541. [Google Scholar] [CrossRef]
- Mediero, A.; Kara, F.M.; Wilder, T.; Cronstein, B.N. Adenosine A2A Receptor Ligation Inhibits Osteoclast Formation. Am. J. Pathol. 2012, 180, 775–786. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Mazumder, A.; Wilder, T.; Cronstein, B.N. Adenosine regulates bone metabolism via A 1, A 2A, and A 2B receptors in bone marrow cells from normal humans and patients with multiple myeloma. FASEB J. 2013, 27, 3446–3454. [Google Scholar] [CrossRef]
- Takedachi, M.; Oohara, H.; Smith, B.J.; Iyama, M.; Kobashi, M.; Maeda, K.; Long, C.L.; Humphrey, M.B.; Stoecker, B.J.; Toyosawa, S.; et al. CD73-generated adenosine promotes osteoblast differentiation. J. Cell. Physiol. 2012, 227, 2622–2631. [Google Scholar] [CrossRef]
- Bradaschia-Correa, V.; Josephson, A.M.; Egol, A.J.; Mizrahi, M.M.; Leclerc, K.; Huo, J.; Cronstein, B.N.; Leucht, P. Ecto-5′-nucleotidase (CD73) regulates bone formation and remodeling during intramembranous bone repair in aging mice. Tissue Cell 2017, 49, 545–551. [Google Scholar] [CrossRef]
- Wu, W.; Xiao, Z.; Chen, Y.; Deng, Y.; Zeng, D.; Liu, Y.; Huang, F.; Wang, J.; Liu, Y.; Bellanti, J.A. CD39 Produced from Human GMSCs Regulates the Balance of Osteoclasts and Osteoblasts through the Wnt/beta-Catenin Pathway in Osteoporosis. Mol. Ther. 2020, 28, 1518–1532. [Google Scholar] [CrossRef]
- Gardinier, J.D.; Rostami, N.; Juliano, L.; Zhang, C. Bone adaptation in response to treadmill exercise in young and adult mice. Bone Rep. 2018, 8, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.; Rajachar, R.M.; Allen, M.R.; Bloomfield, S.A.; Robey, P.G.; Young, M.F.; Kohn, D.H. Exercise-induced changes in the cortical bone of growing mice are bone- and gender-specific. Bone 2007, 40, 1120–1127. [Google Scholar] [CrossRef] [PubMed]
- Zarębska, E.; Kusy, K.; Słomińska, E.M.; Kruszyna, Ł.; Zieliński, J. Alterations in Exercise-Induced Plasma Adenosine Triphosphate Concentration in Highly Trained Athletes in a One-Year Training Cycle. Metabolites 2019, 9, 230. [Google Scholar] [CrossRef] [PubMed]
- Honnor, R.C.; Dhillon, G.S.; Londos, C. cAMP-dependent protein kinase and lipolysis in rat adipocytes. II. Definition of steady-state relationship with lipolytic and antilipolytic modulators. J. Biol. Chem. 1985, 260, 15130–15138. [Google Scholar]
- Fain, J.N.; Wieser, P.B. Effects of adenosine deaminase on cyclic adenosine monophosphate accumulation, lipolysis, and glucose metabolism of fat cells. J. Biol. Chem. 1975, 250, 1027–1034. [Google Scholar]
- Szkudelski, T.; Szkudelska, K.; Nogowski, L. Effects of adenosine A1 receptor antagonism on lipogenesis and lipolysis in isolated rat adipocytes. Physiol. Res. 2008, 58, 863–871. [Google Scholar]
- Vannucci, S.J.; Klim, C.M.; Martin, L.F.; LaNoue, K.F. A1-adenosine receptor-mediated inhibition of adipocyte adenylate cyclase and lipolysis in Zucker rats. Am. J. Physiol. Metab. 1989, 257, E871–E878. [Google Scholar] [CrossRef]
- Hoffman, B.B.; Dall’Aglio, E.; Hollenbeck, C.; Chang, H.; Reaven, G.M. Suppression of free fatty acids and triglycerides in normal and hypertriglyceridemic rats by the adenosine receptor agonist phenylisopropyladenosine. J. Pharmacol. Exp. Ther. 1986, 239, 715–718. [Google Scholar]
- Johansson, S.M.; Lindgren, E.; Yang, J.-N.; Herling, A.W.; Fredholm, B.B. Adenosine A1 receptors regulate lipolysis and lipogenesis in mouse adipose tissue—Interactions with insulin. Eur. J. Pharmacol. 2008, 597, 92–101. [Google Scholar] [CrossRef]
- Yang, T.; Gao, X.; Sandberg, M.; Zollbrecht, C.; Zhang, X.-M.; Hezel, M.; Liu, M.; Peleli, M.; Lai, E.-Y.; A Harris, R.; et al. Abrogation of adenosine A1 receptor signalling improves metabolic regulation in mice by modulating oxidative stress and inflammatory responses. Diabetologia 2015, 58, 1610–1620. [Google Scholar] [CrossRef]
- Faulhaber-Walter, R.; Jou, W.; Mizel, D.; Li, L.; Zhang, J.; Kim, S.M.; Huang, Y.; Chen, M.; Briggs, J.P.; Gavrilova, O.; et al. Impaired Glucose Tolerance in the Absence of Adenosine A1 Receptor Signaling. Diabetes 2011, 60, 2578–2587. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Ginsberg, H.N.; Erlanger, B.F. Overexpression of the A1 adenosine receptor in adipose tissue protects mice from obesity-related insulin resistance. Diabetes Obes. Metab. 2001, 3, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Rice, A.M.; Fain, J.N.; Rivkees, S.A. A1 adenosine receptor activation increases adipocyte leptin secretion. Endocrinology 2000, 141, 1442–1445. [Google Scholar] [CrossRef]
- Neméth, Z.H.; Bleich, D.; Csóka, B.; Pacher, P.; Mabley, J.G.; Himer, L.; Vizi, E.S.; Deitch, E.A.; Szabó, C.; Cronstein, B.N.; et al. Adenosine receptor activation ameliorates type 1 diabetes. FASEB J. 2007, 21, 2379–2388. [Google Scholar] [CrossRef] [PubMed]
- Figler, R.A.; Wang, G.; Srinivasan, S.; Jung, D.Y.; Zhang, Z.; Pankow, J.; Ravid, K.; Fredholm, B.; Hedrick, C.C.; Rich, S.S.; et al. Links Between Insulin Resistance, Adenosine A2B Receptors, and Inflammatory Markers in Mice and Humans. Diabetes 2011, 60, 669–679. [Google Scholar] [CrossRef]
- Johnston-Cox, H.; Eisenstein, A.S.; Koupenova, M.; Carroll, S.; Ravid, K. The Macrophage A2b Adenosine Receptor Regulates Tissue Insulin Sensitivity. PLoS ONE 2014, 9, e98775. [Google Scholar] [CrossRef]
- Csóka, B.; Koscsó, B.; Törő, G.; Kókai, E.; Virág, L.; Németh, Z.H.; Pacher, P.; Bai, P.; Haskó, G. A2B Adenosine Receptors Prevent Insulin Resistance by Inhibiting Adipose Tissue Inflammation via Maintaining Alternative Macrophage Activation. Diabetes 2014, 63, 850–866. [Google Scholar] [CrossRef]
- Merighi, S.; Borea, P.A.; Gessi, S. Adenosine receptors and diabetes: Focus on the A2B adenosine receptor subtype. Pharmacol. Res. 2015, 99, 229–236. [Google Scholar] [CrossRef]
- Pei, Y.; Li, H.; Cai, Y.; Zhou, J.; Luo, X.; Ma, L.; McDaniel, K.; Zeng, T.; Chen, Y.; Qian, X.; et al. Regulation of adipose tissue inflammation by adenosine 2A receptor in obese mice. J. Endocrinol. 2018, 239, 365–376. [Google Scholar] [CrossRef]
- Cai, Y.; Li, H.; Liu, M.; Pei, Y.; Zheng, J.; Zhou, J.; Luo, X.; Huang, W.; Ma, L.; Yang, Q.; et al. Disruption of adenosine 2A receptor exacerbates NAFLD through increasing inflammatory responses and SREBP1c activity. Hepatology 2018, 68, 48–61. [Google Scholar] [CrossRef]
- Csóka, B.; Törő, G.; Vindeirinho, J.; Varga, Z.V.; Koscsó, B.; Németh, Z.H.; Kókai, E.; Antonioli, L.; Suleiman, M.; Marchetti, P.; et al. A 2A adenosine receptors control pancreatic dysfunction in high-fat-diet-induced obesity. FASEB J. 2017, 31, 4985–4997. [Google Scholar] [CrossRef] [PubMed]
- DeOliveira, C.C.; Caria, C.; Gotardo, E.M.F.; Ribeiro, M.L.; Gambero, A. Role of A 1 and A 2A adenosine receptor agonists in adipose tissue inflammation induced by obesity in mice. Eur. J. Pharmacol. 2017, 799, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Ruan, C.; Gao, P. GW29-e1353 A2A Receptor Activation Attenuates Hypertensive Cardiac Remodeling via Promoting Brown Adipose Tissue-Derived FGF21. J. Am. Coll. Cardiol. 2018, 72, C41. [Google Scholar] [CrossRef]
- Lahesmaa, M.; Oikonen, V.; Helin, S.; Luoto, P.; Din, M.U.; Pfeifer, A.; Nuutila, P.; Virtanen, K.A. Regulation of human brown adipose tissue by adenosine and A2A receptors—Studies with [15O]H2O and [11C]TMSX PET/CT. Eur. J. Nucl. Med. Mol. Imaging 2018, 46, 743–750. [Google Scholar] [CrossRef]
- Xiao, C.; Liu, N.; Jacobson, K.A.; Gavrilova, O.; Reitman, M.L. Physiology and effects of nucleosides in mice lacking all four adenosine receptors. PLoS Biol. 2019, 17, e3000161. [Google Scholar] [CrossRef]
- Gnad, T.; Navarro, G.; Lahesmaa, M.; Reverte-Salisa, L.; Copperi, F.; Cordomi, A.; Naumann, J.; Hochhäuser, A.; Haufs-Brusberg, S.; Wenzel, D.; et al. Adenosine/A2B Receptor Signaling Ameliorates the Effects of Aging and Counteracts Obesity. Cell Metab. 2020, 32, 56–70.e7. [Google Scholar] [CrossRef]
- Jacobson, K.A.; Tosh, D.K.; Jain, S.; Gao, Z.-G. Historical and Current Adenosine Receptor Agonists in Preclinical and Clinical Development. Front. Cell. Neurosci. 2019, 13, 124. [Google Scholar] [CrossRef]
- Ishikawa, J.; Mitani, H.; Bandoh, T.; Kimura, M.; Totsuka, T.; Hayashi, S. Hypoglycemic and hypotensive effects of 6-cyclohexyl-2′-O-methyl-adenosine, an adenosine A1 receptor agonist, in spontaneously hypertensive rat complicated with hyperglycemia. Diabetes Res. Clin. Pract. 1998, 39, 3–9. [Google Scholar] [CrossRef]
- Zannikos, P.N.; Rohatagi, S.; Jensen, B.K. Pharmacokinetic-pharmacodynamic modeling of the antilipolytic effects of an adenosine receptor agonist in healthy volunteers. J. Clin. Pharmacol. 2001, 41, 61–69. [Google Scholar] [CrossRef]
- Shah, B.; Rohatagi, S.; Natarajan, C.; Kirkesseli, S.; Baybutt, R.; Jensen, B.K. Pharmacokinetics, Pharmacodynamics, and Safety of a Lipid-Lowering Adenosine A1 Agonist, RPR749, in Healthy Subjects. Am. J. Ther. 2004, 11, 175–189. [Google Scholar] [CrossRef]
- Dhalla, A.K.; Santikul, M.; Smith, M.; Wong, M.-Y.; Shryock, J.C.; Belardinelli, L. Antilipolytic Activity of a Novel Partial A1 Adenosine Receptor Agonist Devoid of Cardiovascular Effects: Comparison with Nicotinic Acid. J. Pharmacol. Exp. Ther. 2007, 321, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Dhalla, A.K.; Wong, M.Y.; Voshol, P.J.; Belardinelli, L.; Reaven, G.M. A1 adenosine receptor partial agonist lowers plasma FFA and improves insulin resistance induced by high-fat diet in rodents. Am. J. Physiol. Metab. 2007, 292, E1358–E1363. [Google Scholar] [CrossRef] [PubMed]
- Staehr, P.M.; Dhalla, A.K.; Zack, J.; Wang, X.; Ho, Y.L.; Bingham, J.; Belardinelli, L. Reduction of Free Fatty Acids, Safety, and Pharmacokinetics of Oral GS-9667, an A1Adenosine Receptor Partial Agonist. J. Clin. Pharmacol. 2013, 53, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-F.; Cunha, R.A. The belated US FDA approval of the adenosine A2A receptor antagonist istradefylline for treatment of Parkinson’s disease. Purinergic Signal. 2020, 16, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Charvin, D.; Medori, R.; Hauser, R.A.; Rascol, O. Therapeutic strategies for Parkinson disease: Beyond dopaminergic drugs. Nat. Rev. Drug Discov. 2018, 17, 804–822. [Google Scholar] [CrossRef]


{kind=link}
| Receptor | Agonist (+)/Antagonist (−) | Drug Acronym |
|---|---|---|
| A1AR | + | CPA, PIA, CCPA |
| A1AR | − | DPCPX |
| A2aAR | + | CGS 21680 |
| A2aAR | − | SCH442416 |
| A2bAR | + | BAY 60-6583 |
| A2bAR | − | MRS1706, MRS-1754, ATL-801 |
| A3AR | + | IB-MECA, C1-IB-MECA, MRS5698 |
| A3AR | − | MRS1523, MRS1220 |
| All ARs | + | NECA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eisenstein, A.; Chitalia, S.V.; Ravid, K. Bone Marrow and Adipose Tissue Adenosine Receptors Effect on Osteogenesis and Adipogenesis. Int. J. Mol. Sci. 2020, 21, 7470. https://doi.org/10.3390/ijms21207470
Eisenstein A, Chitalia SV, Ravid K. Bone Marrow and Adipose Tissue Adenosine Receptors Effect on Osteogenesis and Adipogenesis. International Journal of Molecular Sciences. 2020; 21(20):7470. https://doi.org/10.3390/ijms21207470
Chicago/Turabian StyleEisenstein, Anna, Shlok V. Chitalia, and Katya Ravid. 2020. "Bone Marrow and Adipose Tissue Adenosine Receptors Effect on Osteogenesis and Adipogenesis" International Journal of Molecular Sciences 21, no. 20: 7470. https://doi.org/10.3390/ijms21207470
APA StyleEisenstein, A., Chitalia, S. V., & Ravid, K. (2020). Bone Marrow and Adipose Tissue Adenosine Receptors Effect on Osteogenesis and Adipogenesis. International Journal of Molecular Sciences, 21(20), 7470. https://doi.org/10.3390/ijms21207470