Abstract
Inflammatory bowel disease (IBD) includes Crohn’s disease (CD) and ulcerative colitis (UC). These are chronic autoimmune diseases of unknown etiology affecting the gastrointestinal tract. The IBD population includes a heterogeneous group of patients with varying disease courses requiring personalized treatment protocols. The complexity of the disease often delays the diagnosis and the initiation of appropriate treatments. In a subset of patients, IBD leads to colitis-associated cancer (CAC). MicroRNAs are single-stranded regulatory noncoding RNAs of 18 to 22 nucleotides with putative roles in the pathogenesis of IBD and colorectal cancer. They have been explored as biomarkers and therapeutic targets. Both tissue-derived and circulating microRNAs have emerged as promising biomarkers in the differential diagnosis and in the prognosis of disease severity of IBD as well as predictive biomarkers in drug resistance. In addition, knowledge of the cellular localization of differentially expressed microRNAs is a prerequisite for deciphering the biological role of these important epigenetic regulators and the cellular localization may even contribute to an alternative repertoire of biomarkers. In this review, we discuss findings based on RT-qPCR, microarray profiling, next generation sequencing and in situ hybridization of microRNA biomarkers identified in the circulation and in tissue biopsies.
1. Introduction
Inflammatory bowel disease (IBD) refers to Crohn’s disease (CD) and ulcerative colitis (UC). In UC, inflammation generally includes the rectum and extends towards the coecum and remains confined to the colon. In contrast, in CD, inflammation can involve any part of the gastrointestinal tract (GI) from the oral cavity to the anus. Both CD and UC are associated with multiple pathogenic factors such as environmental changes, the array of susceptibility gene variants, qualitatively and quantitatively abnormal gut microbiota and broadly dysregulated immune response [1]. Although CD and UC have some common pathological and clinical characteristics, they have several different attributes that imply that they are two distinct disease subtypes. In CD, fissuring ulceration and sub-mucosal fibrosis can be observed along with granulomatous inflammation. In UC, the inflammatory process always involves the rectum [2] and general histological findings include crypt distortion, infiltration of lymphocytes and granulocytes and chronic inflammation, usually confined to the lamina propria [3]. The diagnosis of IBD is usually established by a collective assessment of clinical presentation and endoscopic, histopathological, radiographic and laboratory findings. A definitive diagnosis of IBD cannot be made without detailed endoscopic and histologic assessment [4]. However, a subset of IBD cases cannot be classified as either CD or UC but are categorized as IBD unclassified (IBDU). Molecular biomarkers may support differential diagnosis of IBDU cases into CD or UC, or even be helpful in determining if IBDU represents a unique IBD diagnostic entity.
IBD starts developing at a younger age, including in infants [5], and is often characterized by a considerable diagnostic and therapeutic challenge because of the disease’s clinical features and associated complications. The prevalence of IBD in the Western world is projected to be up to 0.5% of the overall population [6]. In Denmark, where one of the highest annual incidence rates of IBD in Europe is seen, the incidence has been increasing over the last three decades [7]. In 2013, the incidence was 9.1 per 100,000 persons and 18.6 per 100,000 persons for CD and UC, respectively [8]. Since the turn of the 21st century, IBD has become a global disease with accelerating incidence rates also in developing countries whose societies have adopted a western diet and lifestyle. Although the incidence rate has become steady in western countries, the burden remains high, as prevalence exceeds 0.3%. The chronical inflammatory condition in the affected colon of IBD patients has been linked to development of neoplastic lesions in the colon. Several studies have shown a higher incidence of colorectal cancer (CRC) in IBD patients [9,10,11]. No biomarkers exist for the identification of IBD patients at risk of developing colitis-associated cancer (CAC), strongly advocating for more translational research in this field.
In this review, we give an overview of microRNAs (miRNAs) as candidate biomarkers in the IBD diagnostic assessment. Changes in miRNA levels are associated with disease development and can be measured both within the diseased tissue and in the circulation by a variety of molecular methods. MiRNAs have been found to be well conserved in archived tissue specimens, enabling retrospective analyses of clinical sample cohorts.
2. MicroRNA—An Introduction
MiRNAs play a central role in the regulation of several immune-mediated disorders including IBD [12,13,14]. MiRNAs are a group of small noncoding RNAs, approximately 18–22 nucleotides [15] which are found conserved across species. Their discovery was first described first in 1993 in Caenorhabditis elegans [16]. MiRNAs are transcribed as primary transcripts by RNA polymerase, processed into a precursor miRNA by the RNase III enzyme, Drosha, and exported from the nucleus to the cytoplasm. The precursor miRNA is cleaved by the RNase III enzyme, Dicer, into its mature form, which becomes stably incorporated into an RNA induced silencing complex (RISC). The miRNA guides the binding of the RNA-induced silencing complex to complementary sequences in the 3′-untranslated regions (UTR) of target mRNA molecules, resulting in either mRNA degradation or translational inhibition [17]. During stages of miRNA biogenesis, several factors can influence the development of the mature miRNA. These include regulation of transcription, cleavage of the stem loop structures by Drosha and Dicer enzymes, and editing as well as regulation of miRNA turnover. Each of these mechanisms acts as part of a signaling network that modulates gene expression in response to cellular or environmental changes.
MiRNA expression has been shown to be of importance in a wide variety of human diseases such as cancer, autoimmune, cardiovascular, and neurodegenerative diseases [14,18,19,20,21,22,23,24]. The miRNAs not only circulate in the human peripheral blood in a stable form, they are also present in other body fluids such as urine, saliva, milk, cerebrospinal fluid, and feces [25,26,27,28]. The miRNAs are engaged in disease origin and development, and some are pathology-specific [29], thus, changes in miRNA expression profiles have been addressed for applications in early detection as well as prognostics, diagnostic classification and drug response prediction.
3. MiRNAs in IBD
In IBD, miRNAs have been found to be involved in pathogenesis and have been identified as both diagnostic biomarkers and therapeutic targets [30]. Most of the recent research in the IBD field has measured levels of circulating miRNAs in body fluids such as blood or feces, and in homogenized tissue biopsies using techniques like microarray profiling, RT-qPCR, and NGS [27,31,32,33,34]. Studies have also performed tissue miRNA expression analysis using in situ hybridization (ISH) methods [35,36,37]. ISH methods for expression analyses of miRNAs can determine the cellular origin of miRNA expression and can offer insight into the biology of the disease mechanisms involved. Local expression levels of miRNAs can greatly vary from those of circulating miRNAs, e.g., due to contribution of miRNAs from circulating cells. Esquela-Kerscher and Slack [38] proposed that tumor cells release miRNAs into the neighboring microenvironment and enter circulation during angiogenesis. Some studies suggest that this likely occurs through exosomal release from cells [39,40]. Changes in the levels of circulating miRNA may occur due to other inflammatory reactions or the host immune response rather than only due to the intrinsic changes within the lesion [41]. Thus, as discussed further below, it is not surprising that miRNAs analyzed in tissue biopsies poorly correlate with those found in the circulation [42].
There is an increasing interest in exploring epigenetic mechanisms in common diseases, with notable progress in characterizing the contribution of miRNAs [43]. In their 2008 study, Wu et al. found that miRNAs regulate colonic epithelial cell-derived chemokine expression and were the first to relate miRNAs to IBD [44]. The field of miRNA research has grown rapidly after their discovery in human disease biology including in IBD [43]. We have listed a series of IBD-related miRNA studies from recent years in Table 1, with a focus on sample type and quantitative method. MiR-21, miR-155, and miR-31 have repeatedly been identified and seem to be the most studied miRNAs related to IBD [15,19,35,45,46,47,48]. MiR-21 is possibly the most intriguing miRNA involved in IBD, with associations between miR-21 and IBD being replicated in several studies, as well as functional relevance reported in mouse models of IBD [19,23,24,30,35,49]. Each miRNA can potentially target hundreds of mRNAs resulting in mRNA destabilization and/or inhibition of translation, however, restricted to a specific cellular context, the number of relevant targetable transcripts is probably quite low.

Table 1.
A summary of studies on microRNA research in inflammatory bowel disease (IBD). CD: Crohn’s disease, UC: Ulcerative colitis, HC: Healthy controls, RT-qPCR: Quantitative real time polymerase chain reaction, Biopsy: colon tissue biopsy, ISH: In situ hybridization, QISH: Quantitative in-situ hybridization, PBMC: Peripheral blood mononuclear cells, DSS: Dextran sodium sulphate, AOM: Azoxymethane, TNF: Tumor necrosis factor alpha.
MiRNAs regulate important cellular functions such as cell differentiation and proliferation and signal transduction and apoptosis and exhibit highly specific regulated patterns of gene expression [15]. In autoimmune diseases, miRNAs can act through interference with inflammatory signaling pathways, such as the nuclear transcription factor kappa B (NF-κB) pathway, IL23/IL23R pathway, and IL-6/STAT3 pathway [50,51,52,53,54]. Studying the RhoB pathway of cell adhesion in UC mucosa and cultured colon cancer cells, Yang et al. [36] examined the role of miR-21 in regulation of intestinal epithelial barrier function and found that miR-21 induced the degradation of RhoB mRNA, reduction in RhoB protein, causing loss of tight junctions in intestinal epithelial cells. Tian et al. showed miR-31 to be highly expressed in tissues from IBD patients, and miR-31 reduced the inflammatory response in the Dextran Sodium Sulphate (DSS)-induced colitis mouse model (see below), by preventing the expression of inflammatory cytokine receptors such as IL7R and IL17RA and signaling proteins such as GP130 [55]. Another study based on the DSS model showed that miR-155 directly binds to SHIP-1 mRNA and causes a significant decrease in SHIP-1 levels, which regulate cell membrane trafficking, and thereby contribute to the pathogenesis of colitis [56]. Taken together, these examples indicate the complexity of how miRNAs may act through signaling pathways in the biological settings of IBD.
Studies of circulating miRNAs have shown that miRNAs are potential candidates as biomarkers for diagnosing IBD and various other diseases [57,58,59,60,61]. The high stability of miRNAs in the body fluids and the ability to obtain rapid and accurate quantitative estimates are some merits of using circulating miRNAs as biomarkers in IBD [28]. MiRNAs are not only interesting tools for diagnosis, but also for potential future therapeutic applications by miRNA mimics or miRNA antagonists [62,63].
To study the pathogenesis and intricacy of IBD, the advancement of a variety of animal models has provided important information. The most extensively used mouse model of colitis utilizes DSS, a so-called chemical colitogen with anticoagulant properties, to stimulate epithelial damage. The DSS colitis model is simple and easy to administer. Acute and persistent colitis is achieved by altering the concentration of DSS and the frequency of administration [101]. A genetically engineered in vivo model that has been widely used to examine IBD etiology is the interleukin-10 (IL-10)-deficient mouse model [102]. IL-10 is an anti-inflammatory cytokine. Mutated IL-10 signaling systems shows early and aggressive expansion of systemic inflammation in IBD. IL-10 knockout mice develop spontaneous colitis and CAC [103]. Nata et al. [94] performed miRNA microarray profiling on IL-10-deficient mice and identified that several miRNAs were upregulated, including miR-146b that, through further studies, was found to contribute to increased intestinal inflammation by upregulating NF-κB. Shi et al. [95] showed that knockout of miR-21 in mice improved the survival rate in DSS-induced fatal colitis via protecting against inflammation and tissue injury. Hence, it was suggested that impaired expression of miR-21 in gut may block the onset or progression of IBD. Other animal models used in IBD research include genetically engineered mice, congenic mouse strains, chemically induced models, and cell-transfer models [104]. Most of the studies investigating miRNA expression in IBD have used high-throughput methods such as a microarray combined with RT-qPCR as a validation method for prioritized miRNAs.
4. MiRNA Biomarkers for IBD Diagnosis
The diagnostic assessment of IBD can be challenging; particularly, discriminating CD from UC can be a diagnostic encounter in cases where the inflammatory lesions are limited to the colon. It is estimated that 10–15% of IBD cases are categorized as IBDU [105]. Although many IBDU patients are eventually reclassified as either CD or UC, approximately 75% of the IBDU cases maintain the diagnosis of IBDU, suggesting that most of the IBDU patients have a distinct diagnostic entity of a true overlap phenotype between large bowel CD and typical UC [106,107]. Recently, a study that examined colon biopsies from patients with IBD suggested miR-19a, miR-21, miR-31, miR-146a and miR-375 as a biomarker profile for discriminating CD and UC [108]. A Study by Peck et al. used a next-generation sequencing–based approach and found that a combination of miR-31-5p, miR-215, miR-223-3p, miR-196b-5p and miR-203 could stratify patients with CD according to disease behavior independent of the effect of inflammation [109]. The lack of reproducibility in miRNA profiling analyses of IBD samples in independent studies could be due to the technology applied, as well as the variation in control groups, disease activity and data normalization. It was recently reported that miR-21 is a potential diagnostic marker for discriminating CD from UC, as both RT-qPCR and quantitative ISH (qISH) identified significantly higher levels in UC compared with CD [35]. The authors suggested that miR-21 is not just an unspecific marker of inflammation, but that miR-21 is specific to the immunopathological process of UC. miR-21 ISH analyses reveal complex expression patterns, where the miR-21 staining is identified mainly in cells of the inflamed lamina propria as well as in subsets of epithelial cells of partly damaged crypt structures (example in Figure 1).
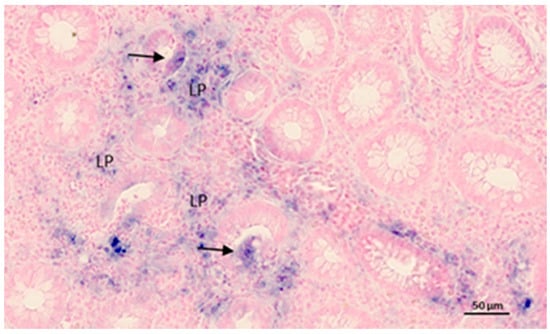
Figure 1.
MiR-21 in situ hybridization in ulcerative colitis. The example shows the inflamed colon mucosa with transversally cut crypts and the lamina propria (indicated by LP). The miR-21 ISH signal is represented by the blue stain and is seen in inflammatory cells located in the lamina propria in and some of the epithelial cells (arrows) in some collapsed crypts. Nuclear Fast Red was used in counterstaining.
5. MiRNAs and CAC
Chronic inflammation is linked to the development of a variety of cancers such as CRC, pancreatic, breast, and skin cancer [110,111] and is a key hallmark of cancer [112]. Local chronic inflammation in the colon, typically caused by an unbalance in the regulation of the immune response, may damage the epithelial barrier, which induces self-sustained inflammation linked to continued microbial influx or increased levels of pro-inflammatory cytokines like tumor necrosis factor alpha (TNF) and IL-1β [110,113,114]. Increased levels of oxidants in inflamed tissue cause cell death, or more deliberately, mutations in epithelial cells that, in turn, can initiate neoplastic growth. The persisting inflammation develops into and probably shapes the tumor microenvironment that is inherent to most solid tumors with their additional presence of blood vessels and fibroblastic cells.
Patients with extensive IBD or diagnosed with IBD in childhood [115], have a shorter life expectancy that may be related to the higher risk of CRC [116,117,118]. CAC represents a type of CRC in which the IBD paved the way for the cancer, probably through mutations in K-RAS and the adenomatous polyposis coli (APC) gene [119]. The risk of CAC may be further increased in untreated IBD patients [120]. Mucosal mapping studies indicate that the chronically inflamed colonic mucosa of patients with IBD undergoes a “field change” in cancer-associated molecular alterations before there is histologic evidence of epithelial dysplasia [121,122], which is one the initial morphological changes in the stepwise progression to CRC [123,124,125].
MiRNAs are believed to take part in the inflammation in IBD and to be implicated in the process from inflammation to CRC [126]. Despite the fact that CD and UC can affect the entire colon, Ranjha et al. [127] found that CRC in UC patients developed primarily in the rectosigmoid areas of the colon, whereas other parts, such as the ascending colon, showed less frequent development of CRC. Analyzing tissue from rectosigmoid and ascending colon, the authors found differences in the miRNA expression patterns, and suggested that the local miRNA profile could contribute to the development of CRC.
MiRNAs likely play both oncogenic and tumor-suppressive roles in the carcinogenesis and progression of CRC by regulating the expression of numerous cancer-related genes. The role of the inflammatory burden has also been studied in animal models and indicates that both the initiation and the progression of colonic neoplasia can be aggravated or accelerated by the inflammatory conditions [30,126,128,129,130]. The DSS-induced colitis model has been used to study the role of multiple miRNAs in IBD and CAC, including miR-21, miR-155, and miR-301a, which will be addressed briefly in the following.
MiR-21 is one of the most prevalent miRNAs in CRC and other cancer types [131,132], and the increased expression levels in CRC are associated with poor prognosis [24,133]. MiR-21 acts on tumor-suppressor genes, like PTEN and PDCD4 [134,135,136], and is thus categorized as an oncomiR [137]. Since miR-21 is upregulated in IBD [35,44,47], and miR-21 reduction in the DSS model lowers inflammation in DSS-induced colitis [95], it is tempting to speculate that miR-21 is a key facilitator of CAC. In support of this hypothesis, a study of human IBD, found that the tumor-suppressor-programmed cell death 4 (PDCD4) was downregulated, while miR-21 was upregulated [134]. Suppression of PDCD4 and NF-κB activation was found along with reduced levels of pro-inflammatory TNF [134,135]. In addition, epithelial miR-21 upregulation in UC was reported to increase intestinal permeability, which is believed to be a key pathophysiological step in the development of IBD [138].
MiR-155 is upregulated in both UC and CD patients compared to healthy controls [48,139,140,141] and is upregulated in both tissue and blood from CRC patients, and is furthermore an indicator of poor prognosis [142,143]. MiR-155 promotes intestinal inflammation in UC and CD, probably via a variety of inflammation-related pathways [46,56,73,79,81,139]. In a recent study by Liu et al. [144,145], it was shown that miR-155 mediates intestinal barrier dysfunction in DSS-induced mice colitis through targeting the HIF-1α/TFF-3 axis. Paraskevi et al. [140] found that miR-155 is the most highly expressed UC-associated miRNA in blood samples, however, in the study by Schönauen [47], the authors did not find increased miR-155 levels in the blood from IBD patients, suggesting that more studies are needed to determine whether miR-155 is a putative blood-related biomarker.
MiR-301a is upregulated in both blood and tissue from IBD and CRC patients [76,141,146]. He et al. [76,129] found increased levels of miR-301a in peripheral blood monocytes and in the mucosa from IBD patients and in mice after administration of DSS. Using the DSS-induced IBD model in mice with an inactivated miR-301a, miR-301a was found to reduce the inflammation through the suppression of BTG anti-proliferation factor 1 (BTG1) and to reduce the development of CAC [129]. Thus, miR-301a should be investigated in future studies to establish possible use as a clinically relevant diagnostic biomarker in IBD and for prediction of CAC.
6. MiRNAs as Predictive Biomarkers and in IBD Treatment
The goal of the treatment of IBD patients is to obtain remission and mucosal healing, and thereby lower surgery rates. The classical therapies include corticosteroids, thiopurines, and amino salicylates (5-ASA), which have been in use for decades. 5-ASA has minor side effects [147,148] and it is very effective for treating mild to moderate UC patients, but not recommended for treatment of patients with CD [149]. The last-line medical treatment in IBD is administration of biologics targeting key elements in the inflammatory process. Anti-TNF therapies include TNF inhibitors that antagonize the pro-inflammatory cytokine TNF [150]. The use of anti-TNF therapy has improved long-term outcomes for IBD patients [149,151]. Even though TNF inhibitors have improved the overall conditions for a large group of IBD patients, approximately 30% of patients fail to respond to TNF inhibitors (primary non-responders), and up to 50% of the patients who initially benefited from treatment with TNF inhibitors lose the response over time (secondary non-responders) [152,153]. Thus, identifying predictors of responders/non-responders and choosing a treatment strategy according to biomarker profiles could improve overall IBD disease management. Interestingly, Morilla et al. [154], found that nine miRNAs, together with five clinical factors correlated with response to treatment of IBD patients, and that neural-network-developed algorithms based on certain miRNA levels identified responders to the anti-TNF antibody therapy, infliximab, vs. non-responders.
Currently used therapies in IBD also include Ustekinumab, Vedolizumab and Tofacitinib. Ustekinumab is a monoclonal antibody against IL-12 and IL-23, which is used in patients with moderate to severe CD who are resistant to anti-TNF treatment [155]. Considering the efficacy of ustekinumab, it is possible to extrapolate the efficacy of miR-29 mimicry as a mechanism to reduce IL-23 levels [12]. With respect to potential secondary target effects, miR-29c has been described as a tumor-suppressor in liver cancer [156]. Vedolizumab binds specifically to α4β7-integrin on T-helper lymphocytes. Blocking the α4β7-integrin results in anti-inflammatory activity that is caused by the inhibition of leukocyte adhesion to endothelial cells, which consequently reduces leukocyte recruitment to affected tissue [157]. Previous studies have suggested a similar effect of miRNAs in the posttranscriptional regulation of leukocyte trafficking [158]. Harris et al. [158] described how endogenous miR-126 inhibits leukocyte adherence through the regulation of an intercellular adhesion molecule expressed by endothelial cells (VCAM-1). Tofacitinib is a janus kinase (JAK) inhibitor, approved for treating moderate-to-severely active UC patients who have deteriorated disease and did not improve after conventional or antibody-based therapies [159,160]. Pathak et al. [51] identified SOCS1, a potent molecular switch that tunes the JAK pathway that is also a direct target of miR-155.
In general, miRNA-based therapies comprise two fundamental strategies: miRNA antagonism and mimicry [161,162,163,164]. Physiologic miRNA over-expression resulting in pathologically reduced target gene expression can be hindered by using miRNA antagonists, while reduced miRNA expression resulting in enhanced target function can be restored by utilizing miRNA mimics [12]. A study by Lu et al. [56] reported that a so-called antagomir towards miR-155 alleviated DSS-induced intestinal inflammation in mice, and the authors propose that anti-miR-155 could be a promising candidate for a novel IBD therapy. Jin et al. conducted a study on miR-133a and its target UCP2 (mitochondrial uncoupling protein 2) using the DSS-induced IBD mouse model [77]. miR-133a levels were found to be decreased upon DSS treatment, and by introducing a miR-133a mimic, the DSS-induced IBD was alleviated, suggesting that miRNA mimics could also function as therapy in IBD [77].
7. Circulating miRNAs vs. Tissue miRNAs
It is of importance to determine whether miRNA dysregulation in the circulation reflects similar changes in the lesion. The detection and quantification of circulating miRNAs and the interpretation of their impending role as novel non-invasive biomarkers could be very beneficial in the diagnosis and treatment of IBD. As mentioned above, miRNAs can be detected in distinct body fluids such as saliva, plasma or urine [32]. Current diagnostic and predictive findings in IBD on miRNA expression profiling have mainly focused on the assessment of miRNAs in blood. Even though blood samples can be relatively easily obtained from IBD patients, miRNA measurement in blood samples, as with other biological samples, comes with some inherent obstacles, such as sample procurement, storage, measurement platform and normalization of the acquired data. Circulating miRNAs may derive from both the diseased tissue and by leakage from the normal vascular network and circulating cells. Obtaining tissue samples, on the other hand, requires an invasive procedure, where small biopsies from the affected part of the bowel are obtained during endoscopy. The tissue samples can be either frozen or fixed in formalin and paraffin-embedded (FFPE) for histological examination. MiRNAs can be isolated from both fresh-frozen and FFPE tissue samples. Normalization of miRNA data from both blood and tissue samples is an important step for data interpretation in the comparison between patients, and between different study cohorts. MiRNA expression levels measured in tissue samples will have been derived from cells in the normal tissue and from activated cells in the lesion. To be able to find the same miRNAs in the tissue as in the circulation would require substantial expression in the lesion and/or for the background level in the circulation to be low. Thus, it may not be surprising that the study by Iborra et al. [42] of tissue biopsies and peripheral blood showed that none of the serum miRNAs corresponded with tissue miRNAs in the CD and UC patients. Feces samples represent another liquid biopsy that is relevant in relation to IBD and may be better linked to expression levels in the diseased mucosa than to the levels in the blood circulation. Schönauen et al. [47] analyzed both serum and fecal miRNAs in IBD and found increased levels of miR-16, miR-21, and miR-223 in both sera and feces from the IBD patients compared to controls. In addition, the authors found that fecal levels, but not sera levels, of miR-16 and miR-223 correlated with clinical parameters, like C-reactive protein and calprotectin. Thus, fecal samples seem to be a promising alternative to blood for miRNA profiling in IBD.
As seen in Table 1, most miRNA studies have used high-throughput methods such as RT-qPCR and microarray for miRNA analysis in IBD. It is important to note that these techniques require homogenization of the tissue to isolate the miRNAs. Homogenization of the tissue will degrade the spatial arrangement and, hence, will give an overview of the miRNA expression at the tissue level. ISH using Locked Nucleic Acid (LNA) probes is a method that allows detection of miRNAs in tissue sections [165]. Detection of miRNAs at the cellular level determines the cellular origin of expression and can provide evidence on expression levels in different cell populations and tissue compartments [35]. More knowledge about the cellular localization of miRNAs in the framework of IBD is needed as this will provide a vital link between the growing amounts of miRNA biomarkers discovered in IBD and functional studies identifying various miRNA target genes. Thorlacius-Ussing et al. used quantitative ISH on IBD tissue samples and showed that miR-126 levels are increased in UC and expressed in endothelial cells and miR-21 is expressed in subsets of monocytes/macrophages and T cells [35]. As also suggested from Figure 1, ISH data provide information of contextual expression in the tissue, as exemplified by focal upregulation in certain tissue compartments. Simple histological analysis from ISH analysis if IBD tissue can often determine if a miRNA is expressed in the epithelial or stromal (lamina propria) compartment. Nielsen and Holmstrøm presented a method to combine miRNA ISH using LNA-containing probes with immunohistochemical detection of cell-specific protein markers in order to better characterize the miRNA’s cellular characteristics [166]. This approach could also be used to monitor parallel downregulation of the specific downstream target protein. MiRNA ISH is a powerful tool when also combined with parallel characterization of the cell population in question and of mRNAs using combined staining methods [23]. Thus, for better understanding of the role of miRNAs in IBD and CAC, miRNA ISH analyses will be a helpful tool both for validating expression and for deciphering the related inflammatory molecular context.
8. Concluding Remarks
MiRNAs in IBD research started with the extensive pioneering work by Wu et al. in 2008 [44], who found altered expression of several miRNAs in tissue from IBD patients. Since then, there have been tremendous advancements in the field both regarding mechanistic studies and studies evaluating the use of miRNAs as diagnostic and predictive biomarkers in IBD. The miRNAs are involved in the regulation of the NF-κB and the IL-6 pathways, regulating the inflammatory activity. The inflammation is fueled by cytokines like TNF, which is currently a key therapeutic target. Thus, the dysregulated miRNAs may be considered also as therapeutic targets in IBD. Tracking the immune status in IBD based on miRNA signatures determined from liquid or tissue biopsies, may be powerful for designing individualized therapies that could be, e.g., combinations of conventional drugs and biologically active drugs, like anti-TNF. In this review, we discussed the possibility of using miRNA expression profiles to understand the link between inflammation in IBD and CAC, where animal models of IBD have provided new information on the role of miRNAs both as biomarkers and as possible therapeutic targets. Future studies may apply new sequencing techniques and histology-based multiplexing analyses in well-annotated independent patient cohorts to address the possible value of miRNAs as diagnostic and predictive biomarkers.
Author Contributions
Conceptualization, J.P.J., B.S.N., L.B.R. and E.H.; resources, J.P.J., B.S.N., L.B.R., E.H., M.M. and E.L.; data curation, J.P.J., B.S.N., L.B.R., E.H., M.M. and E.L.; writing—original draft preparation, J.P.J. and B.S.N.; writing—review and editing, J.P.J., B.S.N., L.B.R., E.H., M.M. and E.L.; supervision, B.S.N., L.B.R., E.H., M.M. and E.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Souza, H.S.P.; Fiocchi, C. Immunopathogenesis of IBD: Current state of the art. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Bouma, G.; Strober, W. The immunological and genetic basis of inflammatory bowel disease. Nat. Rev. Immunol. 2003, 3, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, B.A.; Gokhale, R.; Cho, J.H. Clinical aspects and pathophysiology of inflammatory bowel disease. Clin. Microbiol. Rev. 2002, 15, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Shen, B. Overview of Diagnosis and Medical Treatment of Inflammatory Bowel Diseases; Academic Press: New York, NY, USA, 2018; ISBN 9780128113882. [Google Scholar]
- Kappelman, M.D.; Grand, R.J. Does inflammatory bowel disease develop in infants? Inflamm. Bowel Dis. 2008, 14 (Suppl. 2), S6–S8. [Google Scholar] [CrossRef] [PubMed]
- Molodecky, N.A.; Soon, I.S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W.; et al. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology 2012, 142, 46–54.e42. [Google Scholar] [CrossRef]
- Burisch, J.; Jess, T.; Martinato, M.; Lakatos, P.L. The burden of inflammatory bowel disease in Europe. J. Crohn’s Colitis 2013, 7, 322–337. [Google Scholar] [CrossRef]
- Lophaven, S.N.; Lynge, E.; Burisch, J. The incidence of inflammatory bowel disease in Denmark 1980–2013: A nationwide cohort study. Aliment. Pharmacol. Ther. 2017, 45, 961–972. [Google Scholar] [CrossRef]
- Hendriksen, C.; Kreiner, S.; Binder, V. Long term prognosis in ulcerative colitis—Based on results from a regional patient group from the county of Copenhagen. Gut 1985, 26, 158–163. [Google Scholar] [CrossRef]
- Loftus, E.V., Jr. Clinical epidemiology of inflammatory bowel disease: Incidence, prevalence, and environmental influences. Gastroenterology 2004, 126, 1504–1517. [Google Scholar] [CrossRef]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef]
- Chapman, C.G.; Pekow, J. The emerging role of miRNAs in inflammatory bowel disease: A review. Therap. Adv. Gastroenterol. 2015, 8, 4–22. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, J.S. MicroRNAs: How many in inflammatory bowel disease? Curr. Opin. Gastroenterol. 2016, 32, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, J. microRNAs as therapeutic targets in intestinal diseases. ExRNA 2019, 1, 1–12. [Google Scholar] [CrossRef]
- Ambros, V. microRNAs: Tiny Regulators with Great Potential. Cell 2001, 107, 823–826. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans Heterochronic Gene lin-4 Encodes Small RNAs with Antisense Complementarity to &II-14 Rosalind. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function Review. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef]
- Zarjou, A.; Yang, S.; Abraham, E.; Agarwal, A.; Liu, G. Identification of a microRNA signature in renal fibrosis: Role of miR-21. AJP Ren. Physiol. 2011, 301, F793–F801. [Google Scholar] [CrossRef]
- Agarwal, S.; Hanna, J.; Sherman, M.E.; Figueroa, J.; Rimm, D.L. Quantitative assessment of miR34a as an independent prognostic marker in breast cancer. Br. J. Cancer 2015, 112, 61–68. [Google Scholar] [CrossRef][Green Version]
- Xuan, Y.; Yang, H.; Zhao, L.; Bond Lau, W.; Bonnie, L.; Ning, R.; Yuehong, H.; Tao, Y.; Xia, Z.; Shengtao, Z.; et al. MicroRNAs in colorectal cancer: Small molecules with big functions. Cancer Lett. 2015, 360, 89–105. [Google Scholar] [CrossRef]
- Nielsen, B.S.; Balslev, E.; Poulsen, T.; Nielsen, D.; Møller, T.; Mortensen, C.E.; Holmstrøm, K.; Høgdall, E. miR-21 expression in cancer cells may not predict resistance to adjuvant trastuzumab in primary breast cancer. Front. Oncol. 2014, 4, 207. [Google Scholar] [CrossRef] [PubMed]
- Møller, T.; James, J.P.; Holmstrøm, K.; Sørensen, F.B.; Lindebjerg, J.; Nielsen, B.S. Co-Detection of miR-21 and TNF-α mRNA in Budding Cancer Cells in Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 1907. [Google Scholar] [CrossRef] [PubMed]
- Kjaer-Frifeldt, S.; Hansen, T.F.; Nielsen, B.S.; Joergensen, S.; Lindebjerg, J.; Soerensen, F.B.; dePont Christensen, R.; Jakobsen, A.; Group, D.C.C. The prognostic importance of miR-21 in stage II colon cancer: A population-based study. Br. J. Cancer 2012, 107, 1169–1174. [Google Scholar] [CrossRef]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef]
- Galimberti, D.; Villa, C.; Fenoglio, C.; Serpente, M.; Ghezzi, L.; Cioffi, S.M.G.; Arighi, A.; Fumagalli, G.; Scarpini, E. Circulating miRNAs as potential biomarkers in alzheimer’s disease. J. Alzheimer’s Dis. 2014, 42, 1261–1267. [Google Scholar] [CrossRef]
- Correia, C.N.; Nalpas, N.C.; McLoughlin, K.E.; Browne, J.A.; Gordon, S.V.; MacHugh, D.E.; Shaughnessy, R.G. Circulating microRNAs as potential biomarkers of infectious disease. Front. Immunol. 2017, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Alamdari-palangi, V.; Vahedi, F.; Shabaninejad, Z.; Dokeneheifard, S.; Movehedpour, A.; Taheri-Anganeh, M.; Savardashtaki, A. microRNA in inflammatory bowel disease at a glance. Eur. J. Gastroenterol. Hepatol. 2020, 1–9. [Google Scholar] [CrossRef]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A.O.; Landthaler, M.; et al. A Mammalian microRNA Expression Atlas Based on Small RNA Library Sequencing. Cell 2007, 129, 1401–1414. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, Y.; Zhou, D.; Chen, G.; Li, N. MicroRNAs, intestinal inflammatory and tumor. Bioorg. Med. Chem. Lett. 2019, 29, 2051–2058. [Google Scholar] [CrossRef]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed]
- Tölle, A.; Jung, M.; Rabenhorst, S.; Kilic, E.; Jung, K.; Weikert, S. Identification of microRNAs in blood and urine as tumour markers for the detection of urinary bladder cancer. Oncol. Rep. 2013, 30, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shachar, S.; Yanai, H.; Horev, H.S.; Elad, H.; Baram, L.; Issakov, O.; Tulchinsky, H.; Pasmanik-Chor, M.; Shomron, N.; Dotan, I. MicroRNAs expression in the ileal pouch of patients with ulcerative colitis is robustly up-regulated and correlates with disease phenotypes. PLoS ONE 2016, 11, e159956. [Google Scholar] [CrossRef]
- Thorlacius-Ussing, G.; Schnack Nielsen, B.; Andersen, V.; Holmstrøm, K.; Pedersen, A.E. Expression and Localization of miR-21 and miR-126 in Mucosal Tissue from Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2017, 23, 739–752. [Google Scholar] [CrossRef]
- Yang, Y.; Ma, Y.; Shi, C.; Chen, H.; Zhang, H.; Chen, N.; Zhang, P.; Wang, F.; Yang, J.; Yang, J.; et al. Overexpression of miR-21 in patients with ulcerative colitis impairs intestinal epithelial barrier function through targeting the Rho GTPase RhoB. Biochem. Biophys. Res. Commun. 2013, 434, 746–752. [Google Scholar] [CrossRef]
- Nagy, Z.B.; Barták, B.K.; Kalmár, A.; Galamb, O.; Wichmann, B.; Dank, M.; Igaz, P.; Tulassay, Z.; Molnár, B. Comparison of Circulating miRNAs Expression Alterations in Matched Tissue and Plasma Samples During Colorectal Cancer Progression. Pathol. Oncol. Res. 2019, 25, 97–105. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—MicroRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Secreto, C.R.; Knox, T.R.; Ding, W.; Mukhopadhyay, D.; Kay, N.E. Circulating microvesicles in B-cell chronic lymphocytic leukemia can stimulate marrow stromal cells: Implications for disease progression. Blood 2010, 115, 1755–1764. [Google Scholar] [CrossRef]
- Lima, L.G.; Chammas, R.; Monteiro, R.Q.; Moreira, M.E.C.; Barcinski, M.A. Tumor-derived microvesicles modulate the establishment of metastatic melanoma in a phosphatidylserine-dependent manner. Cancer Lett. 2009, 283, 168–175. [Google Scholar] [CrossRef]
- Waters, P.S.; McDermott, A.M.; Wall, D.; Heneghan, H.M.; Miller, N.; Newell, J.; Kerin, M.J.; Dwyer, R.M. Relationship between Circulating and Tissue microRNAs in a Murine Model of Breast Cancer. PLoS ONE 2012, 7, e50459. [Google Scholar] [CrossRef] [PubMed]
- Iborra, M.; Bernuzzi, F.; Correale, C.; Vetrano, S.; Fiorino, G.; Beltrán, B.; Marabita, F.; Locati, M.; Spinelli, A.; Nos, P.; et al. Identification of serum and tissue micro-RNA expression profiles in different stages of inflammatory bowel disease. Clin. Exp. Immunol. 2013, 173, 250–258. [Google Scholar] [CrossRef]
- Kalla, R.; Ventham, N.T.; Kennedy, N.A.; Quintana, J.F.; Nimmo, E.R.; Buck, A.H.; Satsangi, J. MicroRNAs: New players in IBD. Gut 2015, 64, 504–517. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zikusoka, M.; Trindade, A.; Dassopoulos, T.; Harris, M.L.; Bayless, T.M.; Brant, S.R.; Chakravarti, S.; Kwon, J.H. MicroRNAs Are Differentially Expressed in Ulcerative Colitis and Alter Expression of Macrophage Inflammatory Peptide-2α. Gastroenterology 2008, 135, 1624–1635.e24. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Kelly, O.B.; Smith, M.I.; Kabakchiev, B.; Silverberg, M.S. Differential miRNA expression in ileal and colonic tissues reveals an altered immunoregulatory molecular profile in individuals with Crohn’s disease versus healthy subjects. J. Crohn’s Colitis 2019, 13, 1459–1469. [Google Scholar] [CrossRef]
- Béres, N.J.; Szabó, D.; Kocsis, D.; Szucs, D.; Kiss, Z.; Müller, K.E.; Lendvai, G.; Kiss, A.; Arató, A.; Sziksz, E.; et al. Role of Altered Expression of MIR-146a, MIR-155, and MIR-122 in Pediatric Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2016. [Google Scholar] [CrossRef]
- Schönauen, K.; Le, N.; Von Arnim, U.; Schulz, C.; Malfertheiner, P.; Link, A. Circulating and Fecal microRNAs as Biomarkers for Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2018, 24, 1547–1557. [Google Scholar] [CrossRef] [PubMed]
- Valmiki, S.; Ahuja, V.; Paul, J. MicroRNA exhibit altered expression in the inflamed colonic mucosa of ulcerative colitis patients. World J. Gastroenterol. 2017, 23, 5324. [Google Scholar] [CrossRef]
- Feng, Y.; Tsao, C. Emerging role of microRNA-21 in cancer (Review). Biomed. Rep. 2016, 5, 395–402. [Google Scholar] [CrossRef]
- Chen, W.-X.; Ren, L.-H.; Shi, R.-H. Implication of miRNAs for inflammatory bowel disease treatment: Systematic review. World J. Gastrointest. Pathophysiol. 2014, 5, 63. [Google Scholar] [CrossRef]
- Pathak, S.; Grillo, A.R.; Scarpa, M.; Brun, P.; D’Incà, R.; Nai, L.; Banerjee, A.; Cavallo, D.; Barzon, L.; Palù, G.; et al. MiR-155 modulates the inflammatory phenotype of intestinal myofibroblasts by targeting SOCS1 in ulcerative colitis. Exp. Mol. Med. 2015, 47, e164. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Kwon, H.Y.; Ha Thi, H.T.; Lee, H.J.; Kim, G.Il.; Hahm, K.-B.; Hong, S. MicroRNA-132 and microRNA-223 control positive feedback circuit by regulating FOXO3a in inflammatory bowel disease. J. Gastroenterol. Hepatol. 2016, 31, 1727–1735. [Google Scholar] [CrossRef] [PubMed]
- Pierdomenico, M.; Cesi, V.; Cucchiara, S.; Vitali, R.; Prete, E.; Costanzo, M.; Aloi, M.; Oliva, S.; Stronati, L. NOD2 Is Regulated By Mir-320 in Physiological Conditions but this Control Is Altered in Inflamed Tissues of Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2016, 22, 315–326. [Google Scholar] [CrossRef]
- Moein, S.; Vaghari-Tabari, M.; Qujeq, D.; Majidinia, M.; Nabavi, S.M.; Yousefi, B. MiRNAs and inflammatory bowel disease: An interesting new story. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef]
- Tian, Y.; Xu, J.; Li, Y.; Zhao, R.; Du, S.; Lv, C.; Wu, W.; Liu, R.; Sheng, X.; Song, Y.; et al. MicroRNA-31 Reduces Inflammatory Signaling and Promotes Regeneration in Colon Epithelium, and Delivery of Mimics in Microspheres Reduces Colitis in Mice. Gastroenterology 2019, 156, 2281–2296.e6. [Google Scholar] [CrossRef]
- Lu, Z.J.; Wu, J.J.; Jiang, W.L.; Xiao, J.H.; Tao, K.Z.; Ma, L.; Zheng, P.; Wan, R.; Wang, X.P. MicroRNA-155 promotes the pathogenesis of experimental colitis by repressing SHIP-1 expression. World J. Gastroenterol. 2017, 23, 976–985. [Google Scholar] [CrossRef]
- Oliveira, D.N.P.; Carlsen, A.L.; Heegaard, N.H.H.; Prahm, K.P.; Christensen, I.J.; Høgdall, C.K.; Høgdall, E.V. Diagnostic plasma miRNA-profiles for ovarian cancer in patients with pelvic mass. PLoS ONE 2019, 14, e225249. [Google Scholar] [CrossRef]
- Thakral, S.; Ghoshal, K. miR-122 is a unique molecule with great potential in diagnosis, prognosis of liver disease, and therapy both as miRNA mimic and antimir. Curr. Gene Ther. 2015, 15, 142–150. [Google Scholar] [CrossRef]
- Ma, J.; Lin, Y.; Zhan, M.; Mann, D.L.; Stass, S.A.; Jiang, F. Differential miRNA expressions in peripheral blood mononuclear cells for diagnosis of lung cancer. HHS Public Interes. 2016, 95, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Coskun, M.; Bjerrum, J.T.; Seidelin, J.B.; Nielsen, O.H. MicroRNAs in inflammatory bowel disease—Pathogenesis, diagnostics and therapeutics. World J. Gastroenterol. 2012, 18, 4629–4634. [Google Scholar] [CrossRef]
- El-Khoury, V.; Pierson, S.; Kaoma, T.; Bernardin, F.; Berchem, G. Assessing cellular and circulating miRNA recovery: The impact of the RNA isolation method and the quantity of input material. Sci. Rep. 2015, 6, 19529. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Cao, D.L.; Zhao, L.X.; Han, Y.; Zhang, Y.L. MicroRNA-146a-5p attenuates visceral hypersensitivity through targeting chemokine CCL8 in the spinal cord in a mouse model of colitis. Brain Res. Bull. 2018, 139, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, S.; Qiu, Y.; He, Y.; Chen, B.; Mao, R.; Cui, Y.; Zeng, Z.; Chen, M. Upregulation of miR-665 promotes apoptosis and colitis in inflammatory bowel disease by repressing the endoplasmic reticulum stress components XBP1 and ORMDL3. Cell Death Dis. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Nijhuis, A.; Curciarello, R.; Mehta, S.; Feakins, R.; Bishop, C.L.; Lindsay, J.O.; Silver, A. MCL-1 is modulated in Crohn’s disease fibrosis by miR-29b via IL-6 and IL-8. Cell Tissue Res. 2017, 368, 325–335. [Google Scholar] [CrossRef]
- Zidar, N.; Boštjančič, E.; Jerala, M.; Kojc, N.; Drobne, D.; Štabuc, B.; Glavač, D. Down-regulation of microRNAs of the miR-200 family and up-regulation of Snail and Slug in inflammatory bowel diseases—Hallmark of epithelial−mesenchymal transition. J. Cell. Mol. Med. 2016, 20, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Chen, S.; Hu, W. MicroRNA-141 Is Involved in Ulcerative Colitis Pathogenesis via Aiming at CXCL5. J. Interferon Cytokine Res. 2017, 37, 415–420. [Google Scholar] [CrossRef]
- Koukos, G.; Polytarchou, C.; Kaplan, J.L.; Morley-Fletcher, A.; Gras-Miralles, B.; Kokkotou, E.; Baril-Dore, M.; Pothoulakis, C.; Winter, H.S.; Iliopoulos, D. MicroRNA-124 regulates STAT3 expression and is down-regulated in colon tissues of pediatric patients with ulcerative colitis. Gastroenterology 2013, 145, 842. [Google Scholar] [CrossRef]
- Cheng, X.; Zhang, X.; Su, J.; Zhang, Y.; Zhou, W.; Zhou, J.; Wang, C.; Liang, H.; Chen, X.; Shi, R.; et al. MiR-19b downregulates intestinal SOCS3 to reduce intestinal inflammation in Crohn’s disease. Sci. Rep. 2015, 5, 10397. [Google Scholar] [CrossRef]
- Yu, M.; Luo, Y.; Cong, Z.; Mu, Y.; Qiu, Y.; Zhong, M. MicroRNA-590-5p Inhibits Intestinal Inflammation by Targeting YAP. J. Crohn’s Colitis 2018, 12, 993–1004. [Google Scholar] [CrossRef]
- Bai, J.; Yu, J.; Wang, J.; Xue, B.; He, N.; Tian, Y.; Yang, L.; Wang, Y.; Wang, Y.; Tang, Q. DNA Methylation of miR-122 Aggravates Oxidative Stress in Colitis Targeting SELENBP1 Partially by p65NF-κB Signaling. Oxidative Med. Cell. Longev. 2019, 2019, 5294105. [Google Scholar] [CrossRef]
- Wu, W.; He, C.; Liu, C.; Cao, A.T.; Xue, X.; Evans-Marin, H.L.; Sun, M.; Fang, L.; Yao, S.; Pinchuk, I.V.; et al. miR-10a inhibits dendritic cell activation and Th1/Th17 cell immune responses in IBD. Gut 2015, 64, 1755–1764. [Google Scholar] [CrossRef]
- Tang, W.-J.; Peng, K.-Y.; Tang, Z.-F.; Wang, Y.-H.; Xue, A.-J.; Huang, Y. MicroRNA-15a—Cell division cycle 42 signaling pathway in pathogenesis of pediatric inflammatory bowel disease. World J. Gastroenterol. 2018, 24, 5234–5245. [Google Scholar] [CrossRef] [PubMed]
- Szcs, D.; Béres, N.J.; Rokonay, R.; Boros, K.; Borka, K.; Kiss, Z.; Arató, A.; Szabó, A.J.; Vannay, Á.; Sziksz, E.; et al. Increased duodenal expression of miR-146a and -155 in pediatric Crohn’s disease. World J. Gastroenterol. 2016, 22, 6027–6035. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Li, Y.; Li, L.; Yu, Q.; Chao, K.; Zhou, G.; Qiu, Y.; Feng, R.; Huang, S.; He, Y.; et al. Circulating microRNA146b-5p is superior to C-reactive protein as a novel biomarker for monitoring inflammatory bowel disease. Aliment. Pharmacol. Ther. 2019, 49, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; He, Q.; Guo, Z.; Xiong, F.; Li, Y.; Pan, Y.; Gao, C.; Li, L.; He, C. MicroRNA-425 facilitates pathogenic Th17 cell differentiation by targeting forkhead box O1 (Foxo1) and is associated with inflammatory bowel disease. Biochem. Biophys. Res. Commun. 2018, 496, 352–358. [Google Scholar] [CrossRef]
- He, C.; Shi, Y.; Wu, R.; Sun, M.; Fang, L.; Wu, W.; Liu, C.; Tang, M.; Li, Z.; Wang, P.; et al. MIR-301a promotes intestinal mucosal inflammation through induction of IL-17A and TNF-α in IBD. Gut 2016, 65, 1938–1950. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Chen, D.; Zheng, R.H.; Zhang, H.; Chen, Y.P.; Zun, X. MiRNA-133a-UCP2 pathway regulates inflammatory bowel disease progress by influencing inflammation, oxidative stress and energy metabolism. World J. Gastroenterol. 2017, 23, 76–86. [Google Scholar] [CrossRef]
- Coskun, M. miR-20b, miR-98, miR-125b-1*, and let-7e* as new potential diagnostic biomarkers in ulcerative colitis. World J. Gastroenterol. 2013, 19, 4289. [Google Scholar] [CrossRef]
- Zheng, Y.; Ge, W.; Ma, Y.; Xie, G.; Wang, W.; Han, L.; Bian, B.; Li, L.; Shen, L. miR-155 regulates IL-10-producing CD24hiCD27+ B cells and impairs their function in patients with Crohn’s disease. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Fang, K.; Law, I.K.M.; Padua, D.; Sideri, A.; Huang, V.; Kevil, C.G.; Iliopoulos, D.; Pothoulakis, C. MicroRNA-31-3p Is Involved in Substance P (SP)-Associated Inflammation in Human Colonic Epithelial Cells and Experimental Colitis. Am. J. Pathol. 2018, 188, 586–599. [Google Scholar] [CrossRef]
- Takagi, T.; Naito, Y.; Mizushima, K.; Hirata, I.; Yagi, N.; Tomatsuri, N.; Ando, T.; Oyamada, Y.; Isozaki, Y.; Hongo, H.; et al. Increased expression of microRNA in the inflamed colonic mucosa of patients with active ulcerative colitis. J. Gastroenterol. Hepatol. (Aust.) 2010, 25, S129–S133. [Google Scholar] [CrossRef]
- Zhang, H.; Li, W. MicroRNA-15 Activates NF-κB pathway via down regulating expression of adenosine A2 receptor in ulcerative colitis. Cell. Physiol. Biochem. 2018, 51, 1932–1944. [Google Scholar] [CrossRef] [PubMed]
- Pekow, J.R.; Dougherty, U.; Mustafi, R.; Zhu, H.; Kocherginsky, M.; Rubin, D.T.; Hanauer, S.B.; Hart, J.; Chang, E.B.; Fichera, A.; et al. MiR-143 and miR-145 are downregulated in ulcerative colitis: Putative regulators of inflammation and protooncogenes. Inflamm. Bowel Dis. 2012, 18, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Minacapelli, C.D.; Bajpai, M.; Geng, X.; Van Gurp, J.; Poplin, E.; Amenta, P.S.; Brant, S.R.; Das, K.M. miR-206 as a Biomarker for Response to Mesalamine Treatment in Ulcerative Colitis. Inflamm. Bowel Dis. 2019, 25. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Chen, X.; Chen, Q.; Shi, L.; Liang, H.; Zhou, Z.; Liu, Q.; Pang, W.; Hou, D.; Wang, C.; et al. MicroRNA-193a-3p reduces intestinal inflammation in response to microbiota via down-regulation of colonic PepT1. J. Biol. Chem. 2015, 290, 16099–16115. [Google Scholar] [CrossRef]
- Chen, B.; She, S.; Li, D.; Liu, Z.; Yang, X.; Zeng, Z.; Liu, F. Role of miR-19a targeting TNF-α in mediating ulcerative colitis. Scand. J. Gastroenterol. 2013, 48, 815–824. [Google Scholar] [CrossRef]
- Lu, X.; Yu, Y.; Tan, S. The role of the miR-21-5p-mediated inflammatory pathway in ulcerative colitis. Exp. Ther. Med. 2019, 19, 981–989. [Google Scholar] [CrossRef]
- Chen, Y.; Ge, W.; Xu, L.; Qu, C.; Zhu, M.; Zhang, W.; Xiao, Y. miR-200b is involved in intestinal fibrosis of Crohn’s disease. Int. J. Mol. Med. 2012, 29, 601–606. [Google Scholar] [CrossRef]
- Olaru, A.V.; Selaru, F.M.; Mori, Y.; Vazquez, C.; David, S.; Paun, B.; Cheng, Y.; Jin, Z.; Yang, J.; Agarwal, R.; et al. Dynamic changes in the expression of MicroRNA-31 during inflammatory bowel disease-associated neoplastic transformation. Inflamm. Bowel Dis. 2011, 17, 221–231. [Google Scholar] [CrossRef]
- Bian, Z.; Li, L.; Cui, J.; Zhang, H.; Liu, Y.; Zhang, C.-Y.; Zen, K. Role of miR-150-targeting c-Myb in colonic epithelial disruption during dextran sulphate sodium-induced murine experimental colitis and human ulcerative colitis. J. Pathol. 2011, 225, 544–553. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, C.; Liu, Y.; Tang, L.; Zheng, M.; Xu, C.; Song, J.; Meng, X. MiR-122 targets NOD2 to decrease intestinal epithelial cell injury in Crohn’s disease. Biochem. Biophys. Res. Commun. 2013, 438, 133–139. [Google Scholar] [CrossRef]
- Huang, Z.; Shi, T.; Zhou, Q.; Shi, S.; Zhao, R.; Shi, H.; Dong, L.; Zhang, C.; Zeng, K.; Chen, J.; et al. MIR-141 regulates colonic leukocytic trafficking by targeting CXCL12β during murine colitis and human crohn’s disease. Gut 2014, 63, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.T.; Dalmasso, G.; Yan, Y.; Laroui, H.; Dahan, S.; Mayer, L.; Sitaraman, S.V.; Merlin, D. MicroRNA-7 modulates CD98 expression during intestinal epithelial cell differentiation. J. Biol. Chem. 2010, 285, 1479–1489. [Google Scholar] [CrossRef] [PubMed]
- Nata, T.; Fujiya, M.; Ueno, N.; Moriichi, K.; Konishi, H.; Tanabe, H.; Ohtake, T.; Ikuta, K.; Kohgo, Y. MicroRNA-146b improves intestinal injury in mouse colitis by activating nuclear factor-κB and improving epithelial barrier function. J. Gene Med. 2013, 15, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Liang, Y.; Yang, J.; Xia, Y.; Chen, H.; Han, H.; Yang, Y.; Wu, W.; Gao, R.; Qin, H. MicroRNA-21 Knockout Improve the Survival Rate in DSS Induced Fatal Colitis through Protecting against Inflammation and Tissue Injury. PLoS ONE 2013, 8, e66814. [Google Scholar] [CrossRef]
- Pekow, J.; Meckel, K.; Dougherty, U.; Haider, H.I.; Deng, Z.; Hart, J.; Rubin, D.T.; Bissonnette, M. Increased mucosal expression of miR-215 precedes the development of neoplasia in patients with long-standing ulcerative colitis. Oncotarget 2018, 9, 20709–20720. [Google Scholar] [CrossRef]
- Feng, Y.; Dong, Y.W.; Song, Y.N.; Xiao, J.H.; Guo, X.Y.; Jiang, W.L.; Lu, L.G. MicroRNA-449a is a potential predictor of colitis-associated colorectal cancer progression. Oncol. Rep. 2018, 40, 1684–1694. [Google Scholar] [CrossRef]
- Lou, C.; Li, Y. Functional role of microRNA-135a in colitis. J. Inflamm. 2018, 15, 7. [Google Scholar] [CrossRef]
- Bai, J.; Li, Y.; Shao, T.; Zhao, Z.; Wang, Y.; Wu, A.; Chen, H.; Li, S.; Jiang, C.; Xu, J.; et al. Integrating analysis reveals microRNA-mediated pathway crosstalk among Crohn’s disease, ulcerative colitis and colorectal cancer. Mol. Biosyst. 2014, 10, 2317–2328. [Google Scholar] [CrossRef]
- Li, W.; Han, W.; Zhao, X.; Wang, H. [Changes of expression of miR-155 in colitis-associated colonic carcinogenesis]. Zhonghua Zhong Liu Za Zhi 2014, 36, 257–262. [Google Scholar]
- Eichele, D.D.; Kharbanda, K.K. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef]
- Keubler, L.M.; Buettner, M.; Häger, C.; Bleich, A. A multihit model: Colitis lessons from the interleukin-10-deficient mouse. Inflamm. Bowel Dis. 2015, 21, 1967–1975. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, E.; Kanneganti, M.; Mino-Kenudson, M. Animal models of colitis-associated carcinogenesis. J. Biomed. Biotechnol. 2011, 2011. [Google Scholar] [CrossRef]
- Mizoguchi, A.; Mizoguchi, E. Animal models of IBD: Linkage to human disease. Curr. Opin. Pharmacol. 2010, 10, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Guindi, M.; Riddell, R.H. Indeterminate colitis. J. Clin. Pathol. 2004, 57, 1233–1244. [Google Scholar] [CrossRef]
- Tontini, G.E.; Vecchi, M.; Pastorelli, L.; Neurath, M.F.; Neumann, H. Differential diagnosis in inflammatory bowel disease colitis: State of the art and future perspectives. World J. Gastroenterol. 2015, 21, 21–46. [Google Scholar] [CrossRef]
- Birimberg-Schwartz, L.; Zucker, D.M.; Akriv, A.; Cucchiara, S.; Cameron, F.L.; Wilson, D.C.; Łazowska, I.; Yianni, L.; Paul, S.P.; Romano, C.; et al. Development and validation of diagnostic criteria for IBD subtypes including IBdunclassified in children: A multicentre study from the pediatric IBD porto group of ESPGHAN. J. Crohn’s Colitis 2017, 1078–1084. [Google Scholar] [CrossRef]
- Schaefer, J.S.; Attumi, T.; Opekun, A.R.; Abraham, B.; Hou, J.; Shelby, H.; Graham, D.Y.; Streckfus, C.; Klein, J.R. MicroRNA signatures differentiate Crohn’s disease from ulcerative colitis. BMC Immunol. 2015, 16, 5. [Google Scholar] [CrossRef]
- Peck, B.C.E.; Weiser, M.; Lee, S.E.; Gipson, G.R.; Iyer, V.B.; Sartor, R.B.; Herfarth, H.H.; Long, M.D.; Hansen, J.J.; Isaacs, K.L.; et al. MicroRNAs classify different disease behavior phenotypes of Crohn’s disease and may have prognostic utility. Inflamm. Bowel Dis. 2015, 21, 2178–2187. [Google Scholar] [CrossRef]
- Kantono, M.; Guo, B. Inflammasomes and Cancer: The Dynamic Role of the Inflammasome in Tumor Development. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Khalafalla, F.G.; Khan, M.W. Inflammation and Epithelial-Mesenchymal Transition in Pancreatic Ductal Adenocarcinoma: Fighting Against Multiple Opponents. Cancer Growth Metastasis 2017, 10. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Shalapour, S.; Karin, M. Cruel to Be Kind: Epithelial, Microbial, and Immune Cell Interactions in Gastrointestinal Cancers. Annu. Rev. Immunol. 2020, 38, 649–671. [Google Scholar] [CrossRef] [PubMed]
- Todoric, J.; Karin, M. The Fire within: Cell-Autonomous Mechanisms in Inflammation-Driven Cancer. Cancer Cell 2019, 35, 714–720. [Google Scholar] [CrossRef] [PubMed]
- Malham, M.; Jakobsen, C.; Paerregaard, A.; Virta, L.J.; Kolho, K.L.; Wewer, V. The incidence of cancer and mortality in paediatric onset inflammatory bowel disease in Denmark and Finland during a 23-year period: A population-based study. Aliment. Pharmacol. Ther. 2019, 50, 33–39. [Google Scholar] [CrossRef]
- Chen, R.; Lai, L.A.; Brentnall, T.A.; Pan, S. Biomarkers for colitis-associated colorectal cancer. World J. Gastroenterol. 2016, 22, 7882–7891. [Google Scholar] [CrossRef]
- Duricova, D.; Pedersen, N.; Elkjaer, M.; Gamborg, M.; Munkholm, P.; Jess, T. Overall and cause-specific mortality in Crohn’s disease: A meta-analysis of population-based studies. Inflamm. Bowel Dis. 2010, 16, 347–353. [Google Scholar] [CrossRef]
- Jess, T.; Gamborg, M.; Matzen, P.; Munkholm, P.; Sorensen, T.I. Increased Risk of Intestinal Cancer in Crohn’s Disease: A Meta-Analysis of Population-Based Cohort Studies. Am. J. Gastroenterol. 2005, 100, 2724–2729. [Google Scholar] [CrossRef]
- Foersch, S.; Neurath, M.F. Colitis-associated neoplasia: Molecular basis and clinical translation. Cell. Mol. Life Sci. 2014, 71, 3523–3535. [Google Scholar] [CrossRef]
- Romano, M.; De Francesco, F.; Zarantonello, L.; Ruffolo, C.; Ferraro, G.A.; Zanus, G.; Giordano, A.; Bassi, N.; Cillo, U. From inflammation to cancer in inflammatory bowel disease: Molecular perspectives. Anticancer Res. 2016, 36, 1447–1460. [Google Scholar]
- Galandiuk, S.; Rodriguezjusto, M.; Jeffery, R.; Nicholson, A.M.; Cheng, Y.; Oukrif, D.; Elia, G.; Leedham, S.J.; McDonald, S.A.C.; Wright, N.A.; et al. Field cancerization in the intestinal epithelium of patients with Crohn’s ileocolitis. Gastroenterology 2012, 142, 855. [Google Scholar] [CrossRef]
- Risques, R.A.; Lai, L.A.; Himmetoglu, C.; Ebaee, A.; Li, L.; Feng, Z.; Bronner, M.P.; Al-Lahham, B.; Kowdley, K.V.; Lindor, K.D.; et al. Ulcerative colitis-associated colorectal cancer arises in a field of short telomeres, senescence, and inflammation. Cancer Res. 2011, 71, 1669–1679. [Google Scholar] [CrossRef]
- Karvellas, C.J.; Fedorak, R.N.; Hanson, J.; Wong, C.K.W. Increased risk of colorectal cancer in ulcerative colitis patients diagnosed after 40 years of age. Can. J. Gastroenterol. 2007, 21, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Den Low, E.N.; Mokhtar, N.M.; Wong, Z.; Raja Ali, R.A. Colonic mucosal transcriptomic changes in patients with long-duration ulcerative colitis revealed colitis-associated cancer pathways. J. Crohn’s Colitis 2019, 13, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, T.M.; Brückner, M.; Eveslage, M.; Tepasse, P.; Pott, F.; Thoennissen, N.H.; Hengst, K.; Ross, M.; Bettenworth, D. The Risk of Colorectal Cancer in Patients with Ulcerative Colitis. Dig. Dis. Sci. 2015, 60, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Josse, C.; Bours, V. MicroRNAs and inflammation in colorectal cancer. Adv. Exp. Med. Biol. 2016, 937, 53–69. [Google Scholar] [CrossRef]
- Ranjha, R.; Aggarwal, S.; Bopanna, S.; Ahuja, V.; Paul, J. Site-specific MicroRNA expression may lead to different subtypes in ulcerative colitis. PLoS ONE 2015, 10, e0142869. [Google Scholar] [CrossRef]
- Itzkowitz, S.H.; Yio, X. Inflammation and cancer—IV. Colorectal cancer in inflammatory bowel disease: The role of inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G7–G17. [Google Scholar] [CrossRef]
- He, C.; Yu, T.; Shi, Y.; Ma, C.; Yang, W.; Fang, L.; Sun, M.; Wu, W.; Xiao, F.; Guo, F.; et al. MicroRNA 301A Promotes Intestinal Inflammation and Colitis-Associated Cancer Development by Inhibiting BTG1. Gastroenterology 2017, 152, 1434–1448.e15. [Google Scholar] [CrossRef]
- Signs, S.A.; Fisher, R.C.; Tran, U.; Chakrabarti, S.; Sarvestani, S.K.; Xiang, S.; Liska, D.; Roche, V.; Lai, W.; Gittleman, H.R.; et al. Stromal miR-20a controls paracrine CXCL8 secretion in colitis and colon cancer. Oncotarget 2018, 9, 13048–13059. [Google Scholar] [CrossRef]
- Krichevsky, A.M.; Gabriely, G. miR-21: A small multi-faceted RNA. J. Cell. Mol. Med. 2008, 13, 39–53. [Google Scholar] [CrossRef]
- Bautista-Sánchez, D.; Arriaga-Canon, C.; Pedroza-Torres, A.; De La Rosa-Velázquez, I.A.; González-Barrios, R.; Contreras-Espinosa, L.; Montiel-Manríquez, R.; Castro-Hernández, C.; Fragoso-Ontiveros, V.; Álvarez-Gómez, R.M.; et al. The Promising Role of miR-21 as a Cancer Biomarker and Its Importance in RNA-Based Therapeutics. Mol. Ther. Nucleic Acids 2020, 20, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, B.S.; Jørgensen, S.; Fog, J.U.; Søkilde, R.; Christensen, I.J.; Hansen, U.; Brünner, N.; Baker, A.; Møller, S.; Nielsen, H.J. High levels of microRNA-21 in the stroma of colorectal cancers predict short disease-free survival in stage II colon cancer patients. Clin. Exp. Metastasis 2011, 28, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, K.; Fassan, M.; Mescoli, C.; Pizzi, M.; Balistreri, M.; Albertoni, L.; Pucciarelli, S.; Scarpa, M.; Sturniolo, G.C.; Angriman, I.; et al. PDCD4/miR-21 dysregulation in inflammatory bowel disease-associated carcinogenesis. Virchows Arch. 2013, 462, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Frankel, L.B.; Christoffersen, N.R.; Jacobsen, A.; Lindow, M.; Krogh, A.; Lund, A.H. Programmed cell death 4 (PDCD4) is an important functional target of the microRNA miR-21 in breast cancer cells. J. Biol. Chem. 2008, 283, 1026–1033. [Google Scholar] [CrossRef] [PubMed]
- Chu, E.C.; Tarnawski, A.S. PTEN regulatory functions in tumor suppression and cell biology. Med. Sci. Monit. 2004, 10, RA235-41. [Google Scholar]
- Cho, W.C.S. OncomiRs: The discovery and progress of microRNAs in cancers. Mol. Cancer 2007, 6, 60. [Google Scholar] [CrossRef]
- Ando, Y.; Yang, G.-X.; Kenny, T.P.; Kawata, K.; Zhang, W.; Huang, W.; Leung, P.S.C.; Lian, Z.-X.; Okazaki, K.; Ansari, A.A.; et al. Overexpression of microRNA-21 is associated with elevated pro- inflammatory cytokines in dominant-negative TGF-β receptor type II mouse. J. Autoimmun. 2013, 41, 111–119. [Google Scholar] [CrossRef]
- Hou, J.; Hu, X.; Chen, B.; Chen, X.; Zhao, L.; Chen, Z.; Liu, F.; Liu, Z. miR-155 targets Est-1 and induces ulcerative colitis via the IL-23/17/6-mediated Th17 pathway. Pathol. Res. Pract. 2017, 213, 1289–1295. [Google Scholar] [CrossRef]
- Paraskevi, A.; Theodoropoulos, G.; Papaconstantinou, I.; Mantzaris, G.; Nikiteas, N.; Gazouli, M. Circulating MicroRNA in inflammatory bowel disease. J. Crohn’s Colitis 2012, 6, 900–904. [Google Scholar] [CrossRef]
- Kara, M.; Yumrutas, O.; Ozcan, O.; Celik, O.I.; Bozgeyik, E.; Bozgeyik, I.; Tasdemir, S. Differential expressions of cancer-associated genes and their regulatory miRNAs in colorectal carcinoma. Gene 2015, 567, 81–86. [Google Scholar] [CrossRef]
- Shibuya, H.; Iinuma, H.; Shimada, R.; Horiuchi, A.; Watanabe, T. Clinicopathological and prognostic value of microRNA-21 and microRNA-155 in colorectal cancer. Oncology 2011, 79, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.-c.; Fan, Y.-s.; Chen, H.-b.; Zhao, D.-w. Investigation of microRNA-155 as a serum diagnostic and prognostic biomarker for colorectal cancer. Tumor Biol. 2014, 36, 1619–1625. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, Z.; Xiang, J.; Gu, X. MicroRNA-155 acts as a tumor suppressor in colorectal cancer by targeting CTHRC1 in vitro. Oncol. Lett. 2018, 15, 5561–5568. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, F.; Li, H.; Fan, H.; Wu, H.; Dong, Y.; Chu, S.; Tan, C.; Wang, Q.; He, H.; et al. MiR-155 contributes to intestinal barrier dysfunction in DSS-induced mice colitis via targeting HIF-1α/TFF-3 axis. Aging (Albany N.Y.) 2020, 12, 14966–14977. [Google Scholar] [CrossRef]
- Karimi, N.; Feizi, M.A.H.; Safaralizadeh, R.; Hashemzadeh, S.; Baradaran, B.; Shokouhi, B.; Teimourian, S. Serum overexpression of miR-301a and miR-23a in patients with colorectal cancer. J. Chin. Med. Assoc. 2019, 82, 215–220. [Google Scholar] [CrossRef]
- Loftus, E.V.; Kane, S.V.; Bjorkman, D. Systematic review: Short-term adverse effects of 5-aminosalicylic acid agents in the treatment of ulcerative colitis. Aliment. Pharmacol. Ther. 2004, 19, 179–189. [Google Scholar] [CrossRef]
- Scarpa, M.; Castagliuolo, I.; Castoro, C.; Pozza, A.; Scarpa, M.; Kotsafti, A.; Angriman, I. Inflammatory colonic carcinogenesis: A review on pathogenesis and immunosurveillance mechanisms in ulcerative colitis. World J. Gastroenterol. 2014, 20, 6774–6785. [Google Scholar] [CrossRef]
- Williams, C.; Panaccione, R.; Ghosh, S.; Rioux, K. Optimizing clinical use of mesalazine (5-aminosalicylic acid) in inflammatory bowel disease. Ther. Adv. Gastroenterol. 2011, 4, 237–248. [Google Scholar] [CrossRef]
- Olesen, C.M.; Coskun, M.; Peyrin-Biroulet, L.; Nielsen, O.H. Mechanisms behind efficacy of tumor necrosis factor inhibitors in inflammatory bowel diseases. Pharmacol. Ther. 2016, 159, 110–119. [Google Scholar] [CrossRef]
- Espinosa Morales, R.; Díaz Borjón, A.; Barile Fabris, L.A.; Esquivel Valerio, J.A.; Medrano Ramírez, G.; Arce Salinas, C.A.; Barreira Mercado, E.R.; Cardiel Ríos, M.H.; Díaz Jouanen, E.; Flores Murrieta, F.J.; et al. Long-term infliximab maintenance therapy for ulcerative colitis: The ACT-1 and -2 extension studies. Inflamm. Bowel Dis. 2012. [Google Scholar] [CrossRef]
- Nielsen, O.H.; Ainsworth, M.A. Tumor necrosis factor inhibitors for inflammatory bowel disease. N. Engl. J. Med. 2013, 369, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Ben-Horin, S.; Chowers, Y. Review article: Loss of response to anti-TNF treatments in Crohn’s disease. Aliment. Pharmacol. Ther. 2011, 33, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Morilla, I.; Uzzan, M.; Laharie, D.; Cazals-Hatem, D.; Denost, Q.; Daniel, F.; Belleannee, G.; Bouhnik, Y.; Wainrib, G.; Panis, Y.; et al. Colonic MicroRNA Profiles, Identified by a Deep Learning Algorithm, That Predict Responses to Therapy of Patients With Acute Severe Ulcerative Colitis. Clin. Gastroenterol. Hepatol. 2019, 17, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Sandborn, W.J.; Gasink, C.; Gao, L.L.; Blank, M.A.; Johanns, J.; Guzzo, C.; Sands, B.E.; Hanauer, S.B.; Targan, S.; Rutgeerts, P.; et al. Ustekinumab induction and maintenance therapy in refractory Crohn’s disease. N. Engl. J. Med. 2012, 367, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.J.; Noh, J.H.; Kim, J.K.; Eun, J.W.; Jung, K.H.; Kim, M.G.; Chang, Y.G.; Shen, Q.; Kim, S.-J.; Park, W.S.; et al. MicroRNA-29c functions as a tumor suppressor by direct targeting oncogenic SIRT1 in hepatocellular carcinoma. Oncogene 2014, 33, 2557–2567. [Google Scholar] [CrossRef] [PubMed]
- Fedyk, E.R.; Wyant, T.; Yang, L.L.; Csizmadia, V.; Burke, K.; Yang, H.; Kadambi, V.J. Exclusive antagonism of the α4β7 integrin by vedolizumab confirms the gut-selectivity of this pathway in primates. Inflamm. Bowel Dis. 2012, 18, 2107–2119. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.A.; Yamakuchi, M.; Ferlito, M.; Mendell, J.T.; Lowenstein, C.J. MicroRNA-126 regulates endothelial expression of vascular cell adhesion molecule 1. Proc. Natl. Acad. Sci. USA 2008, 105, 1516–1521. [Google Scholar] [CrossRef]
- Sandborn, W.J.; Ghosh, S.; Panes, J.; Vranic, I.; Su, C.; Rousell, S.; Niezychowski, W. Tofacitinib, an Oral Janus Kinase Inhibitor, in Active Ulcerative Colitis. N. Engl. J. Med. 2012, 367, 616–624. [Google Scholar] [CrossRef]
- Magro, F.; Estevinho, M.M. Is tofacitinib a game-changing drug for ulcerative colitis? United Eur. Gastroenterol. J. 2020, 8, 755–763. [Google Scholar] [CrossRef]
- Kim, H.; Banerjee, N.; Sirven, M.A.; Minamoto, Y.; Markel, M.E.; Suchodolski, J.S.; Talcott, S.T.; Mertens-Talcott, S.U. Pomegranate polyphenolics reduce inflammation and ulceration in intestinal colitis—Involvement of the miR-145/p70S6K1/HIF1α axis in vivo and in vitro. J. Nutr. Biochem. 2017, 43, 107–115. [Google Scholar] [CrossRef]
- Maschmeyer, P.; Petkau, G.; Siracusa, F.; Zimmermann, J.; Zügel, F.; Kühl, A.A.; Lehmann, K.; Schimmelpfennig, S.; Weber, M.; Haftmann, C.; et al. Selective targeting of pro-inflammatory Th1 cells by microRNA-148a-specific antagomirs in vivo. J. Autoimmun. 2018, 89, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Whiteoak, S.R.; Claridge, A.; Balendran, C.A.; Harris, R.J.; Gwiggner, M.; Bondanese, V.P.; Erlandsson, F.; Hansen, M.B.; Cummings, J.R.F.; Sanchez-Elsner, T. MicroRNA-31 Targets Thymic Stromal Lymphopoietin in Mucosal Infiltrated CD4+ T Cells: A Role in Achieving Mucosal Healing in Ulcerative Colitis? Inflamm. Bowel Dis. 2018, 24, 2377–2385. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.Q.; Wang, J.; Chen, G.X.; Zhang, G.Q.; Zhang, D.Y.; Cai, Y.Y. Overexpression of microRNA-495 improves the intestinal mucosal barrier function by targeting STAT3 via inhibition of the JAK/STAT3 signaling pathway in a mouse model of ulcerative colitis. Pathol. Res. Pract. 2018, 214, 151–162. [Google Scholar] [CrossRef]
- Jørgensen, S.; Baker, A.; Møller, S.; Nielsen, B.S. Robust one-day in situ hybridization protocol for detection of microRNAs in paraffin samples using LNA probes. Methods 2010, 52, 375–381. [Google Scholar] [CrossRef]
- Nielsen, B.S.; Holmstrøm, K. Target Identification and Validation in Drug Discovery; Moll, J., Colombo, R., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2013; Volume 986, ISBN 978-1-62703-310-7. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).