Identification, Characterization and Synthesis of Walterospermin, a Sperm Motility Activator from the Egyptian Black Snake Walterinnesia aegyptia Venom

 ,
, 
 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Results
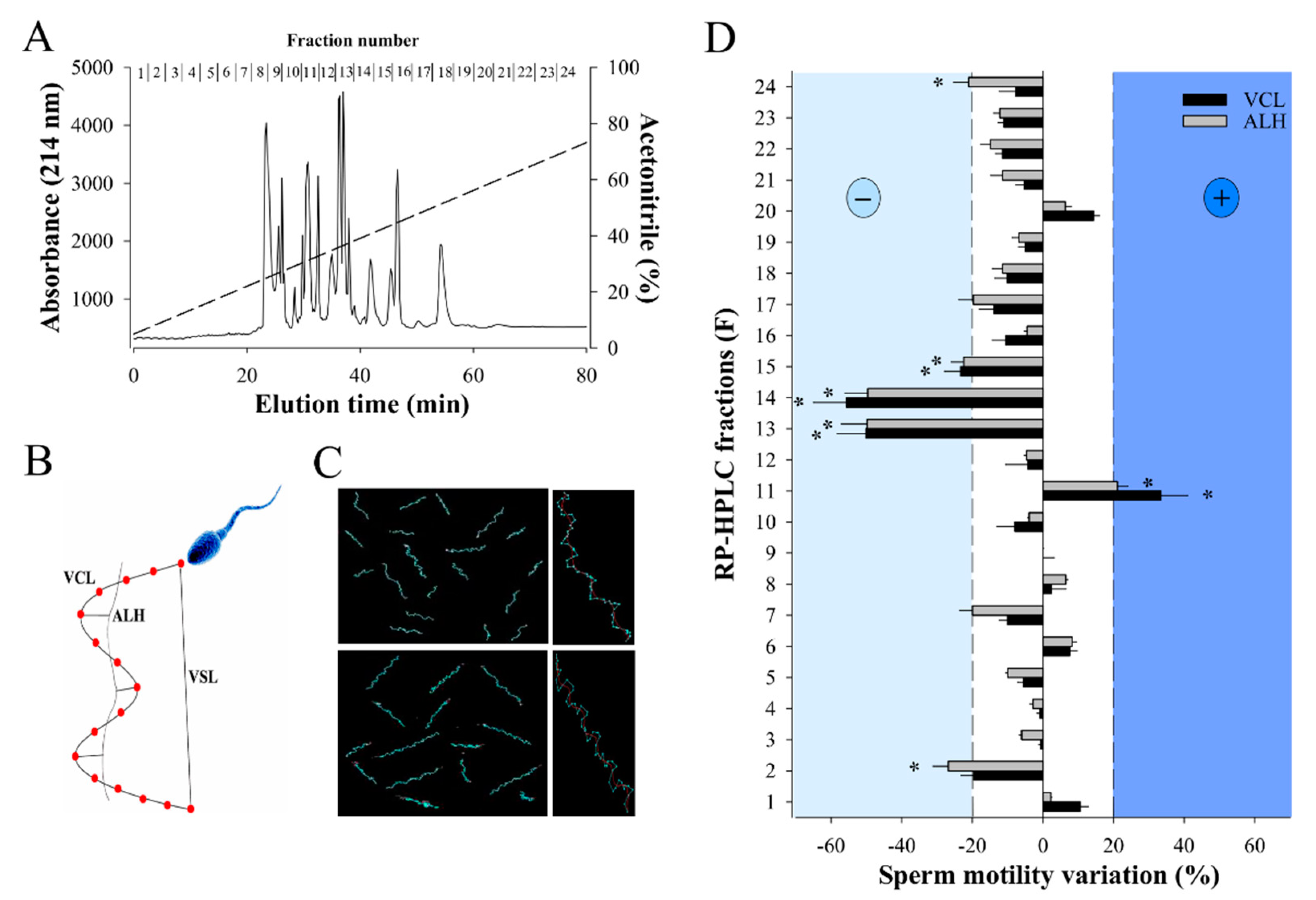
2.1. RP-HPLC Separation of W. aegyptia Venom Yields a Fraction that Positively Stimulates Sperm Motility
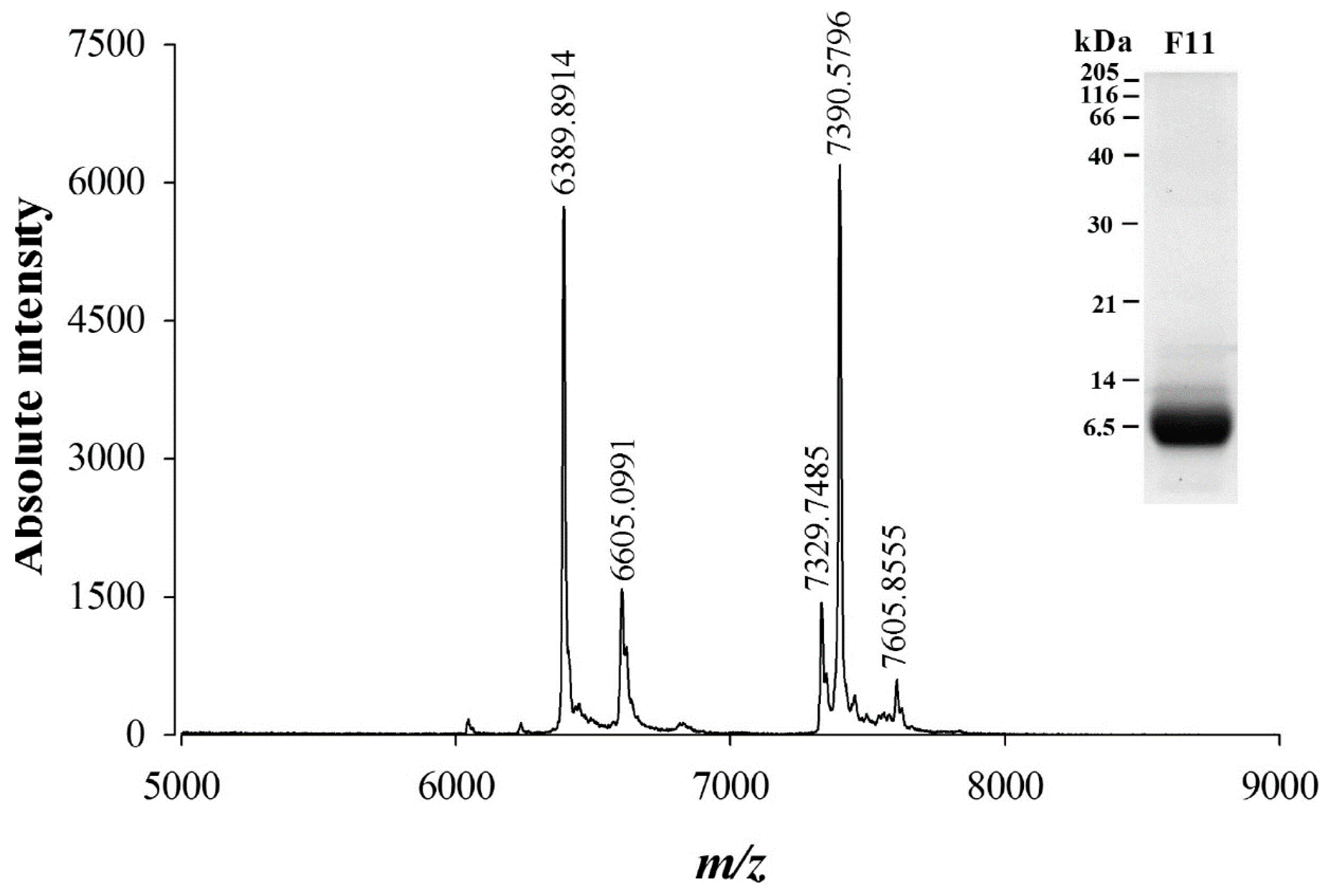
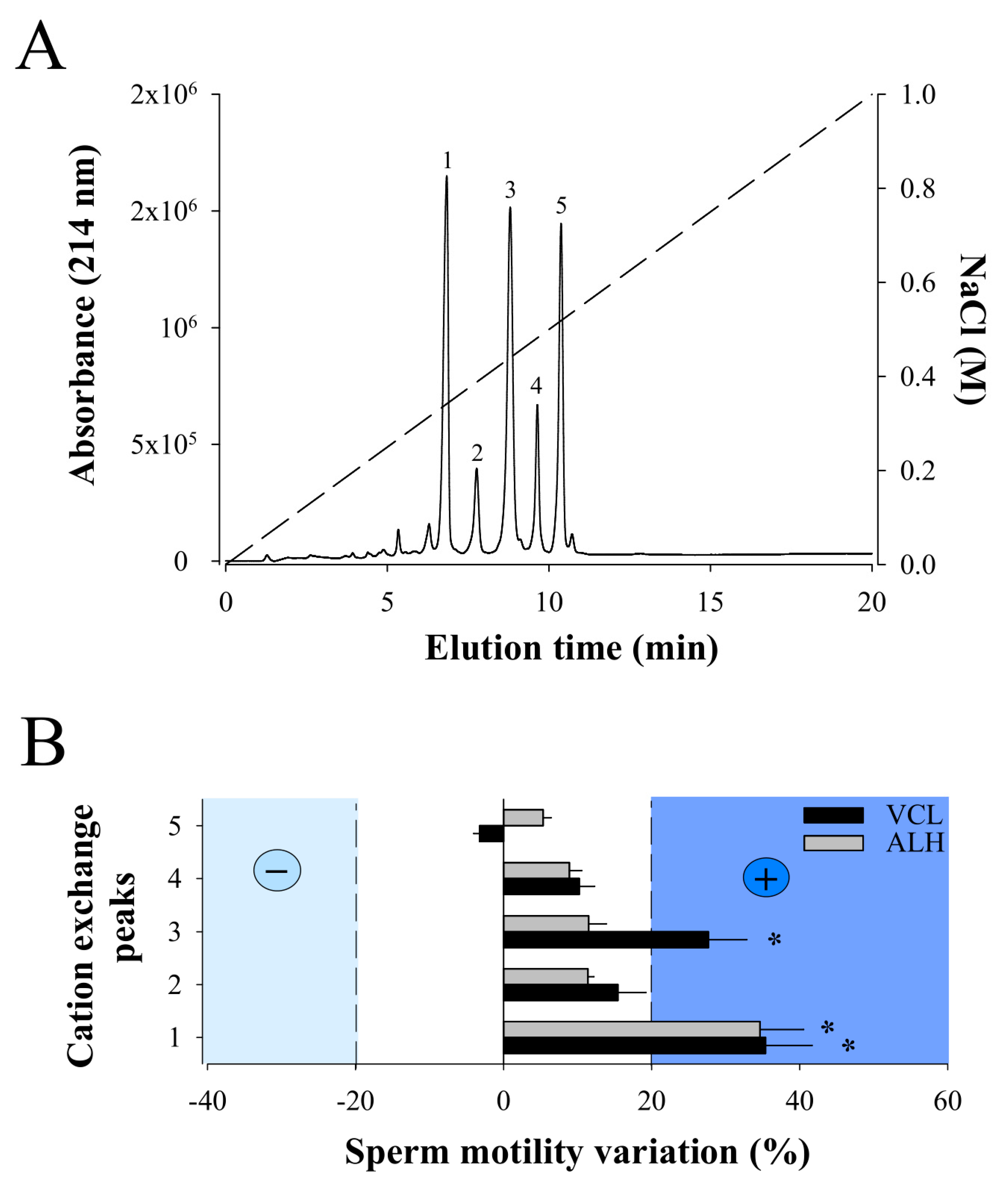
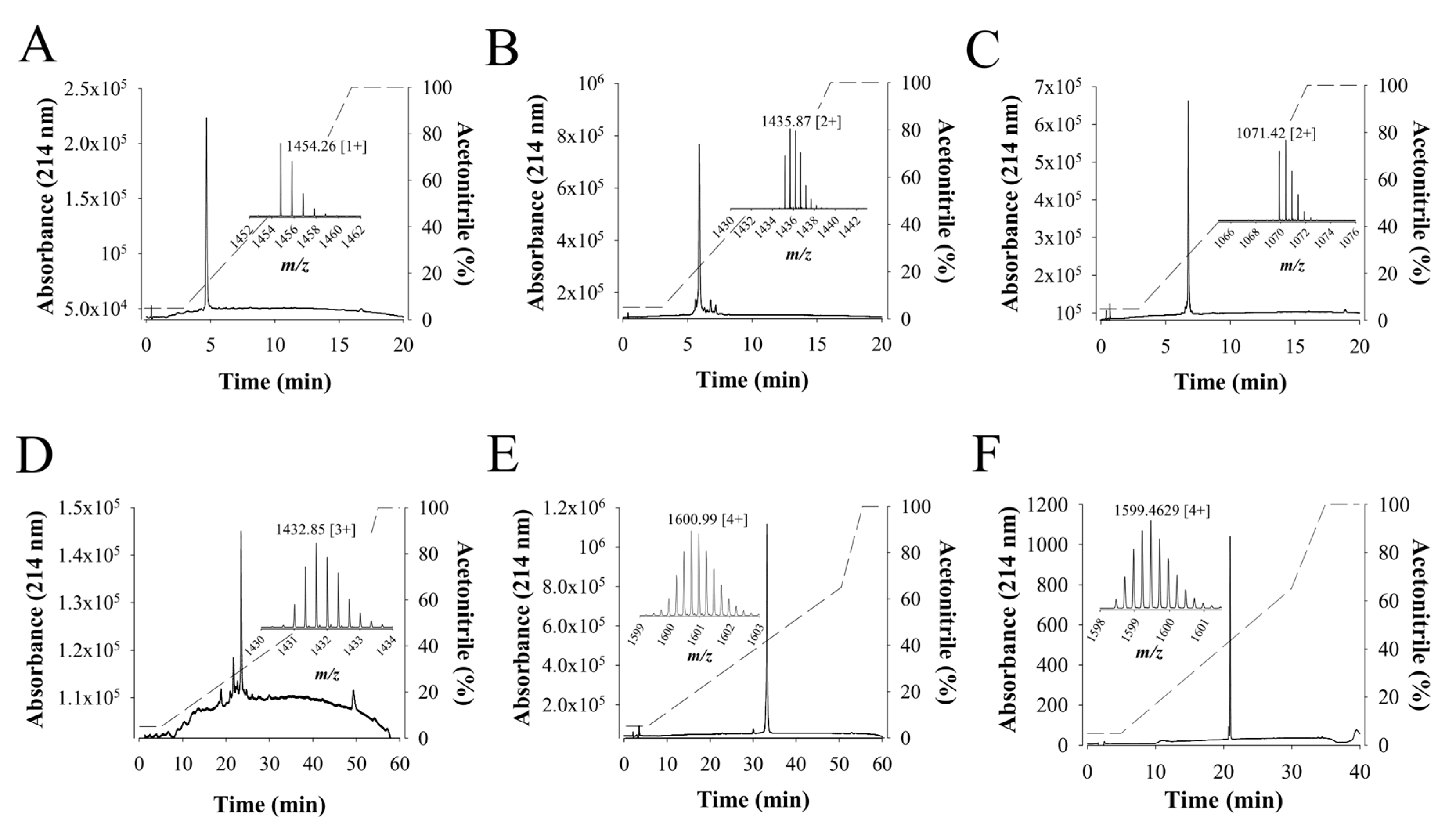
2.2. Cation Exchange Chromatography of F11 Leads to the Full Purification of a Bioactive Compound on Sperm Motility
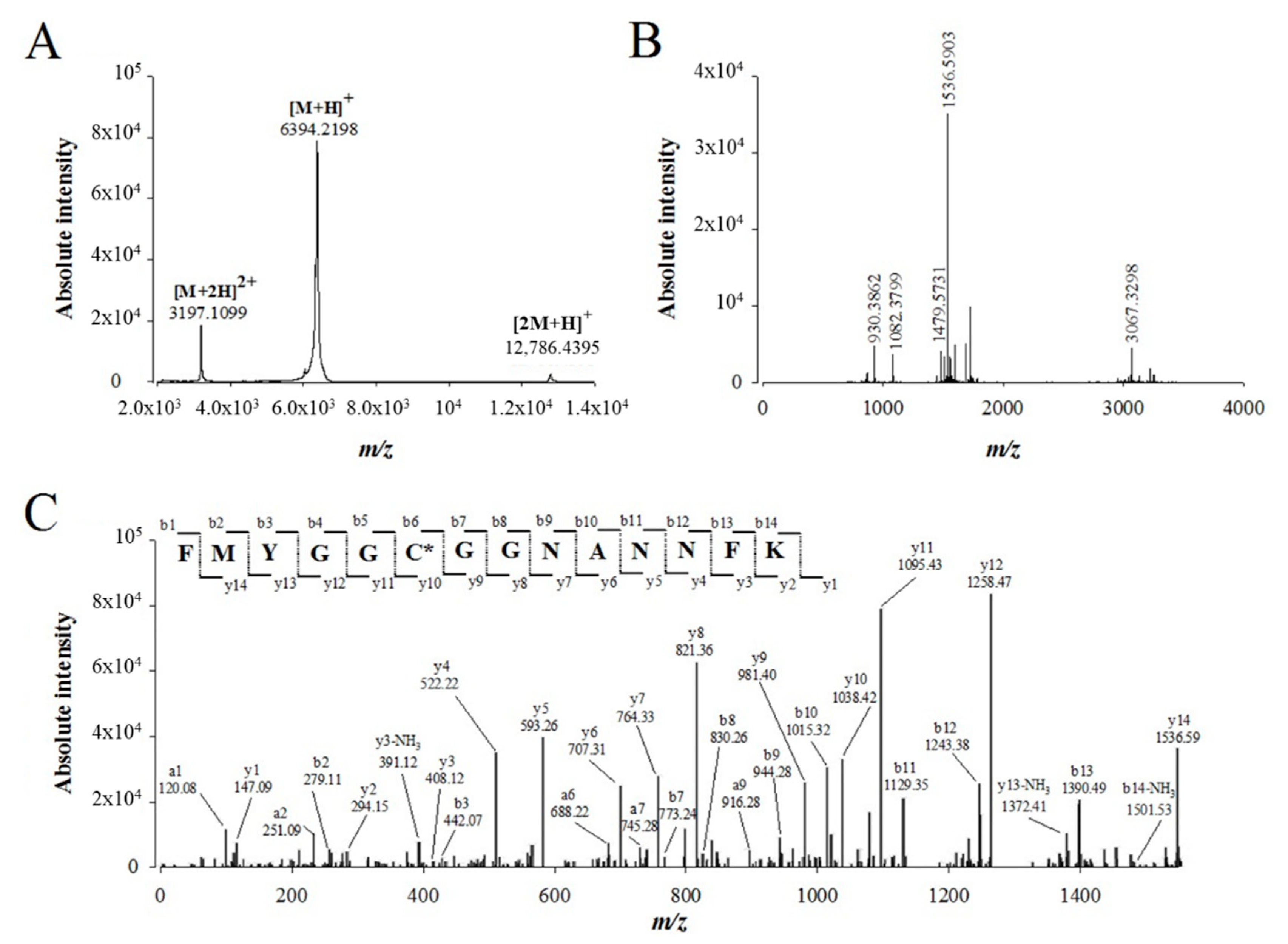
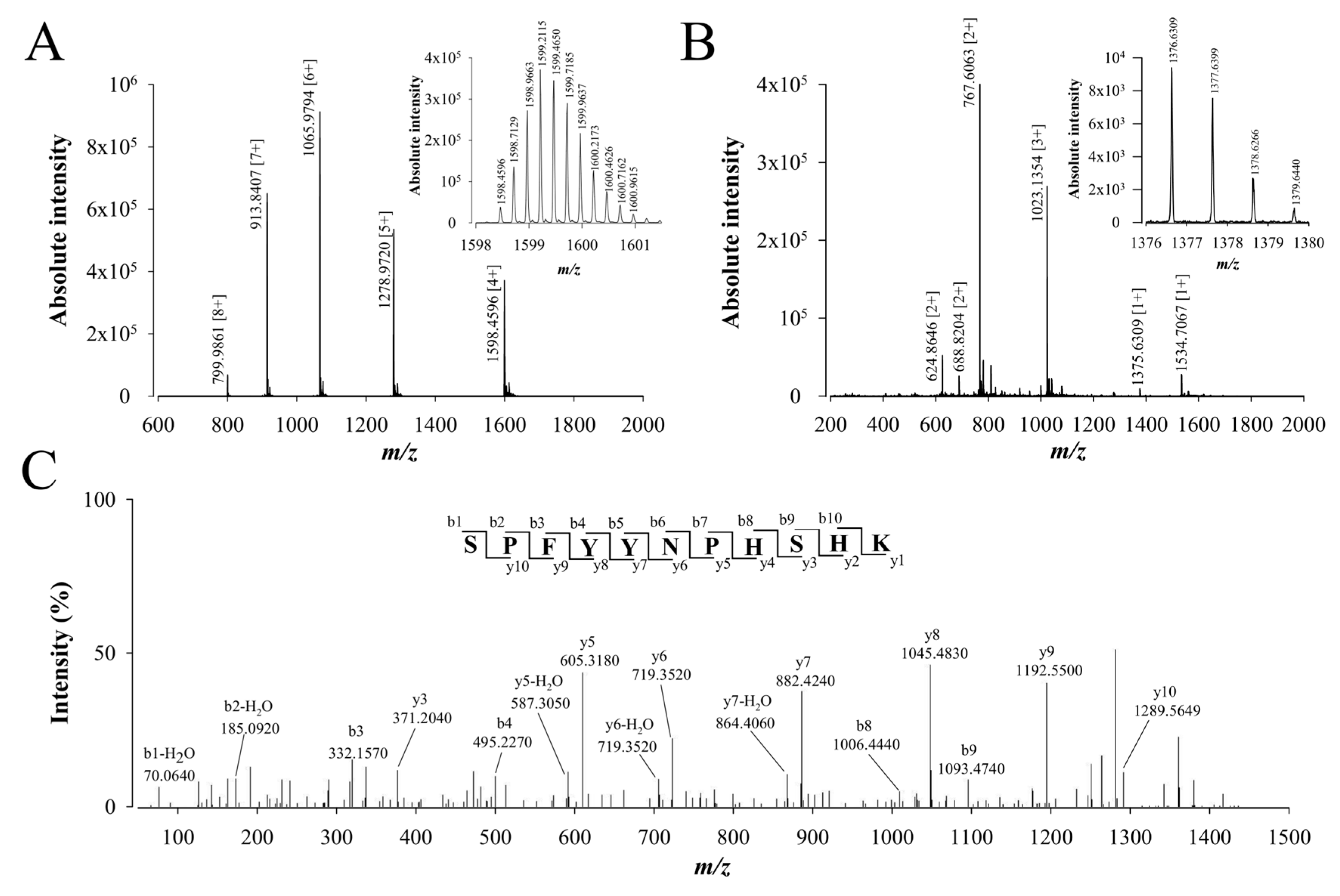
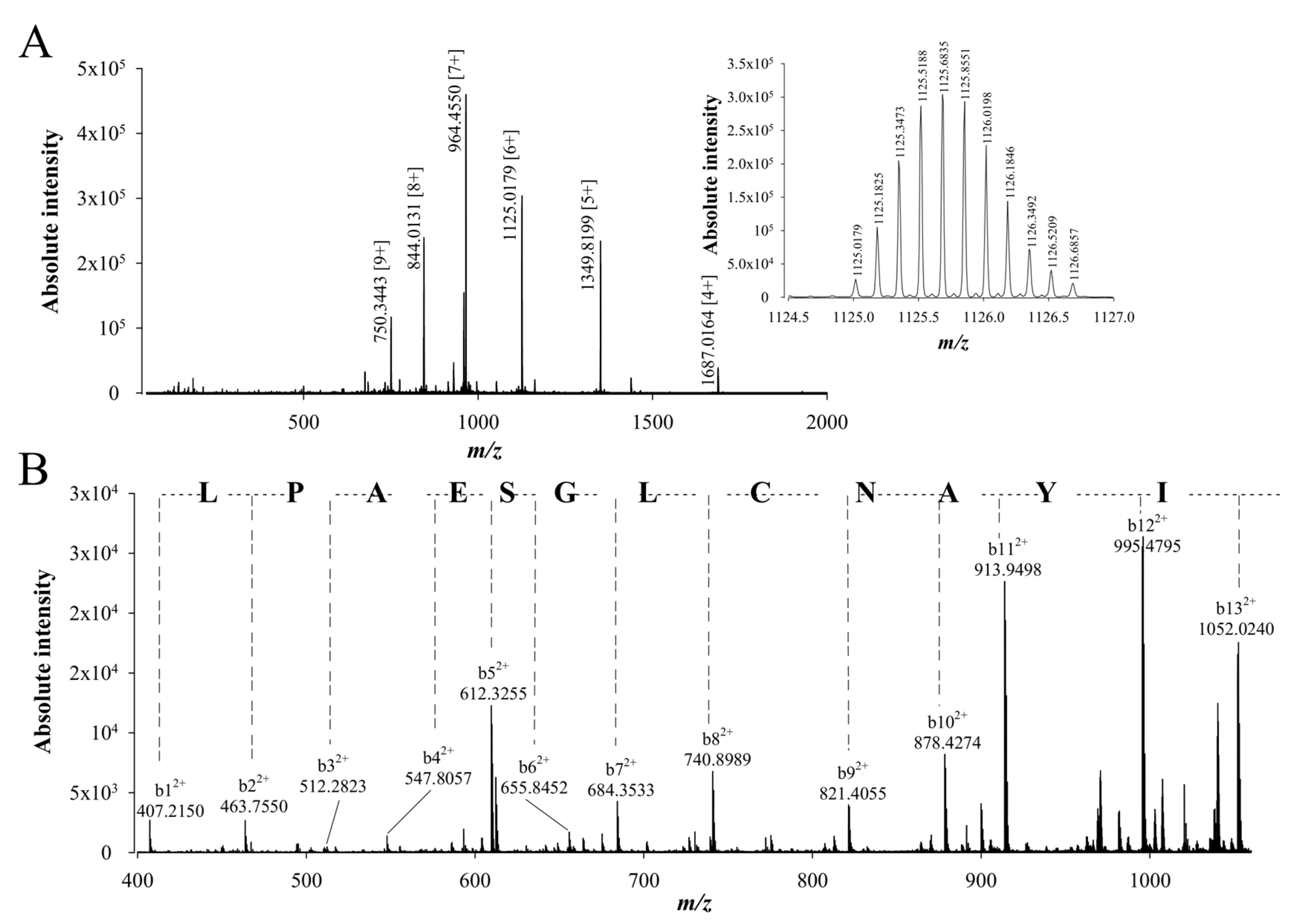
2.3. De Novo Sequencing of Walterospermin by MALDI-TOF/TOF MS/MS and LC-ESI-QTOF MS/MS
2.4. Walterospermin Amino Acid Sequence, Predicted Disulfide Bridges, and Sequence Homologies with Known Toxins
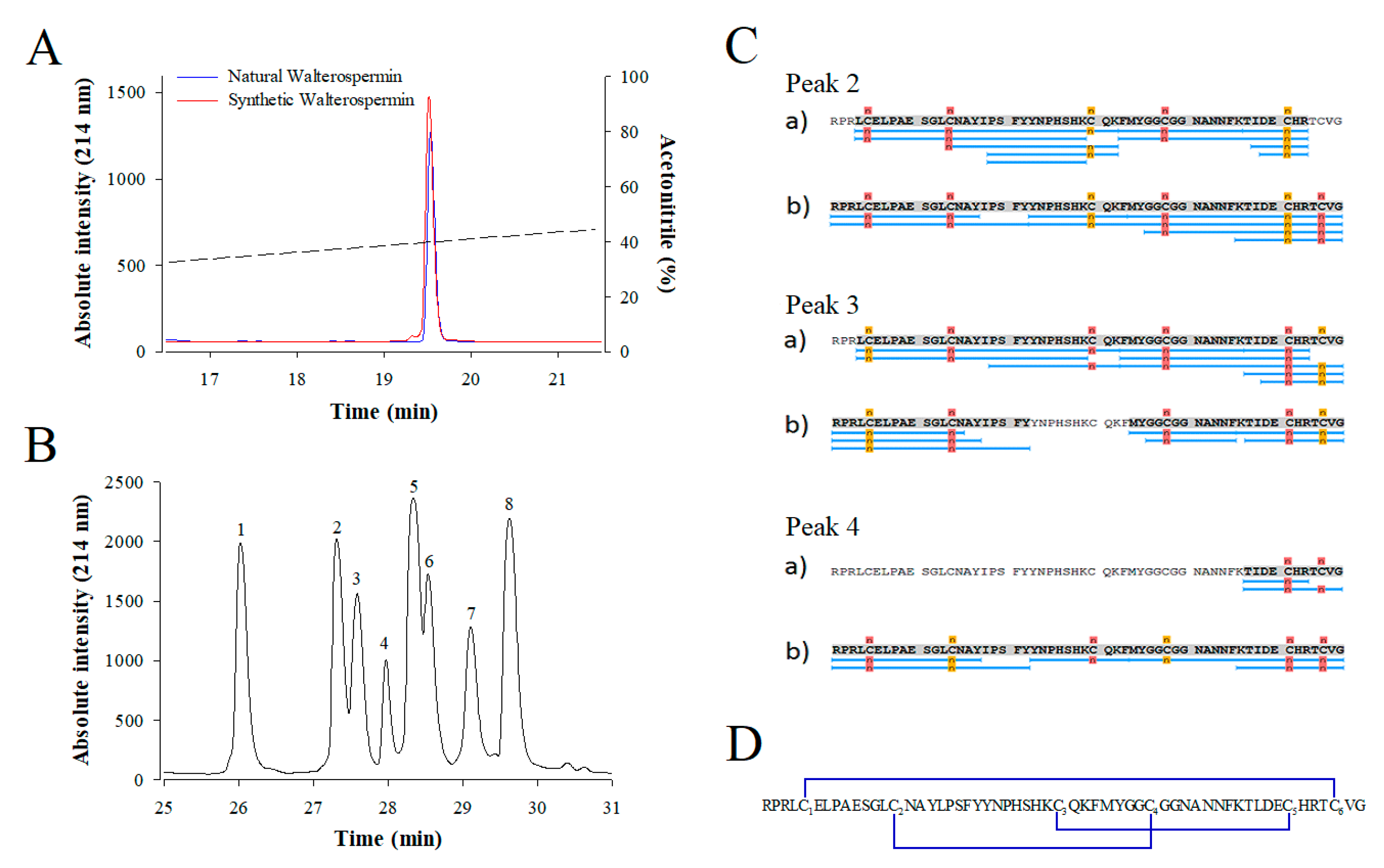
2.5. Chemical Synthesis of Walterospermin
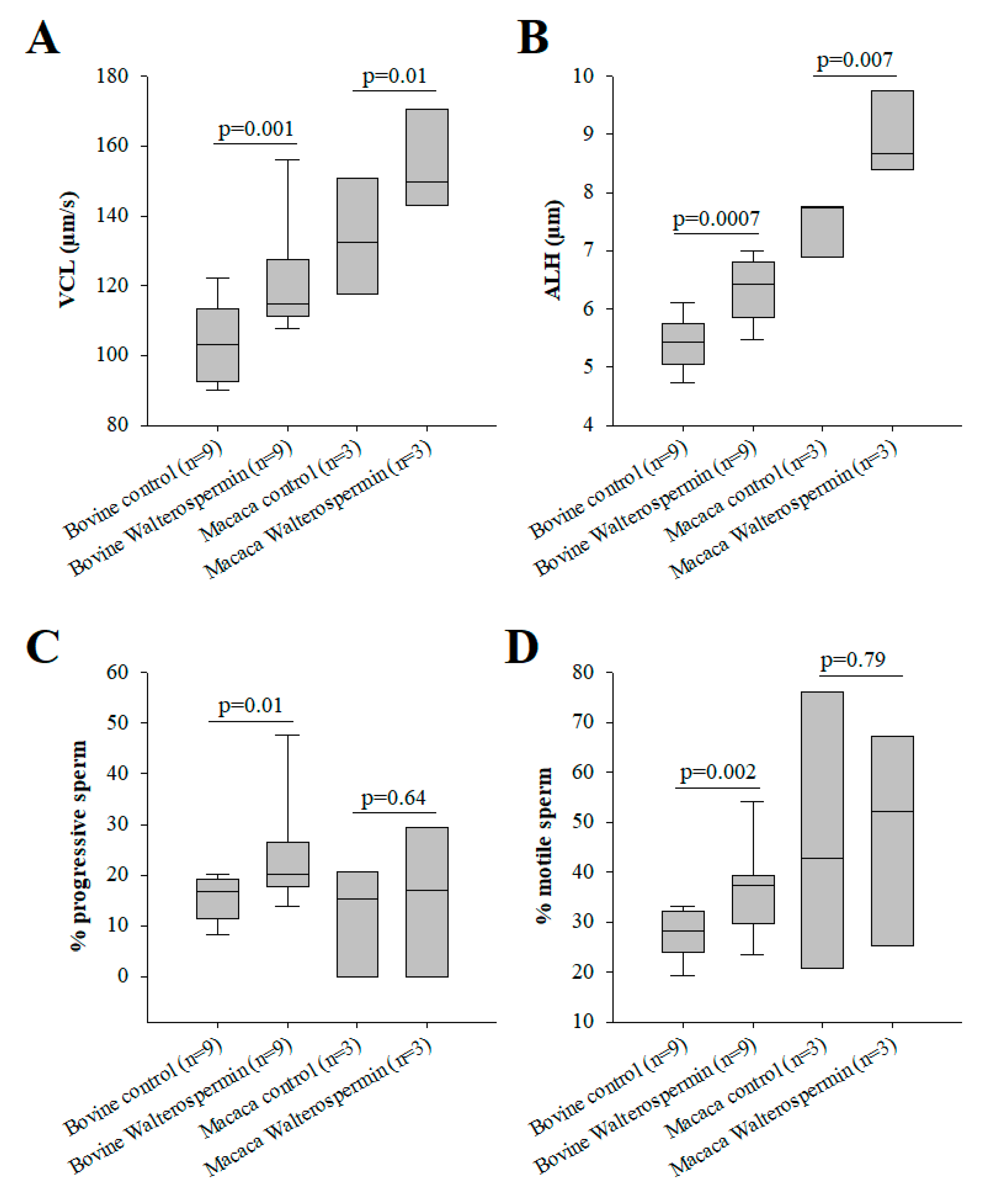
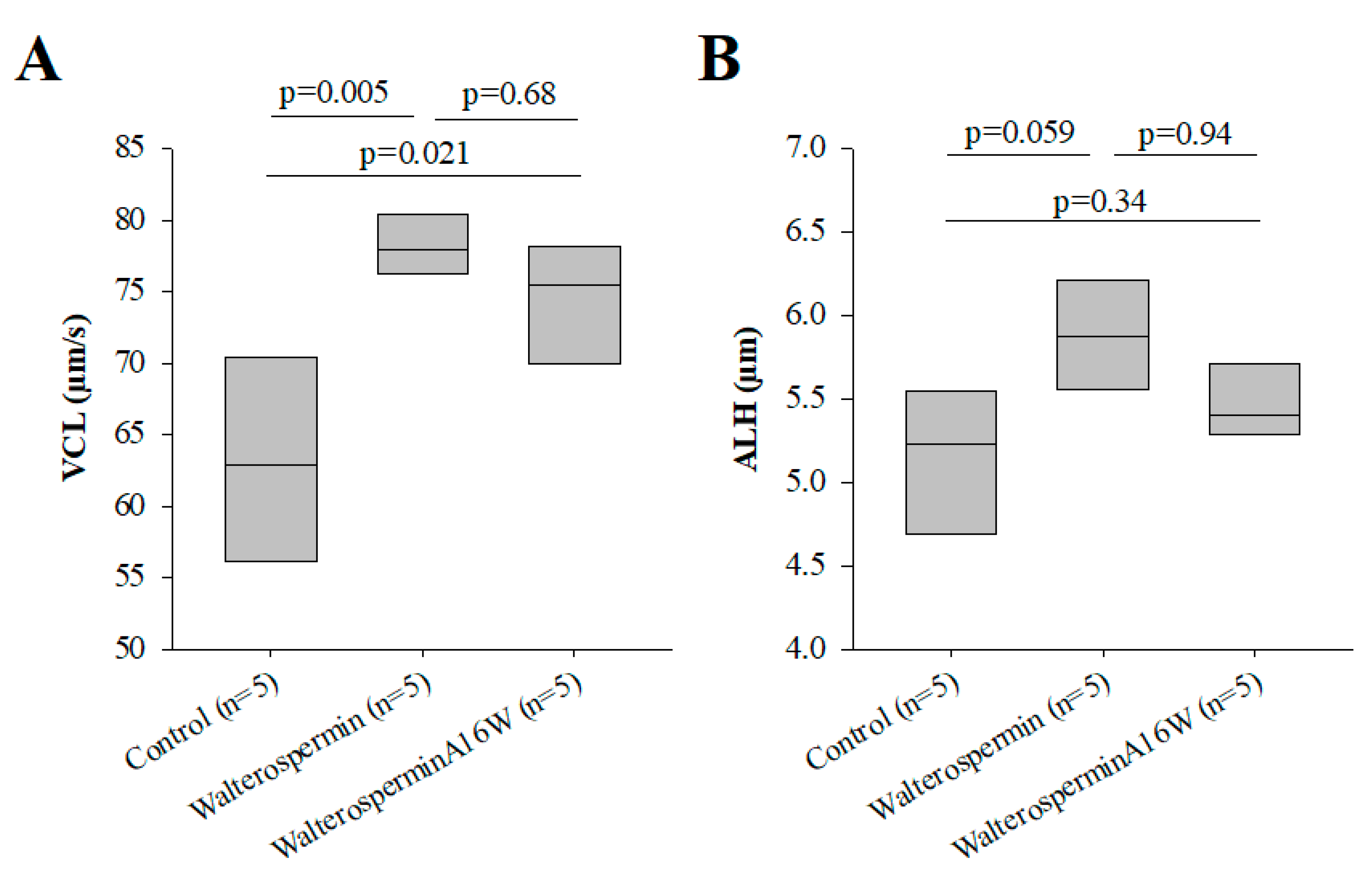
2.6. Functional Evaluation of Synthetic Walterospermin
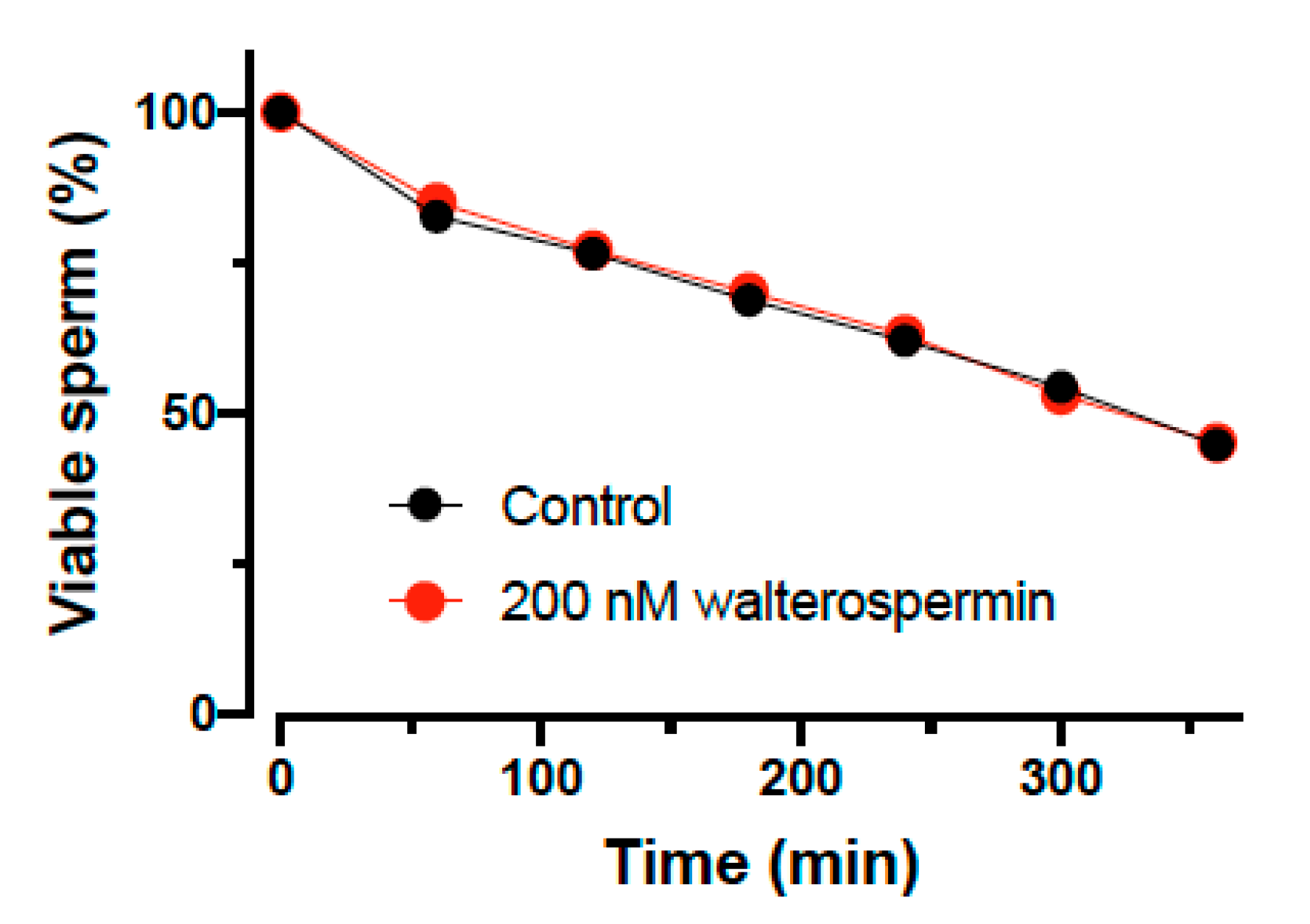
2.7. Impact of Walterospermin on Sperm Viability
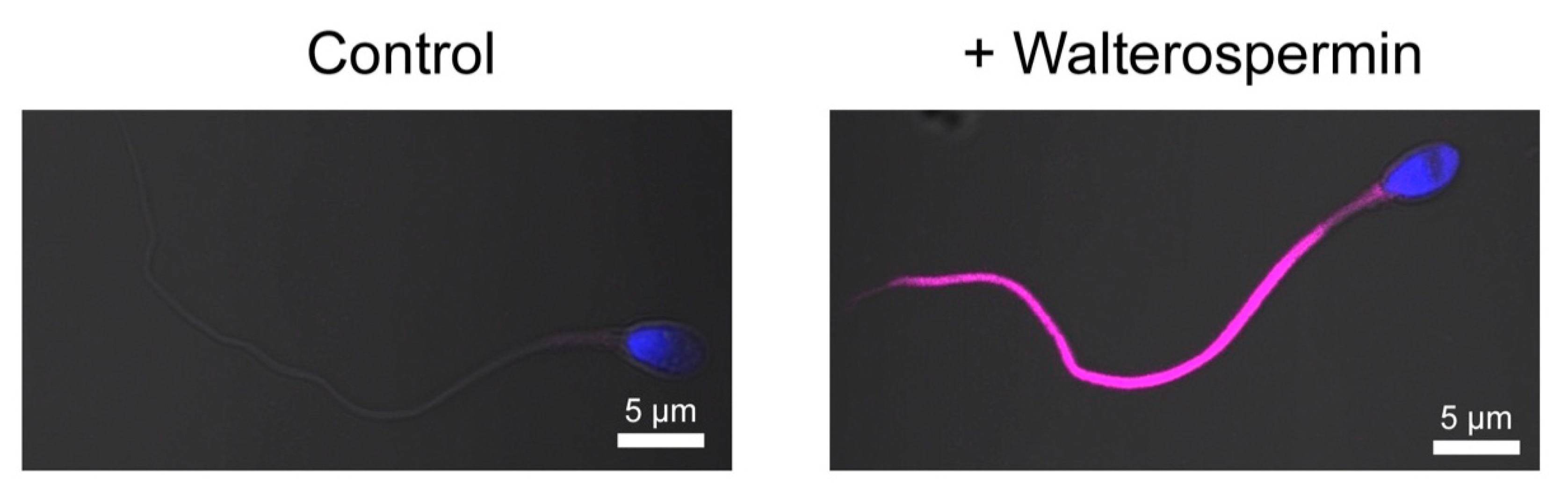
2.8. Fluorescent Synthetic Walterospermin Labels Sperm Flagellum
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. RP-HPLC Fractionation of the Snake Venom
4.3. Cation Exchange Chromatography
4.4. Biological Samples
4.5. Computer Assisted Semen Analysis (CASA)
4.6. Measurement of Vitality of Human Sperm Treated with Walterospermin
4.7. Amino Acid Sequence Characterization
4.7.1. MALDI-TOF MS Analyses
4.7.2. MALDI-TOF/TOF MS/MS Analyses
4.7.3. LC-ESI-QTOF MS/MS Analyses
4.7.4. Top-Down Venomics
4.7.5. Database Submission
4.7.6. Search of Peptides Analogs
4.8. Chemical Synthesis of Walterospermin
4.9. Determination of the Disulfide Bridge Organization of Walterospermin
4.10. Sperm Staining with Fluorescent Cy5-Walterospermin
4.11. Experimental Design and Statistical Rationale
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALH | amplitude of the lateral head displacement |
| CASA | computer-assisted semen analysis |
| CDAP | cyanodiaminopyridinium |
| CHCA | α-cyano-4-hydroxycinnamic acid |
| DDA | data-dependent acquisition method |
| ESI | electrospray ionization |
| FDA | food and drug administration |
| LC | liquid chromatography |
| MALDI | matrix-assisted laser desorption/ionization |
| MS | mass spectrometry |
| NCL | native chemical ligation |
| RP-HPLC | reversed-phase high performance liquid chromatography |
| TCEP | tris (2-carboxyethyl) phosphine hydrochloride |
| TOF | time of flight |
| VAP | average path velocity |
| VCL | curvilinear velocity |
| VSL | straight line velocity |
References
- Menez, A.; Stocklin, R.; Mebs, D. ‘Venomics’ or: The venomous systems genome project. Toxicon 2006, 47, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Porras, J.M. Pharmacology of peptides and proteins in snake venoms. Annu. Rev. Pharm. 1968, 8, 299–318. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Yamazaki, Y.; Brown, R.L.; Fujimoto, Z.; Morita, T.; Mizuno, H. Structures of pseudechetoxin and pseudecin, two snake-venom cysteine-rich secretory proteins that target cyclic nucleotide-gated ion channels: Implications for movement of the C-terminal cysteine-rich domain. Acta Cryst. D Biol. Cryst. 2008, 64, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Diochot, S.; Baron, A.; Salinas, M.; Douguet, D.; Scarzello, S.; Dabert-Gay, A.S.; Debayle, D.; Friend, V.; Alloui, A.; Lazdunski, M.; et al. Black mamba venom peptides target acid-sensing ion channels to abolish pain. Nature 2012, 490, 552–555. [Google Scholar] [CrossRef]
- Serafim, F.G.; Reali, M.; Cruz-Hofling, M.A.; Fontana, M.D. Action of Micrurus dumerilii carinicauda coral snake venom on the mammalian neuromuscular junction. Toxicon 2002, 40, 167–174. [Google Scholar] [CrossRef]
- Siew, J.P.; Khan, A.M.; Tan, P.T.; Koh, J.L.; Seah, S.H.; Koo, C.Y.; Chai, S.C.; Armugam, A.; Brusic, V.; Jeyaseelan, K. Systematic analysis of snake neurotoxins’ functional classification using a data warehousing approach. Bioinformatics 2004, 20, 3466–3480. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Carbajal-Saucedo, A.; Lopez-Vera, E.; Benard-Valle, M.; Smith, E.N.; Zamudio, F.; de Roodt, A.R.; Olvera-Rodriguez, A. Isolation, characterization, cloning and expression of an alpha-neurotoxin from the venom of the Mexican coral snake Micrurus laticollaris (Squamata: Elapidae). Toxicon 2013, 66, 64–74. [Google Scholar] [CrossRef]
- Munawar, A.; Trusch, M.; Georgieva, D.; Hildebrand, D.; Kwiatkowski, M.; Behnken, H.; Harder, S.; Arni, R.; Spencer, P.; Schluter, H.; et al. Elapid snake venom analyses show the specificity of the peptide composition at the level of genera Naja and Notechis. Toxins 2014, 6, 850–868. [Google Scholar] [CrossRef]
- Earl, S.T.; Masci, P.P.; de Jersey, J.; Lavin, M.F.; Dixon, J. Drug development from Australian elapid snake venoms and the Venomics pipeline of candidates for haemostasis: Textilinin-1 (Q8008), Haempatch (Q8009) and CoVase (V0801). Toxicon 2012, 59, 456–463. [Google Scholar] [CrossRef]
- Millers, E.K.; Johnson, L.A.; Birrell, G.W.; Masci, P.P.; Lavin, M.F.; de Jersey, J.; Guddat, L.W. The structure of human microplasmin in complex with textilinin-1, an aprotinin-like inhibitor from the Australian brown snake. PLoS ONE 2013, 8, e54104. [Google Scholar] [CrossRef]
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Williams, D.; Fan, H.W.; Warrell, D.A. Snakebite envenoming from a global perspective: Towards an integrated approach. Toxicon 2010, 56, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Sanz, L.; Angulo, Y.; Lomonte, B.; Gutierrez, J.M. Venoms, venomics, antivenomics. Febs Lett. 2009, 583, 1736–1743. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Garcia, M.L. Therapeutic potential of venom peptides. Nat. Rev. Drug Discov. 2003, 2, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Marcussi, S.; Sant’Ana, C.D.; Oliveira, C.Z.; Rueda, A.Q.; Menaldo, D.L.; Beleboni, R.O.; Stabeli, R.G.; Giglio, J.R.; Fontes, M.R.; Soares, A.M. Snake venom phospholipase A2 inhibitors: Medicinal chemistry and therapeutic potential. Curr. Top. Med. Chem. 2007, 7, 743–756. [Google Scholar] [CrossRef]
- Kapoor, V.K. Natural toxins and their therapeutic potential. Indian J. Exp. Biol. 2010, 48, 228–237. [Google Scholar]
- Biswas, A.; Gomes, A.; Sengupta, J.; Datta, P.; Singha, S.; Dasgupta, A.K.; Gomes, A. Nanoparticle-conjugated animal venom-toxins and their possible therapeutic potential. J. Venom Res. 2012, 3, 15–21. [Google Scholar]
- Hmed, B.; Serria, H.T.; Mounir, Z.K. Scorpion peptides: Potential use for new drug development. J. Toxicol. 2013, 2013, 958797. [Google Scholar] [CrossRef]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin, apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef]
- King, G.F. Venoms to Drugs: Venom as a Source for the Development of Human Therapeutics; Royal Society of Chemistry: Cambridge, UK, 2015; p. 17. [Google Scholar]
- Harvey, A.L. From venom to toxin to drug? Proc. R. Soc. Edinb. Sect. B Biol. Sci. 1992, 99, 55–65. [Google Scholar] [CrossRef]
- Fox, J.W.; Serrano, S.M. Approaching the golden age of natural product pharmaceuticals from venom libraries: An overview of toxins and toxin-derivatives currently involved in therapeutic or diagnostic applications. Curr. Pharm. Des. 2007, 13, 2927–2934. [Google Scholar] [CrossRef]
- Koh, C.Y.; Kini, R.M. From snake venom toxins to therapeutics—Cardiovascular examples. Toxicon 2012, 59, 497–506. [Google Scholar] [CrossRef]
- Takacs, Z.; Nathan, S. Animal Venoms in Medicine. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Academic Press: Oxford, UK, 2014; pp. 252–259. [Google Scholar] [CrossRef]
- McCleary, R.J.; Kini, R.M. Non-enzymatic proteins from snake venoms: A gold mine of pharmacological tools and drug leads. Toxicon 2013, 62, 56–74. [Google Scholar] [CrossRef] [PubMed]
- Mohamed Abd El-Aziz, T.; Garcia Soares, A.; Stockand, J.D. Snake Venoms in Drug Discovery: Valuable Therapeutic Tools for Life Saving. Toxins 2019, 11, 564. [Google Scholar] [CrossRef] [PubMed]
- Darszon, A.; Labarca, P.; Nishigaki, T.; Espinosa, F. Ion channels in sperm physiology. Physiol. Rev. 1999, 79, 481–510. [Google Scholar] [CrossRef] [PubMed]
- Martinez, G.; Hograindleur, J.P.; Voisin, S.; Abi Nahed, R.; Abd El Aziz, T.M.; Escoffier, J.; Bessonnat, J.; Fovet, C.M.; De Waard, M.; Hennebicq, S.; et al. Spermaurin, an La1-like peptide from the venom of the scorpion Scorpio maurus palmatus, improves sperm motility and fertilization in different mammalian species. Mol. Hum. Reprod. 2017, 23, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Escoffier, J.; Couvet, M.; de Pomyers, H.; Ray, P.F.; Seve, M.; Lambeau, G.; De Waard, M.; Arnoult, C. Snake venoms as a source of compounds modulating sperm physiology: Secreted phospholipases A2 from Oxyuranus scutellatus scutellatus impact sperm motility, acrosome reaction and in vitro fertilization in mice. Biochimie 2010, 92, 826–836. [Google Scholar] [CrossRef]
- Curi, S.M.; Ariagno, J.I.; Chenlo, P.H.; Mendeluk, G.R.; Pugliese, M.N.; Sardi Segovia, L.M.; Repetto, H.E.; Blanco, A.M. Asthenozoospermia: Analysis of a large population. Arch. Androl. 2003, 49, 343–349. [Google Scholar] [CrossRef]
- Ortega, C.; Verheyen, G.; Raick, D.; Camus, M.; Devroey, P.; Tournaye, H. Absolute asthenozoospermia and ICSI: What are the options? Hum. Reprod. Update 2011, 17, 684–692. [Google Scholar] [CrossRef]
- Ravaux, B.; Garroum, N.; Perez, E.; Willaime, H.; Gourier, C. A specific flagellum beating mode for inducing fusion in mammalian fertilization and kinetics of sperm internalization. Sci. Rep. 2016, 6, 31886. [Google Scholar] [CrossRef]
- Publicover, S.J.; Barratt, C.L. Sperm motility: Things are moving in the lab! Mol. Hum. Reprod. 2011, 17, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.P.; Wang, H.F.; Li, B.M.; Zeng, X.H. Sperm-specific ion channels: Targets holding the most potential for male contraceptives in development. Contraception 2013, 88, 485–491. [Google Scholar] [CrossRef]
- Abd El Aziz, T.M.; Bourgoin-Voillard, S.; Combemale, S.; Beroud, R.; Fadl, M.; Seve, M.; De Waard, M. Fractionation and proteomic analysis of the Walterinnesia aegyptia snake venom using OFFGEL and MALDI-TOF-MS techniques. Electrophoresis 2015, 36, 2594–2605. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.Y.; Wang, Y.M.; Tsai, I.H. Cloning, characterization and phylogenetic analyses of members of three major venom families from a single specimen of Walterinnesia aegyptia. Toxicon 2008, 51, 1245–1254. [Google Scholar] [CrossRef]
- He, Y.-Y.; Liu, S.-B.; Lee, W.-H.; Qian, J.-Q.; Zhang, Y. Isolation, expression and characterization of a novel dual serine protease inhibitor, OH-TCI, from king cobra venom. Peptides 2008, 29, 1692–1699. [Google Scholar] [CrossRef]
- Peigneur, S.; Billen, B.; Derua, R.; Waelkens, E.; Debaveye, S.; Beress, L.; Tytgat, J. A bifunctional sea anemone peptide with Kunitz type protease and potassium channel inhibiting properties. Biochem. Pharm. 2011, 82, 81–90. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Hu, Y.T.; Yang, W.S.; He, Y.W.; Feng, J.; Wang, B.; Zhao, R.M.; Ding, J.P.; Cao, Z.J.; Li, W.X.; et al. Hg1, novel peptide inhibitor specific for Kv1.3 channels from first scorpion Kunitz-type potassium channel toxin family. J. Biol. Chem. 2012, 287, 13813–13821. [Google Scholar] [CrossRef] [PubMed]
- Soares, T.S.; Watanabe, R.M.; Tanaka-Azevedo, A.M.; Torquato, R.J.; Lu, S.; Figueiredo, A.C.; Pereira, P.J.; Tanaka, A.S. Expression and functional characterization of boophilin, a thrombin inhibitor from Rhipicephalus (Boophilus) microplus midgut. Vet. Parasitol. 2012, 187, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, H.; Kotsyfakis, M.; Francischetti, I.M.; Eum, J.H.; Strand, M.R.; Champagne, D.E. Simukunin from the salivary glands of the black fly Simulium vittatum inhibits enzymes that regulate clotting and inflammatory responses. PLoS ONE 2012, 7, e29964. [Google Scholar] [CrossRef]
- Choo, Y.M.; Lee, K.S.; Yoon, H.J.; Qiu, Y.; Wan, H.; Sohn, M.R.; Sohn, H.D.; Jin, B.R. Antifibrinolytic role of a bee venom serine protease inhibitor that acts as a plasmin inhibitor. PLoS ONE 2012, 7, e32269. [Google Scholar] [CrossRef] [PubMed]
- Grzesiak, A.; Helland, R.; Smalås, A.O.; Krowarsch, D.; Dadlez, M.; Otlewski, J. Substitutions at the P1’ position in BPTI strongly affect the association energy with serine proteinases1. J. Mol. Biol. 2000, 301, 205–217. [Google Scholar] [CrossRef]
- de Lamirande, E.; Gagnon, C. Effects of protease inhibitors and substrates on motility of mammalian spermatozoa. J. Cell Biol. 1986, 102, 1378–1383. [Google Scholar] [CrossRef]
- Harvey, A.L.; Robertson, B. Dendrotoxins: Structure-activity relationships and effects on potassium ion channels. Curr. Med. Chem. 2004, 11, 3065–3072. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L. Twenty years of dendrotoxins. Toxicon 2001, 39, 15–26. [Google Scholar] [CrossRef]
- Xia, J.; Reigada, D.; Mitchell, C.H.; Ren, D. CATSPER channel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagation. Biol. Reprod. 2007, 77, 551–559. [Google Scholar] [CrossRef]
- Qi, H.; Moran, M.M.; Navarro, B.; Chong, J.A.; Krapivinsky, G.; Krapivinsky, L.; Kirichok, Y.; Ramsey, I.S.; Quill, T.A.; Clapham, D.E. All four CatSper ion channel proteins are required for male fertility and sperm cell hyperactivated motility. Proc. Natl. Acad. Sci. USA 2007, 104, 1219–1223. [Google Scholar] [CrossRef]
- Avenarius, M.R.; Hildebrand, M.S.; Zhang, Y.; Meyer, N.C.; Smith, L.L.; Kahrizi, K.; Najmabadi, H.; Smith, R.J. Human male infertility caused by mutations in the CATSPER1 channel protein. Am. J. Hum. Genet. 2009, 84, 505–510. [Google Scholar] [CrossRef]
- Navarro, B.; Kirichok, Y.; Clapham, D.E. KSper, a pH-sensitive K+ current that controls sperm membrane potential. Proc. Natl. Acad. Sci. USA 2007, 104, 7688–7692. [Google Scholar] [CrossRef]
- Lishko, P.V.; Botchkina, I.L.; Fedorenko, A.; Kirichok, Y. Acid extrusion from human spermatozoa is mediated by flagellar voltage-gated proton channel. Cell 2010, 140, 327–337. [Google Scholar] [CrossRef]
- Abd El-Aziz, T.M.; Al Khoury, S.; Jaquillard, L.; Triquigneaux, M.; Martinez, G.; Bourgoin-Voillard, S.; Sève, M.; Arnoult, C.; Beroud, R.; De Waard, M. Actiflagelin, a new sperm activator isolated from Walterinnesia aegyptia venom using phenotypic screening. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 2. [Google Scholar] [CrossRef] [PubMed]
- Samejima, Y.; Aoki-Tomomatsu, Y.; Yanagisawa, M.; Mebs, D. Amino acid sequence of two neurotoxins from the venom of the Egyptian black snake (Walterinnesia aegyptia). Toxicon 1997, 35, 151–157. [Google Scholar] [CrossRef]
- Simon, T.; Bdolah, A. Isolation of phospholipase A from the venom of the desert black snake Walterinnesia aegyptia. Toxicon 1980, 18, 369–373. [Google Scholar] [CrossRef]
- Duhaiman, A.S.; Alhomida, A.S.; Rabbani, N.; Kamal, M.A.; Al-Jafari, A.A. Purification and characterization of acetylcholinesterase from desert cobra (Walterinnesia aegyptia) venom. Biochimie 1996, 78, 46–50. [Google Scholar] [CrossRef]
- Badr, G.; Al-Sadoon, M.K.; El-Toni, A.M.; Daghestani, M. Walterinnesia aegyptia venom combined with silica nanoparticles enhances the functioning of normal lymphocytes through PI3K/AKT, NFkappaB and ERK signaling. Lipids Health Dis. 2012, 11, 27. [Google Scholar] [CrossRef]
- Badr, G.; Al-Sadoon, M.K.; Rabah, D.M.; Sayed, D. Snake (Walterinnesia aegyptia) venom-loaded silica nanoparticles induce apoptosis and growth arrest in human prostate cancer cells. Apoptosis 2013, 18, 300–314. [Google Scholar] [CrossRef]
- Al-Sadoon, M.K.; Rabah, D.M.; Badr, G. Enhanced anticancer efficacy of snake venom combined with silica nanoparticles in a murine model of human multiple myeloma: Molecular targets for cell cycle arrest and apoptosis induction. Cell. Immunol. 2013, 284, 129–138. [Google Scholar] [CrossRef]
- Bringans, S.; Eriksen, S.; Kendrick, T.; Gopalakrishnakone, P.; Livk, A.; Lock, R.; Lipscombe, R. Proteomic analysis of the venom of Heterometrus longimanus (Asian black scorpion). Proteomics 2008, 8, 1081–1096. [Google Scholar] [CrossRef]
- Vizcaino, J.A.; Deutsch, E.W.; Wang, R.; Csordas, A.; Reisinger, F.; Rios, D.; Dianes, J.A.; Sun, Z.; Farrah, T.; Bandeira, N.; et al. ProteomeXchange provides globally coordinated proteomics data submission and dissemination. Nat. Biotechnol. 2014, 32, 223–226. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Precursor Ion (m/z) | Charge Number (z) | MS/MS-Derived Sequence | Protease Used & Mode of Measurement |
|---|---|---|---|
| 1536.60 | 1 | FMYGGC(+57.02)GGNANNFK | Trypsin - MALDI- TOF/TOF MS/MS |
| 930.39 | 1 | TIDEC(+57.02)HR | |
| 2384.07 | 1 | ESGLC(+57.02)EKGFPSFYYNPHSHK | |
| 768.82 | 2 | FMYGGC(+57.02)GGNANNFK | Trypsin - LC-ESI-QTOF MS/MS |
| 465.71 | 2 | TIDEC(+57.02)HR | |
| 688.82 | 2 | SPFYYNPHSHK | |
| 570.26 | 3 | LC(+57.02)ELPAESGLC(+57.02)NAYI | |
| 723.34 | 2 | YNPHSHKC(+57.02)QKF | Chymotrypsin - LC-ESI-QTOF MS/MS |
| 738.34 | 2 | KTIDECHRTC(+57.02)VG | |
| 707.64 | 3 | KFMYGGC(+57.02)GGNANNFKTIDE | |
| 620.82 | 2 | RPRLC(+57.02)ELPAE | V8 protease - LC-ESI-QTOF MS/MS |
| 749.32 | 2 | GC(+57.02)GGNANNFKTIDE | |
| 76.35 | 2 | PHSHKC(+57.02)QKFMYG | |
| 445.19 | 2 | C(+57.02)HRTC(+57.02)VG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd El-Aziz, T.M.; Jaquillard, L.; Bourgoin-Voillard, S.; Martinez, G.; Triquigneaux, M.; Zoukimian, C.; Combemale, S.; Hograindleur, J.-P.; Al Khoury, S.; Escoffier, J.; et al. Identification, Characterization and Synthesis of Walterospermin, a Sperm Motility Activator from the Egyptian Black Snake Walterinnesia aegyptia Venom. Int. J. Mol. Sci. 2020, 21, 7786. https://doi.org/10.3390/ijms21207786
Abd El-Aziz TM, Jaquillard L, Bourgoin-Voillard S, Martinez G, Triquigneaux M, Zoukimian C, Combemale S, Hograindleur J-P, Al Khoury S, Escoffier J, et al. Identification, Characterization and Synthesis of Walterospermin, a Sperm Motility Activator from the Egyptian Black Snake Walterinnesia aegyptia Venom. International Journal of Molecular Sciences. 2020; 21(20):7786. https://doi.org/10.3390/ijms21207786
Chicago/Turabian StyleAbd El-Aziz, Tarek Mohamed, Lucie Jaquillard, Sandrine Bourgoin-Voillard, Guillaume Martinez, Mathilde Triquigneaux, Claude Zoukimian, Stéphanie Combemale, Jean-Pascal Hograindleur, Sawsan Al Khoury, Jessica Escoffier, and et al. 2020. "Identification, Characterization and Synthesis of Walterospermin, a Sperm Motility Activator from the Egyptian Black Snake Walterinnesia aegyptia Venom" International Journal of Molecular Sciences 21, no. 20: 7786. https://doi.org/10.3390/ijms21207786
APA StyleAbd El-Aziz, T. M., Jaquillard, L., Bourgoin-Voillard, S., Martinez, G., Triquigneaux, M., Zoukimian, C., Combemale, S., Hograindleur, J.-P., Al Khoury, S., Escoffier, J., Michelland, S., Bulet, P., Beroud, R., Seve, M., Arnoult, C., & De Waard, M. (2020). Identification, Characterization and Synthesis of Walterospermin, a Sperm Motility Activator from the Egyptian Black Snake Walterinnesia aegyptia Venom. International Journal of Molecular Sciences, 21(20), 7786. https://doi.org/10.3390/ijms21207786