A Brief History of Adherons: The Discovery of Brain Exosomes
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Cell Adhesion Is Dependent Upon High Molecular Weight Protein Complexes
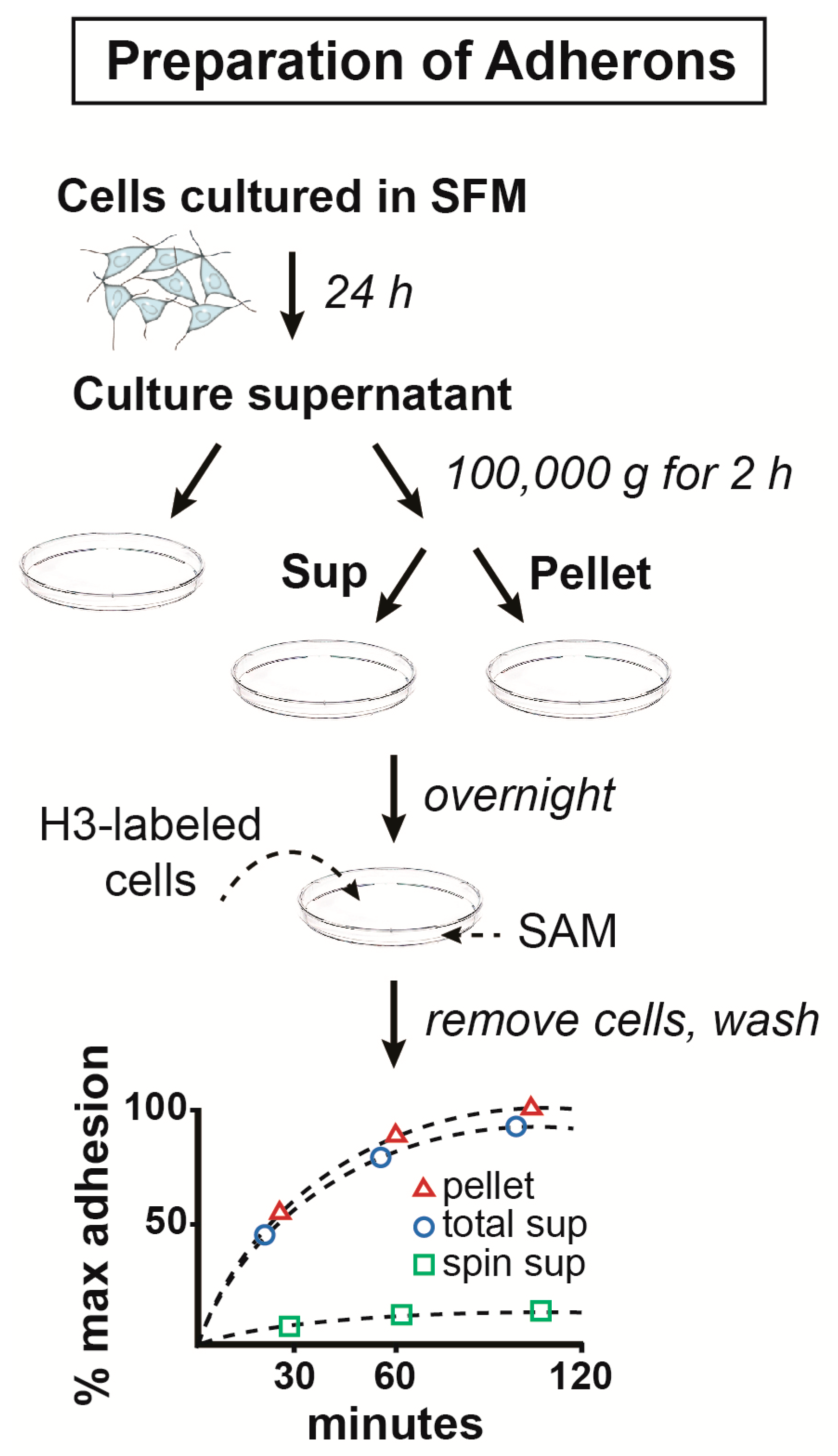
2.1. The Structure and Composition of Adherons
2.2. Isolation and Cloning of a Neurotrophic Factor from Adherons
2.3. Adherons vs. Exosomes
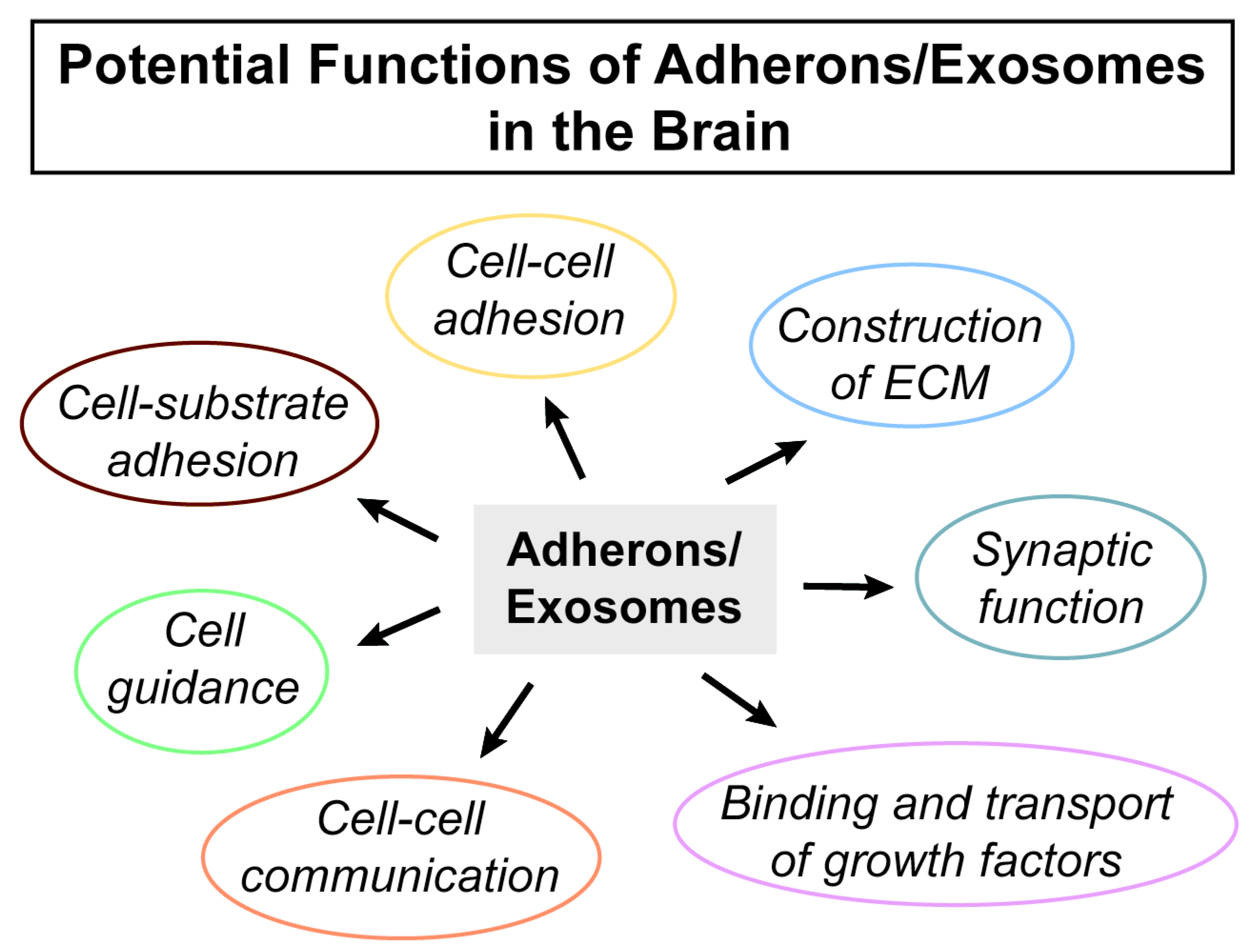
2.4. Exosomes, Adhesion, and the Brain ECM
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BDNF | Brain-derived neurotrophic factor |
| CNS | Central nervous system |
| ECM | Extracellular matrix |
| FGF | Fibroblast growth factor |
| GAG | Glycosaminoglycan |
| GDGF | Glial-derived growth factor |
| HSPG | Heparin sulfate proteoglycan |
| NGF | Nerve growth factor |
| PNN | Perineuronal net |
| SAM | Substrate-attached material |
| TR | Transferrin receptor |
References
- Bargmann, C.I.; Hung, M.C.; Weinberg, R.A. The neu oncogene encodes an epidermal growth factor receptor-related protein. Nature 1986, 319, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Engele, J.; Schubert, D.; Bohn, M.C. Conditioned media derived from glial cell lines promote survival and differentiation of dopaminergic neurons in vitro: Role of mesencephalic glia. J. Neurosci. Res. 1991, 30, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Schubert, D.; Klar, A.; Park, M.; Dargusch, R.; Fischer, W.H. F-spondin promotes nerve precursor differentiation. J. Neurochem. 2006, 96, 444–453. [Google Scholar] [CrossRef]
- Kidokoro, Y.; Heinemann, S.; Schubert, D.; Brandt, B.L.; Klier, F.G. Synapse formation and neurotrophic effects on muscle cell lines. Cold Spring Harb. Symp. Quant. Biol. 1976, 40, 373–388. [Google Scholar] [CrossRef] [PubMed]
- Schubert, D.; Heinemann, S.; Carlisle, W.; Tarikas, H.; Kimes, B.; Patrick, J.; Steinbach, J.H.; Culp, W.; Brandt, B.L. Clonal cell lines from the rat central nervous system. Nature 1974, 249, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Bauer, R.; Bergmann, R.; Beyer, G.J.; Manfrass, P.; Steinbach, J.; Kretzschmar, M.; Enghardt, W.; Hoyer, D.; Gunther, K.; Schubert, H.; et al. Investigations of cerebral glucose utilization into the newborn brain: A [18F]-FDG positron emission tomography study using a high resolution multiwire proportional chamber detector device. Exp. Pathol. 1991, 42, 229–233. [Google Scholar] [CrossRef]
- Schubert, D. The substrate attached material synthesized by clonal cell lines of nerve, glia, and muscle. Brain Res. 1977, 132, 337–346. [Google Scholar] [CrossRef]
- Schubert, D. NGF-induced alterations in protein secretion and substrate-attached material of a clonal nerve cell line. Brain Res. 1978, 155, 196–200. [Google Scholar] [CrossRef]
- Schubert, D.; LaCorbiere, M. Role of a 16S glycoprotein complex in cellular adhesion. Proc. Natl. Acad. Sci. USA 1980, 77, 4137–4141. [Google Scholar] [CrossRef] [Green Version]
- Schubert, D.; LaCorbiere, M. A role of seceted glycosaminoglycans in cell-substratum adhesion. J. Biol. Chem. 1980, 255, 11564–11569. [Google Scholar]
- Schubert, D.; LaCorbiere, M.; Klier, F.G.; Birdwell, C. The structure and function of myoblast adherons. Cold Spring Harb. Symp. Quant. Biol. 1983, 48, 539–549. [Google Scholar] [CrossRef]
- Patel, G.K.; Khan, M.A.; Zubair, H.; Srivastava, S.K.; Khushman, M.; Singh, S.; Singh, A.P. Comparative analysis of exosome isolation methods using culture supernatant for optimum yield, purity and downstream applications. Sci. Rep. 2019, 9, 5335. [Google Scholar] [CrossRef] [Green Version]
- Pan, B.; Johnstone, R. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: Selective externalization of the receptor. Cell 1983, 33, 967–978. [Google Scholar] [CrossRef]
- Schubert, D.; LaCorbiere, M.; Klier, F.G.; Birdwell, C. A role for adherons in neural retina cell adhesion. J. Cell Biol. 1983, 96, 990–998. [Google Scholar] [CrossRef] [PubMed]
- Purushothaman, A.; Bandari, S.K.; Liu, J.; Mobley, J.A.; Brown, E.E.; Sanderson, R.D. Fibronectin on the Surface of Myeloma Cell-derived Exosomes Mediates Exosome-Cell Interactions. J. Biol. Chem. 2016, 291, 1652–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, D.; LaCorbiere, M. The specificity of extracellular glycoprotein complexes in mediating cellular adhesion. J. Neurosci. 1982, 2, 82–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilien, J.; Moscona, A.A. Cell aggregation: Its enhancement by a supernatant from culutres of homologous cells. Science 1967, 157, 70–72. [Google Scholar] [CrossRef] [PubMed]
- Cole, G.J.; Schubert, D.; Glaser, L. Cell-substratum adhesion in chick neural retina depends upon protein-heparan sulfate interactions. J. Cell Biol. 1985, 100, 1192–1199. [Google Scholar] [CrossRef] [Green Version]
- Christianson, H.C.; Belting, M. Heparan sulfate proteoglycan as a cell-surface endocytosis receptor. Matrix Biol. 2014, 35, 51–55. [Google Scholar] [CrossRef]
- Cole, G.; Glaser, L. A heparin-binding domain from N-CAM is involved in neural cell-substratum adhesion. J. Cell Biol. 1986, 102, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Schubert, D.; LaCorbiere, M. Isolation of an adhesion-mediating protein from chick neural retina adherons. J. Cell Biol. 1985, 101, 1071–1077. [Google Scholar] [CrossRef] [Green Version]
- Schubert, D.; LaCorbiere, M.; Esch, F. A chick neural retina adhesion and survival molecule is a retinal-binding protein. J. Cell Biol. 1986, 102, 2295–2301. [Google Scholar] [CrossRef] [Green Version]
- Berman, P.; Gray, P.; Chen, E.; Keyser, K.; Ehrlich, D.; Karten, H.; LaCorbiere, M.; Esch, F.; Schubert, D. Sequence analysis, cellular localization and expression of a neuroretina adhesion and cell survival molecule. Cell 1987, 51, 134–142. [Google Scholar] [CrossRef]
- Matsukawa, T.; Sugitani, K.; Mawatari, K.; Koriyama, Y.; Liu, Z.; Tanaka, M.; Kato, S. Role of purpurin as a retinol-binding protein in goldfish retina during the early stage of optic nerve regeneration: Its priming action on neurite outgrowth. J. Neurosci. 2004, 24, 8346–8353. [Google Scholar] [CrossRef]
- Nagashima, M.; Sakurai, H.; Mawatari, K.; Koriyama, Y.; Matsukawa, T.; Kato, S. Involvement of retinoic acid signaling in goldfish optic nerve regeneration. Neurochem. Int. 2009, 54, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Murayama, D.; Nagashima, M.; Higashi, T.; Mawatari, K.; Matsukawa, T.; Kato, S. Purpurin expression in the zebrafish retina during early development and after optic nerve lesion in adults. Brain Res. 2007, 1153, 34–42. [Google Scholar] [CrossRef]
- Nagashima, M.; Saito, J.; Mawatari, K.; Mori, Y.; Matsukawa, T.; Koriyama, Y.; Kato, S. A hypoplastic retinal lamination in the purpurin knock down embryo in zebrafish. Adv. Exp. Med. Biol. 2010, 664, 517–524. [Google Scholar]
- Kato, S.; Matsukawa, T.; Koriyama, Y.; Sugitani, K.; Ogai, K. A molecular mechanism of optic nerve regeneration in fish: The retinoid signaling pathway. Prog. Retin. Eye Res. 2013, 37, 13–30. [Google Scholar] [CrossRef]
- Park, H.; Shim, J.S.; Kim, B.S.; Jung, H.J.; Huh, T.L.; Kwon, H.J. Purpurin inhibits adipocyte-derived leucine aminopeptidase and angiogenesis in a zebrafish model. Biochem. Biophys. Res. Commun. 2014, 450, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.B.; Gao, H.W.; Peng, C. Quinones as preventive agents in Alzheimer’s diseases: Focus on NLRP3 inflammasomes. J. Pharm. Pharmacol. 2020, 72, 1481–1490. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.; Heuser, J.; Stahl, P. Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J. Cell Biol. 1983, 97, 329–339. [Google Scholar] [CrossRef] [PubMed]
- van der Pol, E.; Boing, A.; Harrison, P.; Sturk, A.; Nieuwland, R. Classification, functions, and clinical relevance of extracellular vesicles. Pharmacol. Rev. 2012, 64, 676–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mincheva-Nilsson, L.; Baranov, V.; Nagaeva, O.; Dehlin, E. Isolation and Characterization of Exosomes from Cultures of Tissue Explants and Cell Lines. Curr. Protoc. Immunol. 2016, 115, 14.42.1–14.42.21. [Google Scholar] [CrossRef]
- Krishnaswamy, V.R.; Benbenishty, A.; Blinder, P.; Sagi, I. Demystifying the extracellular matrix and its proteolytic remodeling in the brain: Structural and functional insights. Cell. Mol. Life Sci. 2019, 76, 3229–3248. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Balzac, C.A.; Lazaro-Pena, M.; Tecle, E.; Gomez, N.; Bulow, H.E. Complex cooperative functions of heparan sulfate proteoglycans shape nervous system development in Caenorhabditis elegans. G3 Genes Genomes Genet. 2014, 4, 1859–1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, R.; Nakamura, F.; Fukunaga, S. Diverse function of perlecan in central nervous system cells in vitro. Anim. Sci. J. 2015, 86, 904–911. [Google Scholar] [CrossRef] [PubMed]
- Buzas, E.I.; Toth, E.A.; Sodar, B.W.; Szabo-Taylor, K.E. Molecular interactions at the surface of extracellular vesicles. Sem. Immunopathol. 2018, 40, 453–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saint-Pol, J.; Gosselet, F.; Duban-Deweer, S.; Pottiez, G.; Karamanos, Y. Targeting and crossing the blood-brain barrier with extracellular vesicles. Cells 2020, 9, 851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, D. The possible role of adhesion in synaptic modification. Trends Neurosci. 1991, 14, 127–130. [Google Scholar] [CrossRef]
- Orlandi, C.; Omori, Y.; Wang, Y.; Cao, Y.; Ueno, A.; Roux, M.J.; Condomitti, G.; de Wit, J.; Kanagawa, M.; Furukawa, T.; et al. Transsynaptic binding of orphan receptor GPR179 to dystroglycan-pikachurin complex is essential for the synaptic organization of photoreceptors. Cell Rep. 2018, 25, 130–145. [Google Scholar] [CrossRef] [Green Version]
- Schubert, D.; Schroeder, R.; LaCorbiere, M.; Saitoh, T.; Cole, G. Amyloid beta protein precursor is possibly a heparan sulfate proteoglycan core protein. Science 1988, 241, 223–226. [Google Scholar] [CrossRef]
- Schubert, D.; LaCorbiere, M.; Saitoh, T.; Cole, G. Characterization of an amyloid beta precursor protein that binds heparin and contains tyrosine sulfate. Proc. Natl. Acad. Sci. USA 1989, 86, 2066–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, D.; Jin, L.-W.; Saitoh, T.; Cole, G. The regulation of amyloid beta protein precursor secretion and its modulatory role in cell adhesion. Neuron 1989, 3, 689–694. [Google Scholar] [CrossRef]
- Klier, F.G.; Cole, G.; Stallcup, W.; Schubert, D. Amyloid beta protein precursor is associated with extracellular matrix. Brain Res. 1990, 515, 336–342. [Google Scholar] [CrossRef]
- Miranda, A.M.; Lasiecka, Z.M.; Xu, Y.; Neufeld, J.; Shahriar, S.; Simoes, S.; Chan, R.B.; Oliveira, T.G.; Small, S.A.; Di Paolo, G. Neuronal lysosomal dysfunction releases exosomes harboring APP C-terminal fragments and unique lipid signatures. Nat. Commun. 2018, 9, 291. [Google Scholar] [CrossRef] [Green Version]
- Bespalov, M.M.; Sidorova, Y.A.; Tumova, S.; Ahonen-Bishopp, A.; Magalhaes, A.C.; Kulesskiy, E.; Paveliev, M.; Rivera, C.; Rauvala, H.; Saarma, M. Heparan sulfate proteoglycan syndecan-3 is a novel receptor for GDNF, neurturin and artemin. J. Cell Biol. 2011, 192, 153–169. [Google Scholar] [CrossRef] [Green Version]
- Saengsawang, W.; Kongoun, S.; Chanda, M. Exercise increases brain-derived neurotrophic factor in serum exosomes. FASEB J. 2017, 31, 839–845. [Google Scholar]
- Goetzi, E.J.; Nogueras-Ortiz, C.; Mustapic, M.; Mullins, R.J.; Abner, E.L.; Schwartz, J.B.; Kapogiannis, D. Deficient neurotrophic factors of CSPG4-type neural cell exosomes in Alzheimer disease. FASEB J. 2019, 33, 231–238. [Google Scholar] [CrossRef]
- Hormung, S.; Dutta, S.; Bitan, G. CNS-derived blood exosomes as a promising source of biomarkers: Opportunities and challenges. Front. Mol. Neurosci. 2020, 13, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; LeBleu, V.S. The biology, function and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schubert, D. A Brief History of Adherons: The Discovery of Brain Exosomes. Int. J. Mol. Sci. 2020, 21, 7673. https://doi.org/10.3390/ijms21207673
Schubert D. A Brief History of Adherons: The Discovery of Brain Exosomes. International Journal of Molecular Sciences. 2020; 21(20):7673. https://doi.org/10.3390/ijms21207673
Chicago/Turabian StyleSchubert, David. 2020. "A Brief History of Adherons: The Discovery of Brain Exosomes" International Journal of Molecular Sciences 21, no. 20: 7673. https://doi.org/10.3390/ijms21207673
APA StyleSchubert, D. (2020). A Brief History of Adherons: The Discovery of Brain Exosomes. International Journal of Molecular Sciences, 21(20), 7673. https://doi.org/10.3390/ijms21207673