Role of CD4+ T Cells in Allergic Airway Diseases: Learning from Murine Models


Abstract
1. Introduction
2. Role of CD4+ T Cells in Allergic Inflammation
3. IgE/Mast Cell- and CD4+ T Cell-Dependent Cascades in Allergic Inflammation
4. T Cell Subsets and Allergic Airway Diseases
5. Antigen-Specific T Cell Receptor Tg Mice
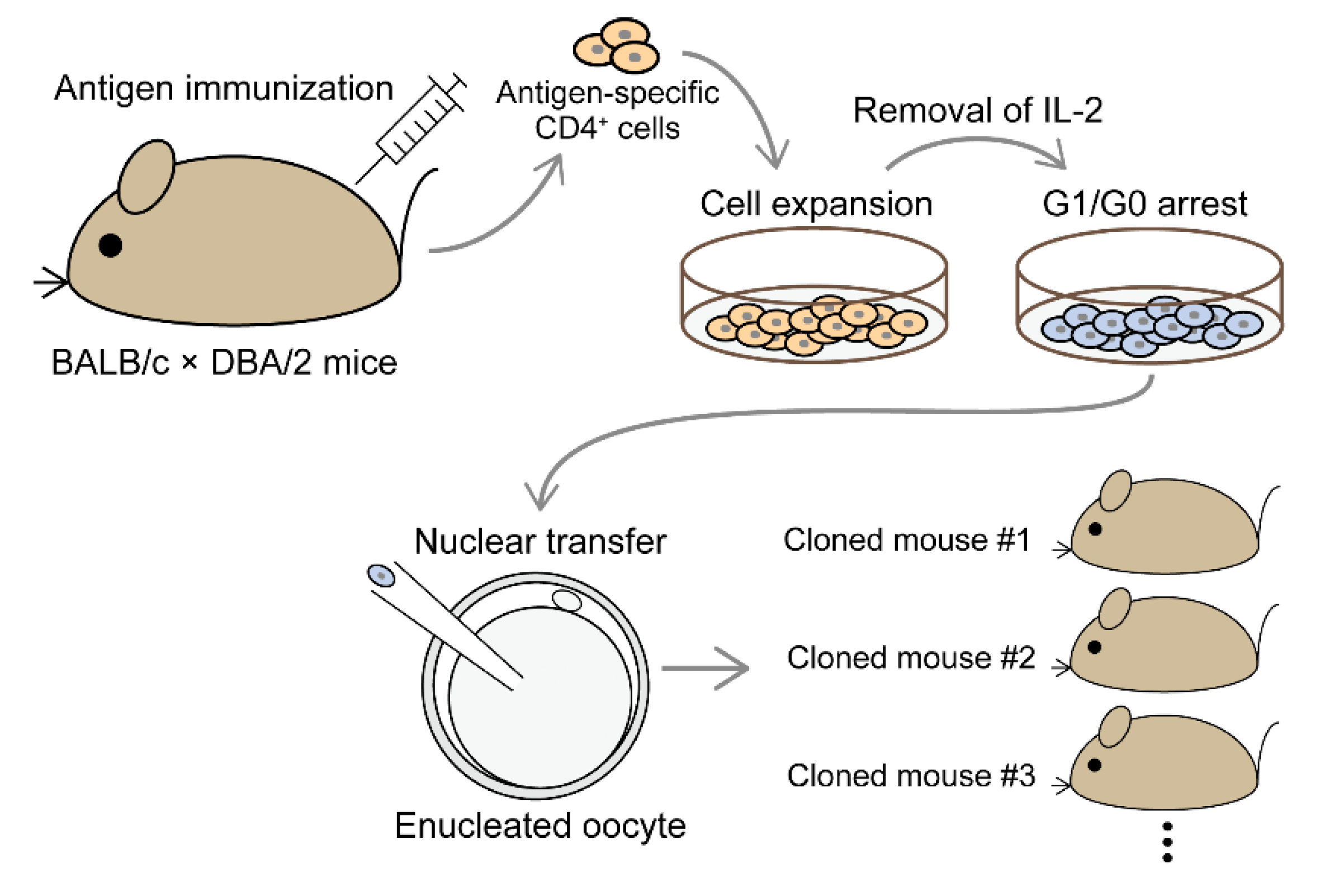
6. Generation of Cloned Mice Derived from Antigen-Specific CD4+ T Cells
7. Antigen Reactivity of CD4+ T Cells in Cloned Mice
8. Usefulness of Antigen-Specific T Cell-Cloned Mice for Investigating Allergic Airway Inflammation In Vivo
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tada, T.; Takemori, T.; Okumura, K.; Nonaka, M.; Tokuhisa, T. Two distinct types of helper T cells involved in the secondary antibody response: Independent and synergistic effects of Ia- and Ia+ helper T cells. J. Exp. Med. 1978, 147, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T.R.; Cherwinski, H.; Bond, M.W.; Giedlin, M.A.; Coffman, R.L. Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J. Immunol. 1986, 136, 2348–2357. [Google Scholar] [PubMed]
- Hamid, Q.; Azzawi, M.; Ying, S.; Moqbel, R.; Wardlaw, A.J.; Corrigan, C.J.; Bradley, B.; Durham, S.R.; Collins, J.V.; Jeffery, P.K.; et al. Expression of mRNA for interleukin-5 in mucosal bronchial biopsies from asthma. J. Clin. Investig. 1991, 87, 1541–1546. [Google Scholar] [CrossRef]
- Kay, A.B.; Ying, S.; Varney, V.; Gaga, M.; Durham, S.R.; Moqbel, R.; Wardlaw, A.J.; Hamid, Q. Messenger RNA expression of the cytokine gene cluster, interleukin 3 (IL-3), IL-4, IL-5, and granulocyte/macrophage colony-stimulating factor, in allergen-induced late-phase cutaneous reactions in atopic subjects. J. Exp. Med. 1991, 173, 775–778. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.; Hamid, Q.; Bentley, A.; Ying, S.; Kay, A.B.; Durham, S.R. Activation of CD4+ T cells, increased TH2-type cytokine mRNA expression, and eosinophil recruitment in bronchoalveolar lavage after allergen inhalation challenge in patients with atopic asthma. J. Allergy Clin. Immunol. 1993, 92, 313–324. [Google Scholar] [CrossRef]
- Krishnaswamy, G.; Liu, M.C.; Su, S.N.; Kumai, M.; Xiao, H.Q.; Marsh, D.G.; Huang, S.K. Analysis of cytokine transcripts in the bronchoalveolar lavage cells of patients with asthma. Am. J. Respir. Cell Mol. Biol. 1993, 9, 279–286. [Google Scholar] [CrossRef]
- Walker, C.; Virchow, J.C., Jr.; Bruijnzeel, P.L.; Blaser, K. T cell subsets and their soluble products regulate eosinophilia in allergic and nonallergic asthma. J. Immunol. 1991, 146, 1829–1835. [Google Scholar]
- Corrigan, C.J.; Haczku, A.; Gemou-Engesaeth, V.; Doi, S.; Kikuchi, Y.; Takatsu, K.; Durham, S.R.; Kay, A.B. CD4 T-lymphocyte activation in asthma is accompanied by increased serum concentrations of interleukin-5. Effect of glucocorticoid therapy. Am. Rev. Respir. Dis. 1993, 147, 540–547. [Google Scholar] [CrossRef]
- Nakajima, H.; Iwamoto, I.; Tomoe, S.; Matsumura, R.; Tomioka, H.; Takatsu, K.; Yoshida, S. CD4+ T-lymphocytes and interleukin-5 mediate antigen-induced eosinophil infiltration into the mouse trachea. Am. Rev. Respir. Dis. 1992, 146, 374–377. [Google Scholar] [CrossRef]
- Ishizaka, K.; Ishizaka, T.; Hornbrook, M.M. Physicochemical properties of reaginic antibody. V. Correlation of reaginic activity wth gamma-E-globulin antibody. J. Immunol. 1966, 97, 840–853. [Google Scholar]
- Milgrom, H.; Berger, W.; Nayak, A.; Gupta, N.; Pollard, S.; McAlary, M.; Taylor, A.F.; Rohane, P. Treatment of childhood asthma with anti-immunoglobulin E antibody (omalizumab). Pediatrics 2001, 108, E36. [Google Scholar] [CrossRef] [PubMed]
- Buhl, R.; Soler, M.; Matz, J.; Townley, R.; O’Brien, J.; Noga, O.; Champain, K.; Fox, H.; Thirlwell, J.; Della Cioppa, G. Omalizumab provides long-term control in patients with moderate-to-severe allergic asthma. Eur. Respir. J. 2002, 20, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Cabrera, P.; Berkman, N.; Buhl, R.; Holgate, S.; Wenzel, S.; Fox, H.; Hedgecock, S.; Blogg, M.; Cioppa, G.D. The effect of treatment with omalizumab, an anti-IgE antibody, on asthma exacerbations and emergency medical visits in patients with severe persistent asthma. Allergy 2005, 60, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Holgate, S.T.; Djukanovic, R.; Casale, T.; Bousquet, J. Anti-immunoglobulin E treatment with omalizumab in allergic diseases: An update on anti-inflammatory activity and clinical efficacy. Clin. Exp. Allergy 2005, 35, 408–416. [Google Scholar] [CrossRef]
- Metz, M.; Vadasz, Z.; Kocaturk, E.; Gimenez-Arnau, A.M. Omalizumab Updosing in Chronic Spontaneous Urticaria: An Overview of Real-World Evidence. Clin. Rev. Allergy Immunol. 2020, 59, 38–45. [Google Scholar] [CrossRef]
- Okudaira, H.; Komagata, Y.; Ogita, T. T cell dependent and independent steps in IgE-B memory cell development. Int. Arch. Allergy Appl. Immunol. 1980, 63, 284–293. [Google Scholar] [CrossRef]
- Nehls, M.; Pfeifer, D.; Schorpp, M.; Hedrich, H.; Boehm, T. New member of the winged-helix protein family disrupted in mouse and rat nude mutations. Nature 1994, 372, 103–107. [Google Scholar] [CrossRef]
- Bulfone-Paus, S.; Bahri, R. Mast Cells as Regulators of T Cell Responses. Front. Immunol. 2015, 6, 394. [Google Scholar] [CrossRef]
- Kitamura, Y.; Go, S.; Hatanaka, K. Decrease of mast cells in W/Wv mice and their increase by bone marrow transplantation. Blood 1978, 52, 447–452. [Google Scholar] [CrossRef]
- Kung, T.T.; Stelts, D.; Zurcher, J.A.; Jones, H.; Umland, S.P.; Kreutner, W.; Egan, R.W.; Chapman, R.W. Mast cells modulate allergic pulmonary eosinophilia in mice. Am. J. Respir. Cell Mol. Biol. 1995, 12, 404–409. [Google Scholar] [CrossRef]
- Nagai, H.; Yamaguchi, S.; Maeda, Y.; Tanaka, H. Role of mast cells, eosinophils and IL-5 in the development of airway hyperresponsiveness in sensitized mice. Clin. Exp. Allergy 1996, 26, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Hamelmann, E.; Joetham, A.; Shultz, L.D.; Larsen, G.L.; Irvin, C.G.; Gelfand, E.W. Development of eosinophilic airway inflammation and airway hyperresponsiveness in mast cell-deficient mice. J. Exp. Med. 1997, 186, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Kaminuma, O.; Kikkawa, H.; Nakata, A.; Asahina, M.; Egan, R.W.; Akiyama, K.; Mori, A. Transient contribution of mast cells to pulmonary eosinophilia but not to hyper-responsiveness. Clin. Exp. Allergy 2002, 32, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Komai, M.; Tanaka, H.; Masuda, T.; Nagao, K.; Ishizaki, M.; Sawada, M.; Nagai, H. Role of Th2 responses in the development of allergen-induced airway remodelling in a murine model of allergic asthma. Br. J. Pharmacol. 2003, 138, 912–920. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, I.; Tomoe, S.; Tomioka, H.; Takatsu, K.; Yoshida, S. Role of CD4+ T lymphocytes and interleukin-5 in antigen-induced eosinophil recruitment into the site of cutaneous late-phase reaction in mice. J. Leukoc. Biol. 1992, 52, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Mukai, K.; Matsuoka, K.; Taya, C.; Suzuki, H.; Yokozeki, H.; Nishioka, K.; Hirokawa, K.; Etori, M.; Yamashita, M.; Kubota, T.; et al. Basophils play a critical role in the development of IgE-mediated chronic allergic inflammation independently of T cells and mast cells. Immunity 2005, 23, 191–202. [Google Scholar] [CrossRef]
- Poddighe, D.; Mathias, C.B.; Brambilla, I.; Marseglia, G.L.; Oettgen, H.C. Importance of Basophils in Eosinophilic Asthma: The Murine Counterpart. J. Biol. Reg. Homeos. Ag. 2018, 32, 335–339. [Google Scholar]
- Nishimura, T.; Saeki, M.; Kaminuma, O.; Matsuoka, K.; Yonekawa, H.; Mori, A.; Hiroi, T. Existence of antigen-specific immunoglobulin E is not sufficient for allergic nasal eosinophil infiltration in mice. Int. Arch. Allergy Immunol. 2013, 161 (Suppl. 2), 125–128. [Google Scholar] [CrossRef]
- Kaminuma, O.; Mori, A.; Ogawa, K.; Nakata, A.; Kikkawa, H.; Naito, K.; Suko, M.; Okudaira, H. Successful transfer of late phase eosinophil infiltration in the lung by infusion of helper T cell clones. Am. J. Respir. Cell Mol. Biol. 1997, 16, 448–454. [Google Scholar] [CrossRef]
- Randolph, D.A.; Stephens, R.; Carruthers, C.J.; Chaplin, D.D. Cooperation between Th1 and Th2 cells in a murine model of eosinophilic airway inflammation. J. Clin. Investig. 1999, 104, 1021–1029. [Google Scholar] [CrossRef]
- Kaminuma, O.; Mori, A. Potential initiation of eosinophilic skin inflammation by antigen-specific T helper type 2 cells. Int. Arch. Allergy Immunol. 2002, 128 (Suppl. 1), 21–23. [Google Scholar] [CrossRef]
- Nishimura, T.; Kaminuma, O.; Saeki, M.; Kitamura, N.; Matsuoka, K.; Yonekawa, H.; Mori, A.; Hiroi, T. Essential Contribution of CD4+ T Cells to Antigen-Induced Nasal Hyperresponsiveness in Experimental Allergic Rhinitis. PLoS ONE 2016, 11, e0146686. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Kaminuma, O.; Kitamura, N.; Hiroi, T. Induced Treg Cells Augment the Th17-Mediated Intestinal Inflammatory Response in a CTLA4-Dependent Manner. PLoS ONE 2016, 11, e0150244. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.S.; Hamid, Q.; Ying, S.; Tsicopoulos, A.; Barkans, J.; Bentley, A.M.; Corrigan, C.; Durham, S.R.; Kay, A.B. Predominant TH2-like bronchoalveolar T-lymphocyte population in atopic asthma. N. Eng. J. Med. 1992, 326, 298–304. [Google Scholar] [CrossRef] [PubMed]
- van Reijsen, F.C.; Bruijnzeel-Koomen, C.A.; Kalthoff, F.S.; Maggi, E.; Romagnani, S.; Westland, J.K.; Mudde, G.C. Skin-derived aeroallergen-specific T-cell clones of Th2 phenotype in patients with atopic dermatitis. J. Allergy Clin. Immunol. 1992, 90, 184–193. [Google Scholar] [CrossRef]
- Bousquet, J.; Chanez, P.; Lacoste, J.Y.; Barneon, G.; Ghavanian, N.; Enander, I.; Venge, P.; Ahlstedt, S.; Simony-Lafontaine, J.; Godard, P.; et al. Eosinophilic inflammation in asthma. N. Eng. J. Med. 1990, 323, 1033–1039. [Google Scholar] [CrossRef]
- Kay, A.B. Asthma and inflammation. J. Allergy Clin. Immunol. 1991, 87, 893–910. [Google Scholar] [CrossRef]
- Bochner, B.S.; Undem, B.J.; Lichtenstein, L.M. Immunological aspects of allergic asthma. Annu. Rev. Immunol. 1994, 12, 295–335. [Google Scholar] [CrossRef]
- Wills-Karp, M. Immunologic basis of antigen-induced airway hyperresponsiveness. Annu. Rev. Immunol. 1999, 17, 255–281. [Google Scholar] [CrossRef]
- Garcia, G.; Taille, C.; Laveneziana, P.; Bourdin, A.; Chanez, P.; Humbert, M. Anti-interleukin-5 therapy in severe asthma. Eur. Respir. Rev. 2013, 22, 251–257. [Google Scholar] [CrossRef]
- Price, D.B.; Rigazio, A.; Campbell, J.D.; Bleecker, E.R.; Corrigan, C.J.; Thomas, M.; Wenzel, S.E.; Wilson, A.M.; Small, M.B.; Gopalan, G.; et al. Blood eosinophil count and prospective annual asthma disease burden: A UK cohort study. Lancet Respir. Med. 2015, 3, 849–858. [Google Scholar] [CrossRef]
- McBrien, C.N.; Menzies-Gow, A. The Biology of Eosinophils and Their Role in Asthma. Front. Med. (Lausanne) 2017, 4, 93. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.; Bode, E.; Boer, L.; Hansel, T.T.; Blaser, K.; Virchow, J.C., Jr. Allergic and nonallergic asthmatics have distinct patterns of T-cell activation and cytokine production in peripheral blood and bronchoalveolar lavage. Am. Rev. Respir. Dis. 1992, 146, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.; Bauer, W.; Braun, R.K.; Menz, G.; Braun, P.; Schwarz, F.; Hansel, T.T.; Villiger, B. Activated T cells and cytokines in bronchoalveolar lavages from patients with various lung diseases associated with eosinophilia. Am. J. Respir. Crit. Care Med. 1994, 150, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Truyen, E.; Coteur, L.; Dilissen, E.; Overbergh, L.; Dupont, L.J.; Ceuppens, J.L.; Bullens, D.M. Evaluation of airway inflammation by quantitative Th1/Th2 cytokine mRNA measurement in sputum of asthma patients. Thorax 2006, 61, 202–208. [Google Scholar] [CrossRef]
- Mori, A.; Suko, M.; Nishizaki, Y.; Kaminuma, O.; Kobayashi, S.; Matsuzaki, G.; Yamamoto, K.; Ito, K.; Tsuruoka, N.; Okudaira, H. IL-5 production by CD4+ T cells of asthmatic patients is suppressed by glucocorticoids and the immunosuppressants FK506 and cyclosporin A. Int. Immunol. 1995, 7, 449–457. [Google Scholar] [CrossRef]
- Nakata, A.; Kaminuma, O.; Ogawa, K.; Fujimura, H.; Fushimi, K.; Kikkawa, H.; Naito, K.; Ikezawa, K.; Egan, R.W.; Mori, A. Correlation between eosinophilia induced by CD4(+) T cells and bronchial hyper-responsiveness. Int. Immunol. 2001, 13, 329–339. [Google Scholar] [CrossRef]
- Humbles, A.A.; Lloyd, C.M.; McMillan, S.J.; Friend, D.S.; Xanthou, G.; McKenna, E.E.; Ghiran, S.; Gerard, N.P.; Yu, C.; Orkin, S.H.; et al. A critical role for eosinophils in allergic airways remodeling. Science 2004, 305, 1776–1779. [Google Scholar] [CrossRef]
- Lee, J.J.; Dimina, D.; Macias, M.P.; Ochkur, S.I.; McGarry, M.P.; O’Neill, K.R.; Protheroe, C.; Pero, R.; Nguyen, T.; Cormier, S.A.; et al. Defining a link with asthma in mice congenitally deficient in eosinophils. Science 2004, 305, 1773–1776. [Google Scholar] [CrossRef]
- Saeki, M.; Kaminuma, O.; Nishimura, T.; Kitamura, N.; Mori, A.; Hiroi, T. Th9 cells elicit eosinophil-independent bronchial hyperresponsiveness in mice. Allergol. Int. 2016, 65 (Suppl. 1), 24–29. [Google Scholar] [CrossRef]
- Li, X.M.; Chopra, R.K.; Chou, T.Y.; Schofield, B.H.; Wills-Karp, M.; Huang, S.K. Mucosal IFN-gamma gene transfer inhibits pulmonary allergic responses in mice. J. Immunol. 1996, 157, 3216–3219. [Google Scholar] [PubMed]
- Cohn, L.; Homer, R.J.; Niu, N.; Bottomly, K. T helper 1 cells and interferon gamma regulate allergic airway inflammation and mucus production. J. Exp. Med. 1999, 190, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Hansen, G.; Berry, G.; DeKruyff, R.H.; Umetsu, D.T. Allergen-specific Th1 cells fail to counterbalance Th2 cell-induced airway hyperreactivity but cause severe airway inflammation. J. Clin. Investig. 1999, 103, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Dardalhon, V.; Awasthi, A.; Kwon, H.; Galileos, G.; Gao, W.; Sobel, R.A.; Mitsdoerffer, M.; Strom, T.B.; Elyaman, W.; Ho, I.C.; et al. IL-4 inhibits TGF-beta-induced Foxp3+ T cells and, together with TGF-beta, generates IL-9+ IL-10+ Foxp3(-) effector T cells. Nat. Immunol. 2008, 9, 1347–1355. [Google Scholar] [CrossRef]
- Veldhoen, M.; Uyttenhove, C.; van Snick, J.; Helmby, H.; Westendorf, A.; Buer, J.; Martin, B.; Wilhelm, C.; Stockinger, B. Transforming growth factor-beta ‘reprograms’ the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset. Nat. Immunol. 2008, 9, 1341–1346. [Google Scholar] [CrossRef]
- Micosse, C.; von Meyenn, L.; Steck, O.; Kipfer, E.; Adam, C.; Simillion, C.; Seyed Jafari, S.M.; Olah, P.; Yawlkar, N.; Simon, D.; et al. Human “TH9” cells are a subpopulation of PPAR-gamma(+) TH2 cells. Sci. Immunol. 2019, 4, eaat5943. [Google Scholar] [CrossRef]
- Harrington, L.E.; Hatton, R.D.; Mangan, P.R.; Turner, H.; Murphy, T.L.; Murphy, K.M.; Weaver, C.T. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat. Immunol. 2005, 6, 1123–1132. [Google Scholar] [CrossRef]
- Langrish, C.L.; Chen, Y.; Blumenschein, W.M.; Mattson, J.; Basham, B.; Sedgwick, J.D.; McClanahan, T.; Kastelein, R.A.; Cua, D.J. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J. Exp. Med. 2005, 201, 233–240. [Google Scholar] [CrossRef]
- Liang, S.C.; Tan, X.Y.; Luxenberg, D.P.; Karim, R.; Dunussi-Joannopoulos, K.; Collins, M.; Fouser, L.A. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J. Exp. Med. 2006, 203, 2271–2279. [Google Scholar] [CrossRef]
- Eyerich, S.; Eyerich, K.; Pennino, D.; Carbone, T.; Nasorri, F.; Pallotta, S.; Cianfarani, F.; Odorisio, T.; Traidl-Hoffmann, C.; Behrendt, H.; et al. Th22 cells represent a distinct human T cell subset involved in epidermal immunity and remodeling. J. Clin. Investig. 2009, 119, 3573–3585. [Google Scholar] [CrossRef]
- Tato, C.M.; Laurence, A.; O’Shea, J.J. Helper T cell differentiation enters a new era: Le roi est mort; vive le roi! J. Exp. Med. 2006, 203, 809–812. [Google Scholar] [CrossRef] [PubMed]
- Szegedi, K.; van Lier, A.; Res, P.C.; Chielie, S.; Bos, J.D.; Kezic, S.; Middelkamp-Hup, M.A.; Luiten, R.M. House dust mite allergens Der f and Der p induce IL-31 production by blood-derived T cells from atopic dermatitis patients. Exp. Dermatol. 2018, 27, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Alvarez, B.; Rodriguez, R.M.; Fraga, M.F.; Lopez-Larrea, C. DNA methylation: A promising landscape for immune system-related diseases. Trends. Genet. 2012, 28, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Potaczek, D.P.; Harb, H.; Michel, S.; Alhamwe, B.A.; Renz, H.; Tost, J. Epigenetics and allergy: From basic mechanisms to clinical applications. Epigenomics 2017, 9, 539–571. [Google Scholar] [CrossRef]
- McKinley, L.; Alcorn, J.F.; Peterson, A.; Dupont, R.B.; Kapadia, S.; Logar, A.; Henry, A.; Irvin, C.G.; Piganelli, J.D.; Ray, A.; et al. TH17 cells mediate steroid-resistant airway inflammation and airway hyperresponsiveness in mice. J. Immunol. 2008, 181, 4089–4097. [Google Scholar] [CrossRef]
- Kaminuma, O.; Ohtomo, T.; Mori, A.; Nagakubo, D.; Hieshima, K.; Ohmachi, Y.; Noda, Y.; Katayama, K.; Suzuki, K.; Motoi, Y.; et al. Selective down-regulation of Th2 cell-mediated airway inflammation in mice by pharmacological intervention of CCR4. Clin. Exp. Allergy 2012, 42, 315–325. [Google Scholar] [CrossRef]
- Saeki, M.; Kaminuma, O.; Nishimura, T.; Kitamura, N.; Mori, A.; Hiroi, T. Th9 cells induce steroid-resistant bronchial hyperresponsiveness in mice. Allergol. Int. 2017, 66, S35–S40. [Google Scholar] [CrossRef]
- Ying, S.; Durham, S.R.; Corrigan, C.J.; Hamid, Q.; Kay, A.B. Phenotype of cells expressing mRNA for TH2-type (interleukin 4 and interleukin 5) and TH1-type (interleukin 2 and interferon gamma) cytokines in bronchoalveolar lavage and bronchial biopsies from atopic asthmatic and normal control subjects. Am. J. Respir. Cell Mol. Biol. 1995, 12, 477–487. [Google Scholar] [CrossRef]
- Pene, J.; Chevalier, S.; Preisser, L.; Venereau, E.; Guilleux, M.H.; Ghannam, S.; Moles, J.P.; Danger, Y.; Ravon, E.; Lesaux, S.; et al. Chronically inflamed human tissues are infiltrated by highly differentiated Th17 lymphocytes. J. Immunol. 2008, 180, 7423–7430. [Google Scholar] [CrossRef]
- Hoppenot, D.; Malakauskas, K.; Lavinskiene, S.; Bajoriuniene, I.; Kalinauskaite, V.; Sakalauskas, R. Peripheral blood Th9 cells and eosinophil apoptosis in asthma patients. Medicina (Kaunas) 2015, 51, 10–17. [Google Scholar] [CrossRef]
- Jia, L.; Wang, Y.; Li, J.; Li, S.; Zhang, Y.; Shen, J.; Tan, W.; Wu, C. Detection of IL-9 producing T cells in the PBMCs of allergic asthmatic patients. BMC Immunol. 2017, 18, 38. [Google Scholar] [CrossRef] [PubMed]
- Murdaca, G.; Colombo, B.M.; Puppo, F. The role of Th17 lymphocytes in the autoimmune and chronic inflammatory diseases. Intern. Emerg. Med. 2011, 6, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.L.; Chen, Z. Establishment of different experimental asthma models in mice. Exp. Ther. Med. 2018, 15, 2492–2498. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.T.; Hagner, S.; Ruchti, F.; Radzikowska, U.; Tan, G.; Altunbulakli, C.; Eljaszewicz, A.; Moniuszko, M.; Akdis, M.; Akdis, C.A.; et al. Tight junction, mucin, and inflammasome-related molecules are differentially expressed in eosinophilic, mixed, and neutrophilic experimental asthma in mice. Allergy 2019, 74, 294–307. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Sakaguchi, N.; Asano, M.; Itoh, M.; Toda, M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol. 1995, 155, 1151–1164. [Google Scholar]
- Palomares, O.; Akdis, M.; Martin-Fontecha, M.; Akdis, C.A. Mechanisms of immune regulation in allergic diseases: The role of regulatory T and B cells. Immunol. Rev. 2017, 278, 219–236. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H. The immunology of asthma. Nat. Immunol. 2015, 16, 45–56. [Google Scholar] [CrossRef]
- Gandhi, N.A.; Bennett, B.L.; Graham, N.M.; Pirozzi, G.; Stahl, N.; Yancopoulos, G.D. Targeting key proximal drivers of type 2 inflammation in disease. Nat. Rev. Drug. Discov. 2016, 15, 35–50. [Google Scholar] [CrossRef]
- Miethe, S.; Guarino, M.; Alhamdan, F.; Simon, H.U.; Renz, H.; Dufour, J.F.; Potaczek, D.P.; Garn, H. Effects of obesity on asthma: Immunometabolic links. Pol. Arch. Intern. Med. 2018, 128, 469–477. [Google Scholar] [CrossRef]
- Jonckheere, A.C.; Bullens, D.M.A.; Seys, S.F. Innate lymphoid cells in asthma: Pathophysiological insights from murine models to human asthma phenotypes. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 53–60. [Google Scholar] [CrossRef]
- Potaczek, D.P.; Miethe, S.; Schindler, V.; Alhamdan, F.; Garn, H. Role of airway epithelial cells in the development of different asthma phenotypes. Cell Signal 2020, 69, 109523. [Google Scholar] [CrossRef] [PubMed]
- Akdis, C.A.; Bachert, C.; Cingi, C.; Dykewicz, M.S.; Hellings, P.W.; Naclerio, R.M.; Schleimer, R.P.; Ledford, D. Endotypes and phenotypes of chronic rhinosinusitis: A PRACTALL document of the European Academy of Allergy and Clinical Immunology and the American Academy of Allergy, Asthma & Immunology. J. Allergy Clin. Immunol. 2013, 131, 1479–1490. [Google Scholar] [PubMed]
- Agache, I.; Akdis, C.A. Endotypes of allergic diseases and asthma: An important step in building blocks for the future of precision medicine. Allergol. Int. 2016, 65, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Barnden, M.J.; Allison, J.; Heath, W.R.; Carbone, F.R. Defective TCR expression in transgenic mice constructed using cDNA-based alpha- and beta-chain genes under the control of heterologous regulatory elements. Immunol. Cell Biol. 1998, 76, 34–40. [Google Scholar] [CrossRef]
- Murphy, K.M.; Heimberger, A.B.; Loh, D.Y. Induction by antigen of intrathymic apoptosis of CD4+CD8+TCRlo thymocytes in vivo. Science 1990, 250, 1720–1723. [Google Scholar] [CrossRef]
- Nishimura, T.; Saeki, M.; Motoi, Y.; Kitamura, N.; Mori, A.; Kaminuma, O.; Hiroi, T. Selective suppression of Th2 cell-mediated lung eosinophilic inflammation by anti-major facilitator super family domain containing 10 monoclonal antibody. Allergol. Int. 2014, 63 (Suppl. 1), 29–35. [Google Scholar] [CrossRef]
- Kaminuma, O.; Nishimura, T.; Kitamura, N.; Saeki, M.; Hiroi, T.; Mori, A. T-Helper Type 2 Cells Direct Antigen-Induced Eosinophilic Skin Inflammation in Mice. Allergy Asthma Immunol. Res. 2018, 10, 77–82. [Google Scholar] [CrossRef]
- Sato, T.; Sasahara, T.; Nakamura, Y.; Osaki, T.; Hasegawa, T.; Tadakuma, T.; Arata, Y.; Kumagai, Y.; Katsuki, M.; Habu, S. Naive T cells can mediate delayed-type hypersensitivity response in T cell receptor transgenic mice. Eur. J. Immunol. 1994, 24, 1512–1516. [Google Scholar] [CrossRef]
- Nakajima-Adachi, H.; Ebihara, A.; Kikuchi, A.; Ishida, T.; Sasaki, K.; Hirano, K.; Watanabe, H.; Asai, K.; Takahashi, Y.; Kanamori, Y.; et al. Food antigen causes TH2-dependent enteropathy followed by tissue repair in T-cell receptor transgenic mice. J. Allergy Clin. Immunol. 2006, 117, 1125–1132. [Google Scholar] [CrossRef]
- Nakajima-Adachi, H.; Kikuchi, A.; Fujimura, Y.; Shibahara, K.; Makino, T.; Goseki-Sone, M.; Kihara-Fujioka, M.; Nochi, T.; Kurashima, Y.; Igarashi, O.; et al. Peyer’s patches and mesenteric lymph nodes cooperatively promote enteropathy in a mouse model of food allergy. PLoS ONE 2014, 9, e107492. [Google Scholar] [CrossRef]
- Nakajima-Adachi, H.; Koike, E.; Totsuka, M.; Hiraide, E.; Wakatsuki, Y.; Kiyono, H.; Hachimura, S. Two distinct epitopes on the ovalbumin 323-339 peptide differentiating CD4(+)T cells into the Th2 or Th1 phenotype. Biosci. Biotechnol. Biochem. 2012, 76, 1979–1981. [Google Scholar] [CrossRef][Green Version]
- Jarman, E.R.; Tan, K.A.; Lamb, J.R. Transgenic mice expressing the T cell antigen receptor specific for an immunodominant epitope of a major allergen of house dust mite develop an asthmatic phenotype on exposure of the airways to allergen. Clin. Exp. Allergy 2005, 35, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, M.M.; Dumoutier, L.; Warnier, G.; Uyttenhove, C.; Van Snick, J.; de Heusch, M.; Stevens, M.; Renauld, J.C. Dual TCR expression biases lung inflammation in DO11.10 transgenic mice and promotes neutrophilia via microbiota-induced Th17 differentiation. J. Immunol. 2011, 187, 3530–3537. [Google Scholar] [CrossRef] [PubMed]
- Constant, S.; Pfeiffer, C.; Woodard, A.; Pasqualini, T.; Bottomly, K. Extent of T cell receptor ligation can determine the functional differentiation of naive CD4+ T cells. J. Exp. Med. 1995, 182, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Hosken, N.A.; Shibuya, K.; Heath, A.W.; Murphy, K.M.; O’Garra, A. The effect of antigen dose on CD4+ T helper cell phenotype development in a T cell receptor-alpha beta-transgenic model. J. Exp. Med. 1995, 182, 1579–1584. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.; Barker, D.J.; Georgiou, X.; Cooper, M.A.; Flicek, P.; Marsh, S.G.E. IPD-IMGT/HLA Database. Nucleic Acids Res. 2020, 48, D948–D955. [Google Scholar] [PubMed]
- Wilmut, I.; Schnieke, A.E.; McWhir, J.; Kind, A.J.; Campbell, K.H. Viable offspring derived from fetal and adult mammalian cells. Nature 1997, 385, 810–813. [Google Scholar] [CrossRef] [PubMed]
- Wakayama, T.; Perry, A.C.; Zuccotti, M.; Johnson, K.R.; Yanagimachi, R. Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature 1998, 394, 369–374. [Google Scholar] [CrossRef]
- Hochedlinger, K.; Jaenisch, R. Monoclonal mice generated by nuclear transfer from mature B and T donor cells. Nature 2002, 415, 1035–1038. [Google Scholar] [CrossRef]
- Serwold, T.; Hochedlinger, K.; Inlay, M.A.; Jaenisch, R.; Weissman, I.L. Early TCR expression and aberrant T cell development in mice with endogenous prerearranged T cell receptor genes. J. Immunol. 2007, 179, 928–938. [Google Scholar] [CrossRef]
- Kirak, O.; Frickel, E.M.; Grotenbreg, G.M.; Suh, H.; Jaenisch, R.; Ploegh, H.L. Transnuclear mice with predefined T cell receptor specificities against Toxoplasma gondii obtained via SCNT. Science 2010, 328, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Wakao, H.; Ogonuki, N.; Miki, H.; Seino, K.; Nambu-Wakao, R.; Noda, S.; Miyoshi, H.; Koseki, H.; Taniguchi, M.; et al. Generation of cloned mice by direct nuclear transfer from natural killer T cells. Curr. Biol. 2005, 15, 1114–1118. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, S.; Inoue, K.; Ogonuki, N.; Hirose, M.; Oikawa, M.; Yo, M.; Ohara, O.; Miyoshi, H.; Ogura, A. Mouse cloning using a drop of peripheral blood. Biol. Reprod. 2013, 89, 24. [Google Scholar] [CrossRef] [PubMed]
- Eggan, K.; Akutsu, H.; Loring, J.; Jackson-Grusby, L.; Klemm, M.; Rideout, W.M., 3rd; Yanagimachi, R.; Jaenisch, R. Hybrid vigor, fetal overgrowth, and viability of mice derived by nuclear cloning and tetraploid embryo complementation. Proc. Natl. Acad. Sci. USA 2001, 98, 6209–6214. [Google Scholar] [CrossRef] [PubMed]
- Ogura, A.; Inoue, K.; Wakayama, T. Recent advancements in cloning by somatic cell nuclear transfer. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20110329. [Google Scholar] [CrossRef] [PubMed]
- Kaminuma, O.; Katayama, K.; Inoue, K.; Saeki, M.; Nishimura, T.; Kitamura, N.; Shimo, Y.; Tofukuji, S.; Ishida, S.; Ogonuki, N.; et al. Hyper-reactive cloned mice generated by direct nuclear transfer of antigen-specific CD4(+) T cells. EMBO Rep. 2017, 18, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Tubo, N.J.; Pagan, A.J.; Taylor, J.J.; Nelson, R.W.; Linehan, J.L.; Ertelt, J.M.; Huseby, E.S.; Way, S.S.; Jenkins, M.K. Single naive CD4+ T cells from a diverse repertoire produce different effector cell types during infection. Cell 2013, 153, 785–796. [Google Scholar] [CrossRef]
- Johnson, J.R.; Wiley, R.E.; Fattouh, R.; Swirski, F.K.; Gajewska, B.U.; Coyle, A.J.; Gutierrez-Ramos, J.C.; Ellis, R.; Inman, M.D.; Jordana, M. Continuous exposure to house dust mite elicits chronic airway inflammation and structural remodeling. Am. J. Respir. Crit. Care Med. 2004, 169, 378–385. [Google Scholar] [CrossRef]
- Ulrich, K.; Hincks, J.S.; Walsh, R.; Wetterstrand, E.M.; Fidock, M.D.; Sreckovic, S.; Lamb, D.J.; Douglas, G.J.; Yeadon, M.; Perros-Huguet, C.; et al. Anti-inflammatory modulation of chronic airway inflammation in the murine house dust mite model. Pulm. Pharmacol. Ther. 2008, 21, 637–647. [Google Scholar] [CrossRef]
- Shimizu, H.; Obase, Y.; Katoh, S.; Mouri, K.; Kobashi, Y.; Oka, M. Critical role of interleukin-5 in the development of a mite antigen-induced chronic bronchial asthma model. Inflamm. Res. 2013, 62, 911–917. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Antigen-Challenged Animals | Antigen-Specific IgE/mast Cells | Antigen-Specific CD4+ T Cells | Eosinophil Infiltration |
|---|---|---|---|
| Non-immunized IgE-Tg | Yes | No | No |
| Immunized wild-type | Yes | Yes | Yes |
| Immunized W/Wv | No | Yes | Yes |
| Immunized wild-type + anti-CD4 antibody | Yes | No | No |
| Immunized W/Wv + anti-CD4 antibody | No | No | No |
| Non-immunized wild-type + antigen-specific CD4+ T cells | No | Yes | Yes |
| Features | Th2 | Th9 | Th1 | Th17 |
|---|---|---|---|---|
| Requirement of IgE/mast cells | No | No | No | No |
| BHR | Yes | Yes | Yes | Yes |
| Eosinophilia | Yes | Yes | No | No |
| Contribution of eosinophils of BHR | Yes | No | No | No |
| Steroid resistance | No | Yes | No | Yes |
| Reactive Antigen | No. of Coned Mice | Requirement of Reconstituted TCRα and/or β for Antigen Reactivity | |||
|---|---|---|---|---|---|
| In Vitro | In Vivo | ||||
| Both rα and rβ | Either rα or rβ | Both rα and rβ | Either rα or rβ | ||
| OVA323-339 | 1 | Yes | N.T. | Yes | Yes |
| Der f | 2 | Yes | No | Yes | Yes |
| Der p | 5 | Yes | No | N.T. | N.T. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miura, K.; Inoue, K.; Ogura, A.; Kaminuma, O. Role of CD4+ T Cells in Allergic Airway Diseases: Learning from Murine Models. Int. J. Mol. Sci. 2020, 21, 7480. https://doi.org/10.3390/ijms21207480
Miura K, Inoue K, Ogura A, Kaminuma O. Role of CD4+ T Cells in Allergic Airway Diseases: Learning from Murine Models. International Journal of Molecular Sciences. 2020; 21(20):7480. https://doi.org/10.3390/ijms21207480
Chicago/Turabian StyleMiura, Kento, Kimiko Inoue, Atsuo Ogura, and Osamu Kaminuma. 2020. "Role of CD4+ T Cells in Allergic Airway Diseases: Learning from Murine Models" International Journal of Molecular Sciences 21, no. 20: 7480. https://doi.org/10.3390/ijms21207480
APA StyleMiura, K., Inoue, K., Ogura, A., & Kaminuma, O. (2020). Role of CD4+ T Cells in Allergic Airway Diseases: Learning from Murine Models. International Journal of Molecular Sciences, 21(20), 7480. https://doi.org/10.3390/ijms21207480