Glutathione Ethyl Ester Protects In Vitro-Maturing Bovine Oocytes against Oxidative Stress Induced by Subsequent Vitrification/Warming

 , , , ,
, , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Results
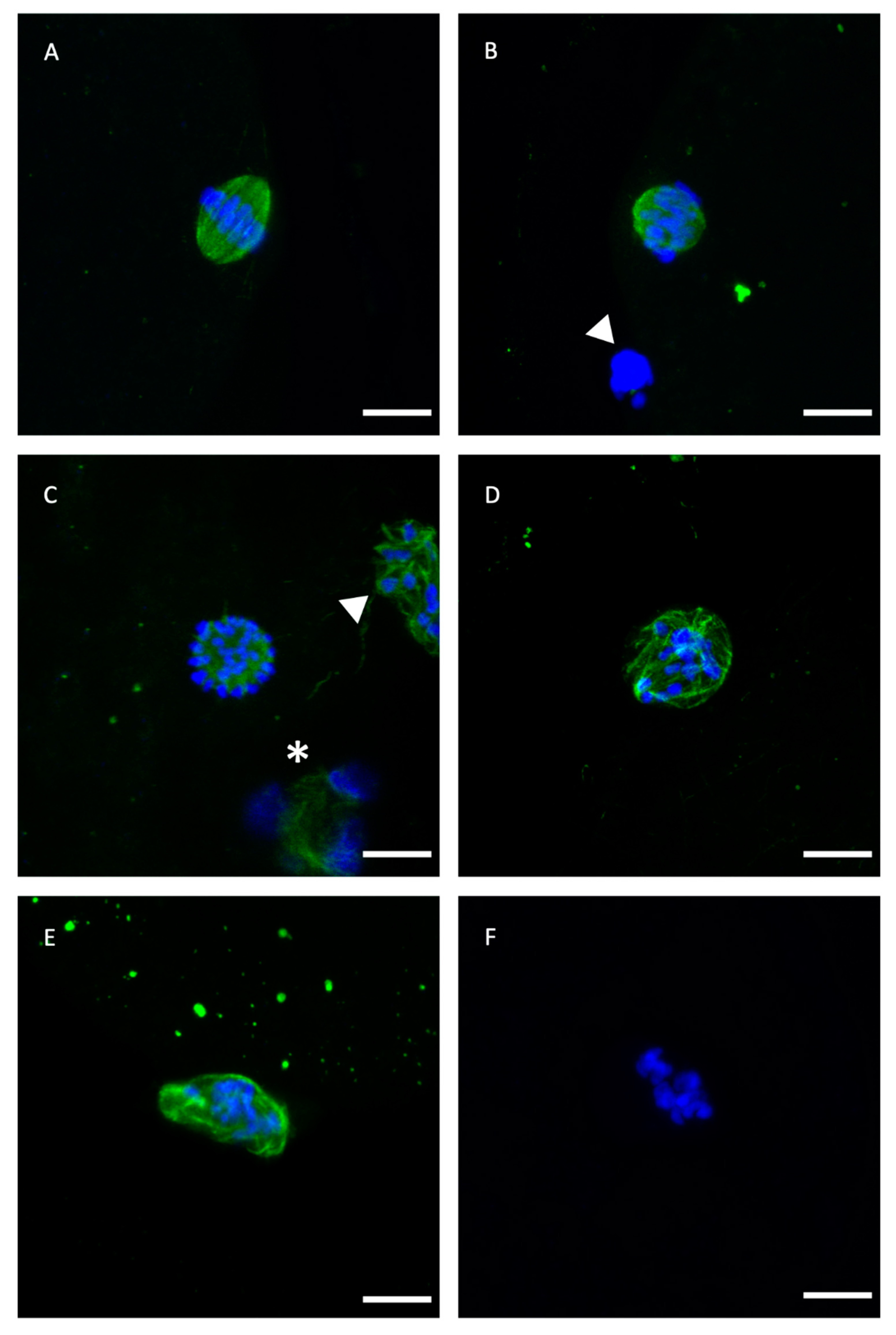
2.1. Meiotic Spindle Status of Vitrified/Warmed Bovine Oocytes after Maturation in IVM Medium Supplemented with GSH-OEt
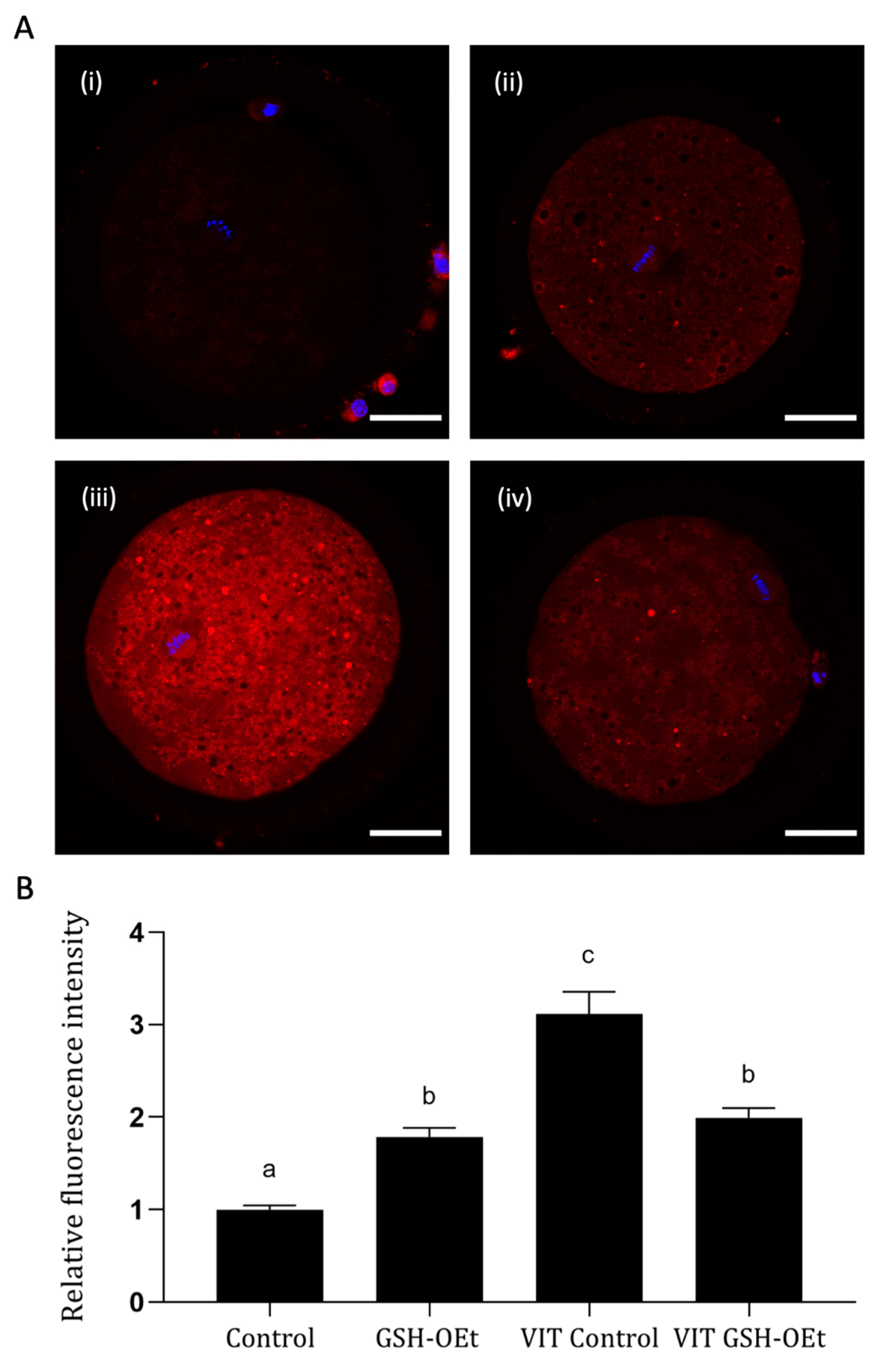
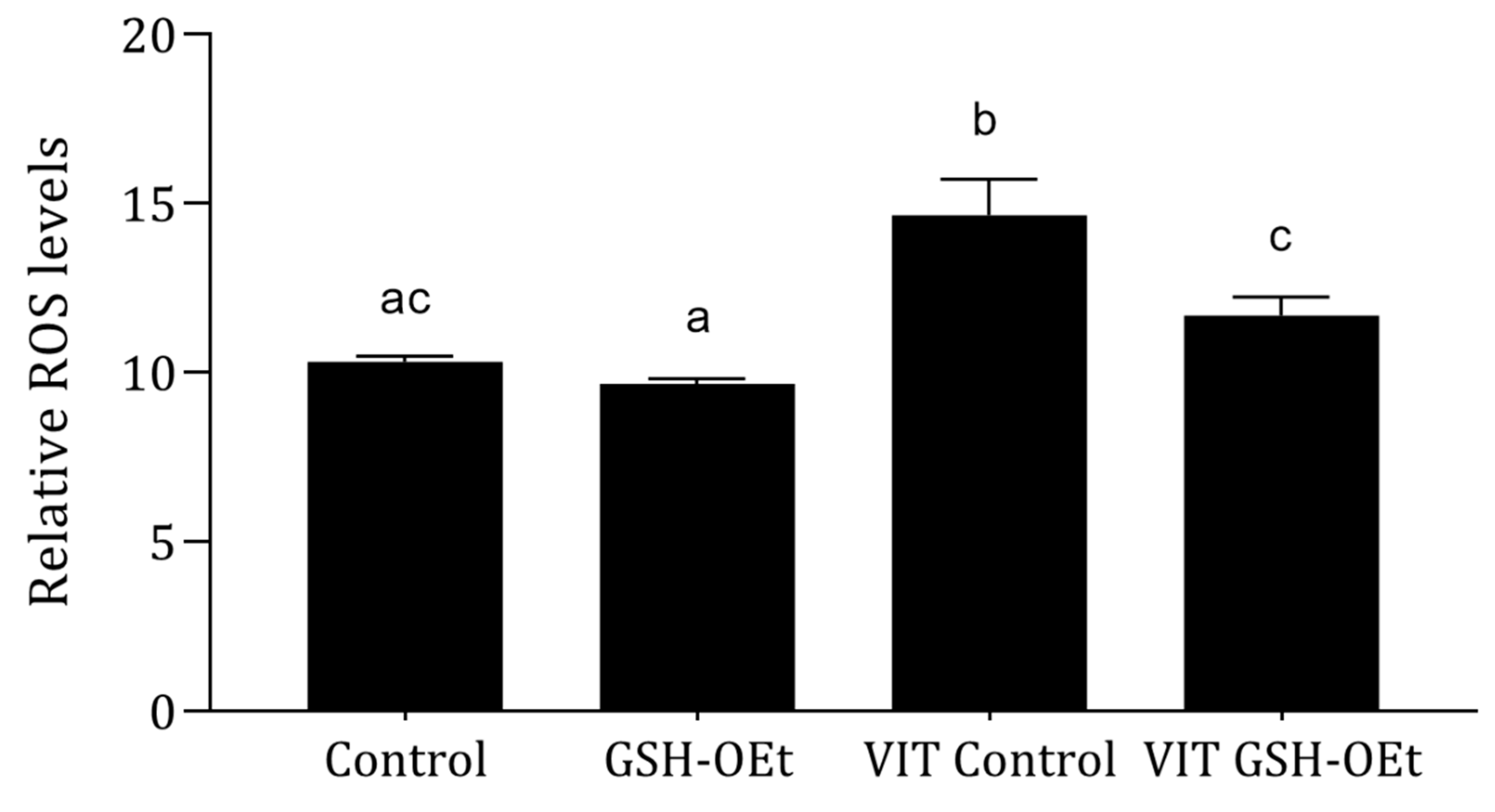
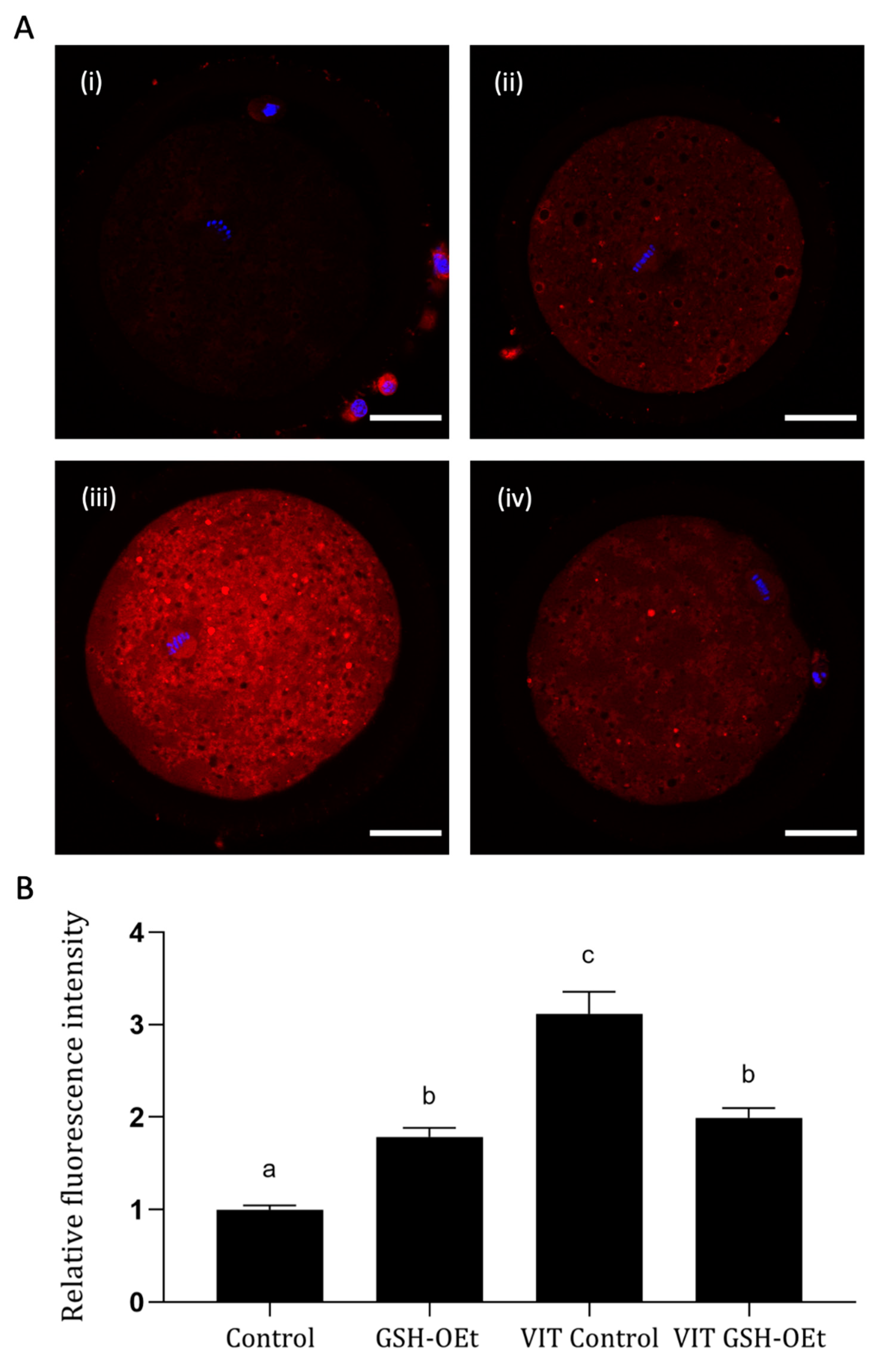
2.2. ROS Production Detected in Vitrified/Warmed Bovine Oocytes after Their Maturation in IVM with or without GSH-OEt
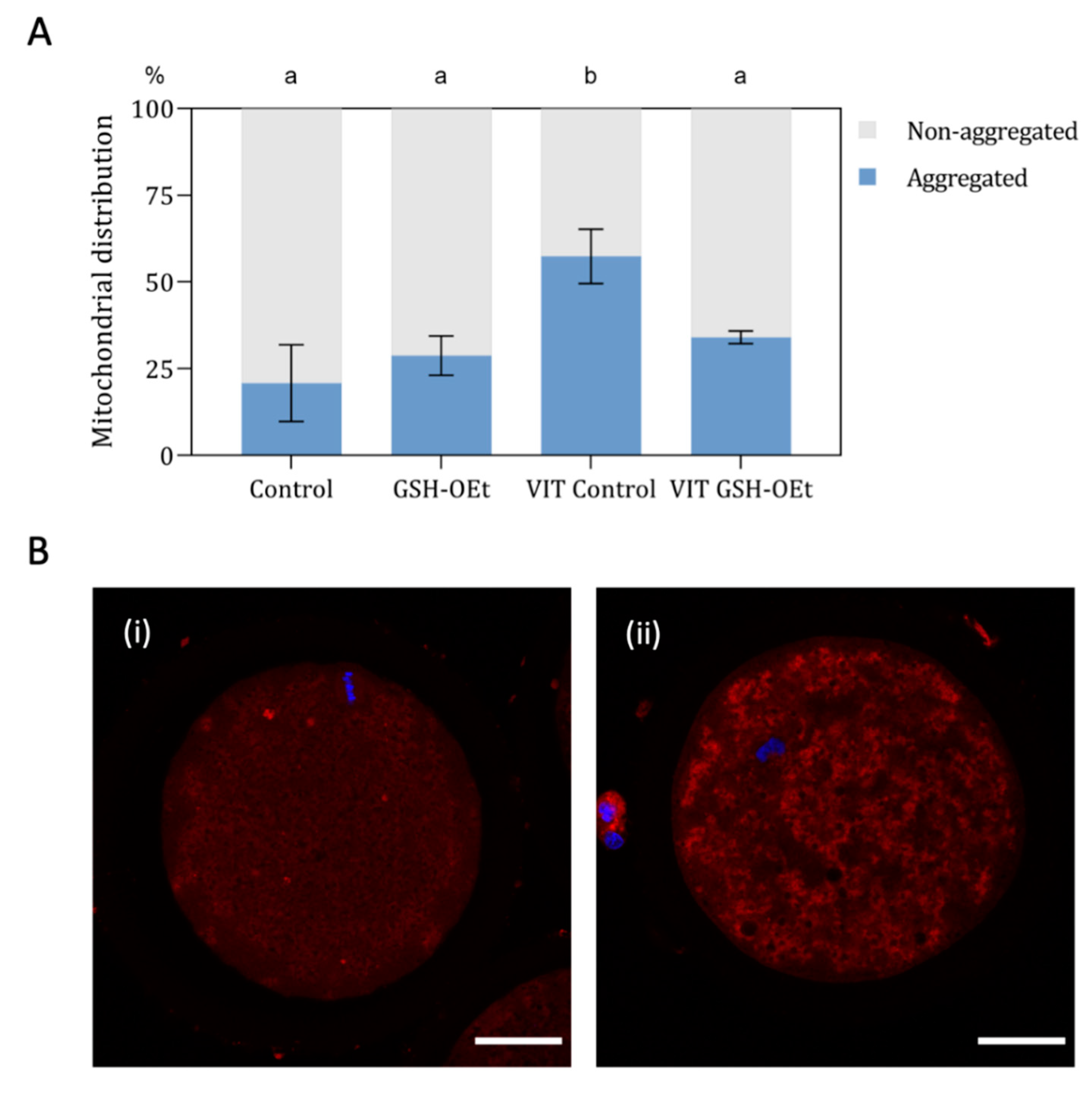
2.3. Mitochondrial Activity and Distribution in Vitrified/Warmed Bovine Oocytes after Their Maturation in IVM Medium Supplemented with GSH-OEt
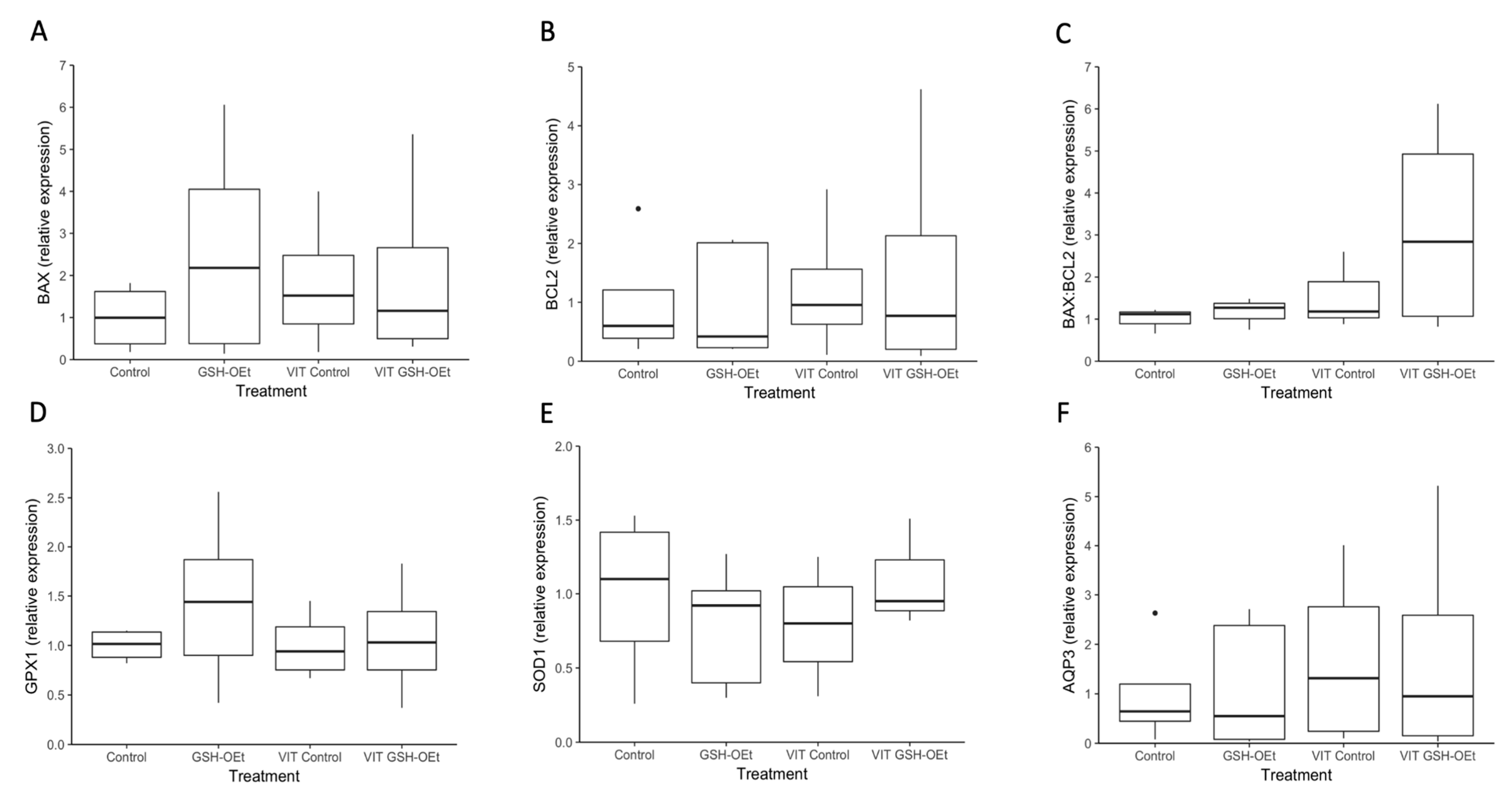
2.4. Gene Expression in Vitrified/Warmed Bovine Oocytes after Their Maturation in IVM Medium Supplemented with GSH-OEt
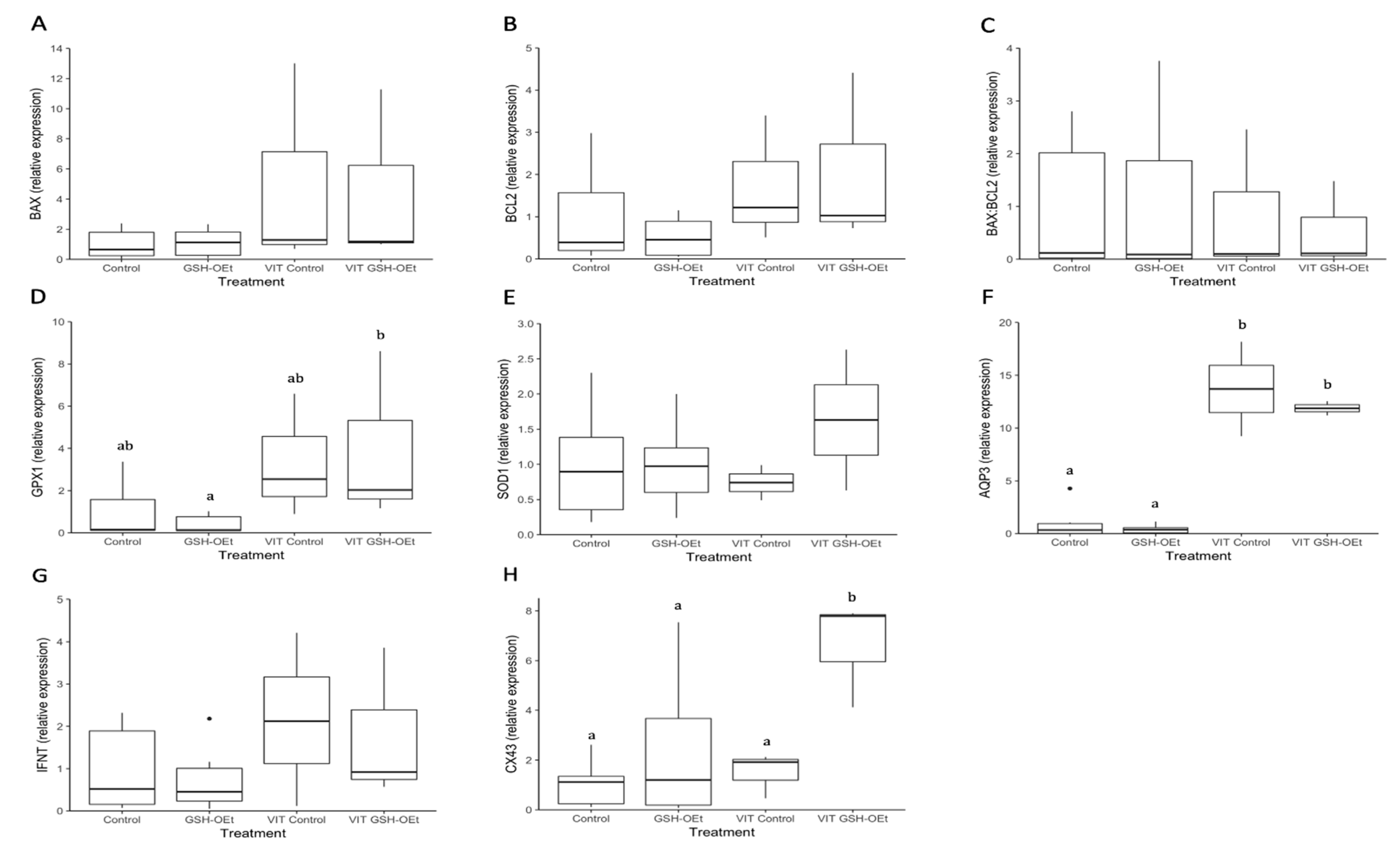
2.5. Developmental Competence and Gene Expression Determined in Embryos Derived from Vitrified/Warmed Bovine Oocytes In Vitro Matured in IVM Medium with or without GSH-OEt
3. Discussion
4. Materials and Methods
4.1. Chemicals and Suppliers
4.2. Oocyte Collection and In Vitro Maturation
4.3. Oocyte Vitrification and Warming
4.3.1. Vitrification Protocol
4.3.2. Warming Protocol
4.4. In Vitro Fertilization and Embryo Culture
4.5. Chromosome and Spindle Organization
4.6. Reactive Oxygen Species
4.7. Mitochondrial Activity and Distribution
4.8. RNA Extraction, Reverse Transcription and Quantitative Real-Time PCR Analysis
4.9. Experimental Design
4.9.1. Meiotic Spindle Status, ROS Production, Mitochondrial Activity and Distribution and Gene Expression Examined in Vitrified/Warmed Bovine Oocytes after Maturation in IVM Medium with or without GSH-OEt
4.9.2. Developmental Competence and Gene Expression Determined in Embryos Derived from Vitrified/Warmed Bovine Oocytes In Vitro Matured in IVM Medium with or without GSH-OEt
5. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- Mogas, T. Update on the vitrification of bovine oocytes and invitro-produced embryos. Reprod. Fertil. Dev. 2018, 31, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Vajta, G.; Kuwayama, M. Improving cryopreservation systems. Theriogenology 2006, 65, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Chian, R.C.; Kuwayama, M.; Tan, L.; Tan, J.; Kato, O.; Nagai, T. High survival rate of bovine oocytes matured in vitro following vitrification. J. Reprod. Dev. 2004, 50, 685–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vajta, G.; Holm, P.; Kuwayama, M.; Booth, P.J.; Jacobsen, H.; Greve, T.; Callesen, H. Open Pulled Straw (OPS) vitrification: A new way to reduce cryoinjuries of bovine ova and embryos. Mol. Reprod. Dev. 1998, 51, 53–58. [Google Scholar] [CrossRef]
- Vieira, A.D.; Mezzalira, A.; Barbieri, D.P.; Lehmkuhl, R.C.; Rubin, M.I.; Vajta, G. Calves born after open pulled straw vitrification of immature bovine oocytes. Cryobiology 2002, 45, 91–94. [Google Scholar] [CrossRef]
- Kono, T.; Kwon, O.Y.; Nakahara, T. Development of vitrified mouse oocytes after in vitro fertilization. Cryobiology 1991, 28, 50–54. [Google Scholar] [CrossRef]
- Cao, Y.X.; Xing, Q.; Li, L.; Cong, L.; Zhang, Z.G.; Wei, Z.L.; Zhou, P. Comparison of survival and embryonic development in human oocytes cryopreserved by slow-freezing and vitrification. Fertil. Steril. 2009, 92, 1306–1311. [Google Scholar] [CrossRef]
- Levi Setti, P.E.; Porcu, E.; Patrizio, P.; Vigiliano, V.; de Luca, R.; d’Aloja, P.; Spoletini, R.; Scaravelli, G. Human oocyte cryopreservation with slow freezing versus vitrification. Results from the National Italian Registry data, 2007–2011. Fertil. Steril. 2014, 102, 90–95.e2. [Google Scholar] [CrossRef]
- Hwang, I.S.; Hochi, S. Recent progress in cryopreservation of bovine oocytes. BioMed Res. Int. 2014, 2014, 570647. [Google Scholar] [CrossRef]
- Gupta, M.K.; Uhm, S.J.; Lee, H.T. Effect of vitrification and beta-mercaptoethanol on reactive oxygen species activity and in vitro development of oocytes vitrified before or after in vitro fertilization. Fertil. Steril. 2010, 93, 2602–2607. [Google Scholar] [CrossRef]
- Zhao, X.M.; Du, W.H.; Wang, D.; Hao, H.S.; Liu, Y.; Qin, T.; Zhu, H.B. Recovery of mitochondrial function and endogenous antioxidant systems in vitrified bovine oocytes during extended in vitro culture. Mol. Reprod. Dev. 2011, 78, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Dumollard, R.; Ward, Z.; Carroll, J.; Duchen, M.R. Regulation of redox metabolism in the mouse oocyte and embryo. Development 2007, 134, 455–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, W.J.; Banerjee, J.; Falcone, T.; Bena, J.; Agarwal, A.; Sharma, R.K. Oxidative stress and tumor necrosis factor-alpha-induced alterations in metaphase II mouse oocyte spindle structure. Fertil. Steril. 2007, 88 (Suppl. 4), 1220–1231. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Yuan, B.; Kwon, J.W.; Ahn, M.; Cui, X.S.; Bang, J.K.; Kim, N.H. Effect of antifreeze glycoprotein 8 supplementation during vitrification on the developmental competence of bovine oocytes. Theriogenology 2016, 86, 485–494 e1. [Google Scholar] [CrossRef] [PubMed]
- Sprícigo, J.; Morato, R.; Arcarons, N.; Yeste, M.; Dode, M.A.; López-Bejar, M.; Mogas, T. L-Carnitine and/or Resveratrol supplementation during the in vitro maturation of bovine oocytes prior to their vitrification and warming: Effects on spindle morphology, apoptosis, molecular profile and embryo development. Mol. Reprod. Dev. 2016. under revision. [Google Scholar]
- Suen, D.F.; Norris, K.L.; Youle, R.J. Mitochondrial dynamics and apoptosis. Genes Dev. 2008, 22, 1577–1590. [Google Scholar] [CrossRef] [Green Version]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Meister, A.; Anderson, M.E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef]
- Zuelke, K.A.; Jeffay, S.C.; Zucker, R.M.; Perreault, S.D. Glutathione (GSH) concentrations vary with the cell cycle in maturing hamster oocytes, zygotes, and pre-implantation stage embryos. Mol. Reprod. Dev. 2003, 64, 106–112. [Google Scholar] [CrossRef]
- Sutovsky, P.; Schatten, G. Depletion of glutathione during bovine oocyte maturation reversibly blocks the decondensation of the male pronucleus and pronuclear apposition during fertilization. Biol. Reprod. 1997, 56, 1503–1512. [Google Scholar] [CrossRef]
- de Matos, D.G.; Furnus, C.C. The importance of having high glutathione (GSH) level after bovine in vitro maturation on embryo development effect of beta-mercaptoethanol, cysteine and cystine. Theriogenology 2000, 53, 761–771. [Google Scholar] [CrossRef]
- Luberda, Z. The role of glutathione in mammalian gametes. Reprod. Biol. 2005, 5, 5–17. [Google Scholar] [PubMed]
- Abeydeera, L.R.; Wang, W.H.; Cantley, T.C.; Prather, R.S.; Day, B.N. Presence of beta-mercaptoethanol can increase the glutathione content of pig oocytes matured in vitro and the rate of blastocyst development after in vitro fertilization. Theriogenology 1998, 50, 747–756. [Google Scholar] [CrossRef]
- de Matos, D.G.; Furnus, C.C.; Moses, D.F.; Martinez, A.G.; Matkovic, M. Stimulation of glutathione synthesis of in vitro matured bovine oocytes and its effect on embryo development and freezability. Mol. Reprod. Dev. 1996, 45, 451–457. [Google Scholar] [CrossRef]
- de Matos, D.G.; Furnus, C.C.; Moses, D.F. Glutathione synthesis during in vitro maturation of bovine oocytes: Role of cumulus cells. Biol. Reprod. 1997, 57, 1420–1425. [Google Scholar] [CrossRef] [Green Version]
- Hara, H.; Yamane, I.; Noto, I.; Kagawa, N.; Kuwayama, M.; Hirabayashi, M.; Hochi, S. Microtubule assembly and in vitro development of bovine oocytes with increased intracellular glutathione level prior to vitrification and in vitro fertilization. Zygote 2014, 22, 476–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curnow, E.C.; Ryan, J.P.; Saunders, D.M.; Hayes, E.S. Developmental potential of bovine oocytes following IVM in the presence of glutathione ethyl ester. Reprod. Fertil. Dev. 2010, 22, 597–605. [Google Scholar] [CrossRef]
- Curnow, E.C.; Ryan, J.P.; Saunders, D.M.; Hayes, E.S. Primate model of metaphase I oocyte in vitro maturation and the effects of a novel glutathione donor on maturation, fertilization, and blastocyst development. Fertil. Steril. 2011, 95, 1235–1240. [Google Scholar] [CrossRef] [Green Version]
- Meister, A. Methods for the selective modification of glutathione metabolism and study of glutathione transport. Methods Enzym. 1985, 113, 571–585. [Google Scholar]
- Trapphoff, T.; Heiligentag, M.; Simon, J.; Staubach, N.; Seidel, T.; Otte, K.; Frohlich, T.; Arnold, G.J.; Eichenlaub-Ritter, U. Improved cryotolerance and developmental potential of in vitro and in vivo matured mouse oocytes by supplementing with a glutathione donor prior to vitrification. Mol. Hum. Reprod. 2016, 22, 867–881. [Google Scholar]
- Takahashi, M. Oxidative stress and redox regulation on in vitro development of mammalian embryos. J. Reprod. Dev. 2012, 58, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanfins, A.; Lee, G.Y.; Plancha, C.E.; Overstrom, E.W.; Albertini, D.F. Distinctions in meiotic spindle structure and assembly during in vitro and in vivo maturation of mouse oocytes. Biol. Reprod. 2003, 69, 2059–2067. [Google Scholar] [CrossRef] [PubMed]
- Schatten, G.; Simerly, C.; Schatten, H. Microtubule configurations during fertilization, mitosis, and early development in the mouse and the requirement for egg microtubule-mediated motility during mammalian fertilization. Proc. Natl. Acad. Sci. USA 1985, 82, 4152–4156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eroglu, A.; Toth, T.L.; Toner, M. Alterations of the cytoskeleton and polyploidy induced by cryopreservation of metaphase II mouse oocytes. Fertil. Steril. 1998, 69, 944–957. [Google Scholar] [CrossRef]
- Oliver, J.M.; Albertini, D.F.; Berlin, R.D. Effects of glutathione-oxidizing agents on microtubule assembly and microtubule-dependent surface properties of human neutrophils. J. Cell Biol. 1976, 71, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Zuelke, K.A.; Jones, D.P.; Perreault, S.D. Glutathione oxidation is associated with altered microtubule function and disrupted fertilization in mature hamster oocytes. Biol. Reprod. 1997, 57, 1413–1419. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Gu, R.; Lu, X.; Zhao, S.; Feng, Y.; Sun, Y. Preincubation with glutathione ethyl ester improves the developmental competence of vitrified mouse oocytes. J. Assist. Reprod. Genet. 2018, 35, 1169–1178. [Google Scholar] [CrossRef]
- Morato, R.; Izquierdo, D.; Paramio, M.T.; Mogas, T. Cryotops versus open-pulled straws (OPS) as carriers for the cryopreservation of bovine oocytes: Effects on spindle and chromosome configuration and embryo development. Cryobiology 2008, 57, 137–141. [Google Scholar] [CrossRef]
- Succu, S.; Gadau, S.D.; Serra, E.; Zinellu, A.; Carru, C.; Porcu, C.; Naitana, S.; Berlinguer, F.; Leoni, G.G. A recovery time after warming restores mitochondrial function and improves developmental competence of vitrified ovine oocytes. Theriogenology 2018, 110, 18–26. [Google Scholar] [CrossRef]
- Somfai, T.; Ozawa, M.; Noguchi, J.; Kaneko, H.; Kuriani Karja, N.W.; Farhudin, M.; Dinnyes, A.; Nagai, T.; Kikuchi, K. Developmental competence of in vitro-fertilized porcine oocytes after in vitro maturation and solid surface vitrification: Effect of cryopreservation on oocyte antioxidative system and cell cycle stage. Cryobiology 2007, 55, 115–126. [Google Scholar] [CrossRef]
- Dadarwal, D.; Adams, G.P.; Hyttel, P.; Brogliatti, G.M.; Caldwell, S.; Singh, J. Organelle reorganization in bovine oocytes during dominant follicle growth and regression. Reprod Biol Endocrinol 2015, 13, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, Y. Updating the markers for oocyte quality evaluation: Intracellular temperature as a new index. Reprod Med. Biol. 2018, 17, 434–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, S.; Mabuchi, T.; Hirata, S.; Shoda, T.; Kasai, T.; Yokota, S.; Shitara, H.; Yonekawa, H.; Hoshi, K. Correlation of abnormal mitochondrial distribution in mouse oocytes with reduced developmental competence. Tohoku J. Exp. Med. 2006, 210, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Brevini, T.A.; Cillo, F.; Antonini, S.; Gandolfi, F. Cytoplasmic remodelling and the acquisition of developmental competence in pig oocytes. Anim. Reprod. Sci. 2007, 98, 23–38. [Google Scholar] [CrossRef]
- Dai, J.; Wu, C.; Muneri, C.W.; Niu, Y.; Zhang, S.; Rui, R.; Zhang, D. Changes in mitochondrial function in porcine vitrified MII-stage oocytes and their impacts on apoptosis and developmental ability. Cryobiology 2015, 71, 291–298. [Google Scholar] [CrossRef]
- Morato, R.; Mogas, T.; Maddox-Hyttel, P. Ultrastructure of bovine oocytes exposed to Taxol prior to OPS vitrification. Mol. Reprod. Dev. 2008, 75, 1318–1326. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Martin, M.; Bonet, S.; Morato, R.; Yeste, M. Supplementing culture and vitrification-warming media with l-ascorbic acid enhances survival rates and redox status of IVP porcine blastocysts via induction of GPX1 and SOD1 expression. Cryobiology 2014, 68, 451–458. [Google Scholar] [CrossRef]
- Favetta, L.A.; St John, E.J.; King, W.A.; Betts, D.H. High levels of p66shc and intracellular ROS in permanently arrested early embryos. Free Radic. Biol. Med. 2007, 42, 1201–1210. [Google Scholar] [CrossRef]
- Yoneda, A.; Suzuki, K.; Mori, T.; Ueda, J.; Watanabe, T. Effects of delipidation and oxygen concentration on in vitro development of porcine embryos. J. Reprod. Dev. 2004, 50, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Arcarons, N.; Vendrell-Flotats, M.; Yeste, M.; Mercade, E.; Lopez-Bejar, M.; Mogas, T. Cryoprotectant role of exopolysaccharide of Pseudomonas sp. ID1 in the vitrification of IVM cow oocytes. Reprod. Fertil. Dev. 2019, 31, 1507–1519. [Google Scholar] [CrossRef]
- Barros, F.D.A.; Adona, P.R.; Guemra, S.; Damiao, B.C.M. Oxidative homeostasis in oocyte competence for in vitro embryo development. Anim. Sci. J. 2019, 90, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Albarracin, J.L.; Morato, R.; Izquierdo, D.; Mogas, T. Vitrification of calf oocytes: Effects of maturation stage and prematuration treatment on the nuclear and cytoskeletal components of oocytes and their subsequent development. Mol. Reprod. Dev. 2005, 72, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Spricigo, J.F.; Morato, R.; Arcarons, N.; Yeste, M.; Dode, M.A.; Lopez-Bejar, M.; Mogas, T. Assessment of the effect of adding L-carnitine and/or resveratrol to maturation medium before vitrification on in vitro-matured calf oocytes. Theriogenology 2017, 89, 47–57. [Google Scholar] [CrossRef]
- Khatir, H.; Carolan, C.; Lonergan, P.; Mermillod, P. Characterization of calf follicular fluid and its ability to support cytoplasmic maturation of cow and calf oocytes. J. Reprod. Fertil. 1997, 111, 267–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renault, T.T.; Dejean, L.M.; Manon, S. A brewing understanding of the regulation of Bax function by Bcl-xL and Bcl-2. Mech. Ageing Dev. 2017, 161 (Pt B), 201–210. [Google Scholar] [CrossRef]
- Ebrahimi, B.; Valojerdi, M.R.; Eftekhari-Yazdi, P.; Baharvand, H. In vitro maturation, apoptotic gene expression and incidence of numerical chromosomal abnormalities following cryotop vitrification of sheep cumulus-oocyte complexes. J. Assist. Reprod Genet. 2010, 27, 239–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhali, A.; Anchamparuthy, V.M.; Butler, S.P.; Pearson, R.E.; Mullarky, I.K.; Gwazdauskas, F.C. Effect of droplet vitrification on development competence, actin cytoskeletal integrity and gene expression in in vitro cultured mouse embryos. Theriogenology 2009, 71, 1408–1416. [Google Scholar] [CrossRef]
- Kuzmany, A.; Havlicek, V.; Wrenzycki, C.; Wilkening, S.; Brem, G.; Besenfelder, U. Expression of mRNA, before and after freezing, in bovine blastocysts cultured under different conditions. Theriogenology 2011, 75, 482–494. [Google Scholar] [CrossRef]
- Madrid Gaviria, S.; Lopez Herrera, A.; Urrego, R.; Restrepo Betancur, G.; Echeverri Zuluaga, J.J. Effect of resveratrol on vitrified in vitro produced bovine embryos: Recovering the initial quality. Cryobiology 2019, 89, 42–50. [Google Scholar] [CrossRef]
- Pereira, B.A.; Zangeronimo, M.G.; Castillo-Martin, M.; Gadani, B.; Chaves, B.R.; Rodriguez-Gil, J.E.; Bonet, S.; Yeste, M. Supplementing Maturation Medium With Insulin Growth Factor I and Vitrification-Warming Solutions With Reduced Glutathione Enhances Survival Rates and Development Ability of in vitro Matured Vitrified-Warmed Pig Oocytes. Front. Physiol. 2018, 9, 1894. [Google Scholar] [CrossRef] [Green Version]
- Turathum, B.; Saikhun, K.; Sangsuwan, P.; Kitiyanant, Y. Effects of vitrification on nuclear maturation, ultrastructural changes and gene expression of canine oocytes. Reprod. Biol. Endocrinol. RbE 2010, 8, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Harding, J.J. Glycation-induced inactivation and loss of antigenicity of catalase and superoxide dismutase. Biochem. J. 1997, 328 (Pt 2), 599–605. [Google Scholar] [CrossRef] [Green Version]
- Cebrian-Serrano, A.; Salvador, I.; Garcia-Rosello, E.; Pericuesta, E.; Perez-Cerezales, S.; Gutierrez-Adan, A.; Coy, P.; Silvestre, M.A. Effect of the bovine oviductal fluid on in vitro fertilization, development and gene expression of in vitro-produced bovine blastocysts. Reprod. Domest. Anim. Zuchthyg. 2013, 48, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Edashige, K. The movement of water and cryoprotectants across the plasma membrane of mammalian oocytes and embryos and its relevance to vitrification. J. Reprod. Dev. 2016, 62, 317–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jablonski, E.M.; Mattocks, M.A.; Sokolov, E.; Koniaris, L.G.; Hughes, F.M., Jr.; Fausto, N.; Pierce, R.H.; McKillop, I.H. Decreased aquaporin expression leads to increased resistance to apoptosis in hepatocellular carcinoma. Cancer Lett. 2007, 250, 36–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, N.; Wan, P.C.; Hao, Z.D.; Gao, F.F.; Yang, L.; Cui, M.S.; Wu, Y.; Liu, J.H.; Liu, S.; Chen, H.; et al. Expression of interferon-tau mRNA in bovine embryos derived from different procedures. Reprod. Domest. Anim. Zuchthyg. 2009, 44, 132–139. [Google Scholar] [CrossRef]
- Zhao, X.M.; Min, J.T.; Du, W.H.; Hao, H.S.; Liu, Y.; Qin, T.; Wang, D.; Zhu, H.B. Melatonin enhances the in vitro maturation and developmental potential of bovine oocytes denuded of the cumulus oophorus. Zygote 2015, 23, 525–536. [Google Scholar] [CrossRef]
- Rizos, D.; Gutierrez-Adan, A.; Perez-Garnelo, S.; De La Fuente, J.; Boland, M.P.; Lonergan, P. Bovine embryo culture in the presence or absence of serum: Implications for blastocyst development, cryotolerance, and messenger RNA expression. Biol. Reprod. 2003, 68, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Adan, A.; Rizos, D.; Fair, T.; Moreira, P.N.; Pintado, B.; de la Fuente, J.; Boland, M.P.; Lonergan, P. Effect of speed of development on mRNA expression pattern in early bovine embryos cultured in vivo or in vitro. Mol. Reprod. Dev. 2004, 68, 441–448. [Google Scholar] [CrossRef]
- Rizos, D.; Ward, F.; Boland, M.P.; Lonergan, P. Effect of culture system on the yield and quality of bovine blastocysts as assessed by survival after vitrification. Theriogenology 2001, 56, 1–16. [Google Scholar] [CrossRef]
- Arcarons, N.; Morato, R.; Spricigo, J.F.; Ferraz, M.A.; Mogas, T. Spindle configuration and developmental competence of in vitro-matured bovine oocytes exposed to NaCl or sucrose prior to Cryotop vitrification. Reprod. Fertil. Dev. 2015, 28, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Martin, M.; Bonet, S.; Morato, R.; Yeste, M. Comparative effects of adding beta-mercaptoethanol or L-ascorbic acid to culture or vitrification-warming media on IVF porcine embryos. Reprod. Fertil. Dev. 2014, 26, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Frigoni, N.A.; Leao, B.C.; Dall’Acqua, P.C.; Mingoti, G.Z. Improving the cytoplasmic maturation of bovine oocytes matured in vitro with intracellular and/or extracellular antioxidants is not associated with increased rates of embryo development. Theriogenology 2016, 86, 1897–1905. [Google Scholar] [CrossRef] [Green Version]
- Esposti, M.D.; Hatzinisiriou, I.; McLennan, H.; Ralph, S. Bcl-2 and mitochondrial oxygen radicals. New approaches with reactive oxygen species-sensitive probes. J. Biol. Chem. 1999, 274, 29831–29837. [Google Scholar] [CrossRef] [Green Version]
- Moawad, A.R.; Xu, B.; Tan, S.L.; Taketo, T. l-carnitine supplementation during vitrification of mouse germinal vesicle stage-oocytes and their subsequent in vitro maturation improves meiotic spindle configuration and mitochondrial distribution in metaphase II oocytes. Hum. Reprod 2014, 29, 2256–2268. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | MII (%) | Normal | Microtubule Configuration (%) * | Chromosome Distribution (%) * | |||
|---|---|---|---|---|---|---|---|
| Abnormal | Absent | Abnormal | Absent | ||||
| Control | 127 | 79.4 ± 9.2 | 74.1 ± 4.4 | 16.2 ± 5.6 a | 9.7 ± 1.1 | 25.9 ± 8.3 | 0 |
| GSH-OEt | 113 | 76.5 ± 6.5 | 65.6 ± 5.7 | 26.2 ± 9.0 ab | 8.1 ± 4.3 | 33.2 ± 10.6 | 1.1 ± 1.1 |
| VIT Control | 71 | 60.0 ± 5.6 | 54.8 ± 7.8 | 32.7 ± 10.8 b | 12.4 ± 1.4 | 45.2 ± 14.5 | 0 |
| VIT GSH-OEt | 53 | 57.4 ± 10.7 | 58.5 ± 8.8 | 31.1 ± 9.3 ab | 10.4 ± 5.8 | 41.5 ± 14 | 0 |
| n | Cleavage Rate 48 hpi | 16-Cell Embryo 96 hpi | Blastocyst Yields | D8 Blastocysts | |||||
|---|---|---|---|---|---|---|---|---|---|
| D7 Blastocyst | D8 Blastocyst | nD8 | Non-Expanded | Expanded | Hatched | ||||
| Control | 381 | 73.09 ± 3.84 a | 53.74 ± 8.80 a | 14.74 ± 1.71 ab | 23.12 ± 5.17 a | 85 | 48.04 ± 14.66 | 24.85 ± 2.76 | 27.11 ± 14.70 a |
| GSH-OEt | 307 | 74.76 ± 3.26 a | 45.62 ± 2.95 a | 20.18 ± 6.45 a | 28.59 ± 8.06 a | 84 | 35.86 ± 9.95 | 41.48 ± 5.48 | 22.66 ± 5.40 a |
| VIT Control | 136 | 42.84 ± 6.88 b | 25.48 ± 4.10 b | 4.09 ± 1.30 b | 5.26 ± 0.58 b | 7 | 44.44 ± 29.40 | 55.56 ± 29.40 | 0 b |
| VIT GSH-OEt | 134 | 45.50 ± 10.16 b | 37.54 ± 2.44 ab | 6.62 ± 0.81 b | 13.78 ± 3.20 ab | 20 | 50.43 ± 12.75 | 40.04 ± 6.71 | 9.52 ± 9.52 ab |
| Symbol | GenBank Accession Number | Primer Sequence (5′–3′) | Fragment Size (bp) |
|---|---|---|---|
| BAX | NM_173894.1 | F: ACCAAGAAGCTGAGCGAGTG | 116 |
| R: CGGAAAAAGACCTCTCGGGG | |||
| BCL2 | NM_001166486.1 | F: GAGTTCGGAGGGGTCATGTG | 211 |
| R: TGAGCAGTGCCTTCAGAGAC | |||
| GPX1 | NM_174076.3 | F: CTGAAGTACGTCCGACCAGG | 153 |
| R: GTCGGTCATGAGAGCAGTGG | |||
| SOD1 | NM_174615.2 | F: ACACAAGGCTGTACCAGTGC | 102 |
| R: CACATTGCCCAGGTCTCCAA | |||
| AQP3 | NM_001079794.1 | F: GTGGACCCCTACAACAACCC | 222 |
| R: CAGGAGCGGAGAGACAATGG | |||
| IFN-τ | AF238612 | F: CTGAAGGTTCACCCAGACCC | 197 |
| R: GAGTCTGTTCATTCGGGCCA | |||
| CX43 | NM_174068.2 | F: TGGAATGCAAGAGAGGTTGAAAGAGG | 294 |
| R: AACACTCTCCAGAACACATGATCG | |||
| PPIA | NM_178320.2 | F: CATACAGGTCCTGGCATCTTGTCC | 108 |
| R: CACGTGCTTGCCATCCAACC | |||
| H3F3A | NM_001014389.2 | F: CATGGCTCGTACAAAGCAGA | 136 |
| R: ACCAGGCCTGTAACGATGAG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Martínez, T.; Vendrell-Flotats, M.; Martínez-Rodero, I.; Ordóñez-León, E.A.; Álvarez-Rodríguez, M.; López-Béjar, M.; Yeste, M.; Mogas, T. Glutathione Ethyl Ester Protects In Vitro-Maturing Bovine Oocytes against Oxidative Stress Induced by Subsequent Vitrification/Warming. Int. J. Mol. Sci. 2020, 21, 7547. https://doi.org/10.3390/ijms21207547
García-Martínez T, Vendrell-Flotats M, Martínez-Rodero I, Ordóñez-León EA, Álvarez-Rodríguez M, López-Béjar M, Yeste M, Mogas T. Glutathione Ethyl Ester Protects In Vitro-Maturing Bovine Oocytes against Oxidative Stress Induced by Subsequent Vitrification/Warming. International Journal of Molecular Sciences. 2020; 21(20):7547. https://doi.org/10.3390/ijms21207547
Chicago/Turabian StyleGarcía-Martínez, Tania, Meritxell Vendrell-Flotats, Iris Martínez-Rodero, Erika Alina Ordóñez-León, Manuel Álvarez-Rodríguez, Manel López-Béjar, Marc Yeste, and Teresa Mogas. 2020. "Glutathione Ethyl Ester Protects In Vitro-Maturing Bovine Oocytes against Oxidative Stress Induced by Subsequent Vitrification/Warming" International Journal of Molecular Sciences 21, no. 20: 7547. https://doi.org/10.3390/ijms21207547
APA StyleGarcía-Martínez, T., Vendrell-Flotats, M., Martínez-Rodero, I., Ordóñez-León, E. A., Álvarez-Rodríguez, M., López-Béjar, M., Yeste, M., & Mogas, T. (2020). Glutathione Ethyl Ester Protects In Vitro-Maturing Bovine Oocytes against Oxidative Stress Induced by Subsequent Vitrification/Warming. International Journal of Molecular Sciences, 21(20), 7547. https://doi.org/10.3390/ijms21207547