Application of Mesenchymal Stem Cells in Inflammatory and Fibrotic Diseases

 ,
,
Abstract
1. Introduction
2. Human Mesenchymal Stem Cells (hMSCs)
2.1. Criteria for the Characterization of hMSCs for Clinical Applications
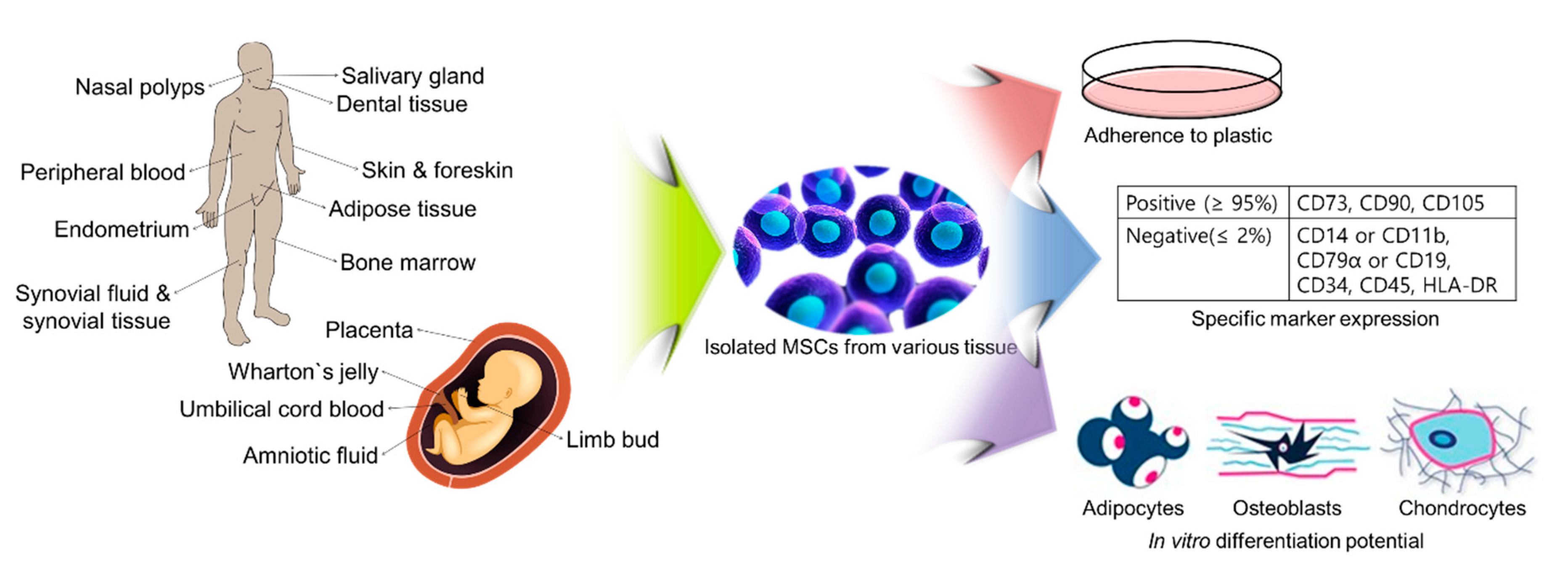
2.2. Isolation of hMSCs from Various Tissues
3. Mesenchymal Stem Cells and Inflammatory Diseases
3.1. Graft-Versus-Host Disease (GVHD)
3.2. Multiple Sclerosis (MS)
3.3. Type 1 Diabetes (T1D)
3.4. Joint Diseases: Osteoarthritis (OA) and Rheumatoid Arthritis (RA)
3.5. Inflammatory Bowel Diseases (IBD)
3.6. Systemic Lupus Erythematosus (SLE)
3.7. Chronic Rhinosinusitis with Nasal Polyp (CRSwNP)
4. Mesenchymal Stem Cells and Fibrotic Diseases
4.1. Lung Fibrosis
4.2. Liver Fibrosis
4.3. Pancreatic Fibrosis
4.4. Heart Fibrosis
5. Conclusion
Author Contributions
Funding
Conflicts of Interest
References
- Wei, X.; Yang, X.; Han, Z.P.; Qu, F.F.; Shao, L.; Shi, Y.F. Mesenchymal stem cells: A new trend for cell therapy. Acta Pharm. Sin. 2013, 34, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Bongso, A.; Richards, M. History and perspective of stem cell research. Best Pr. Res. Clin. Obs. Gynaecol. 2004, 18, 827–842. [Google Scholar] [CrossRef] [PubMed]
- Ilic, D.; Polak, J.M. Stem cells in regenerative medicine: Introduction. Br. Med. Bull. 2011, 98, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.J.; Kaufman, M.H. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981, 292, 154–156. [Google Scholar] [CrossRef]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Cunningham, J.J.; Ulbright, T.M.; Pera, M.F.; Looijenga, L.H. Lessons from human teratomas to guide development of safe stem cell therapies. Nat. Biotechnol. 2012, 30, 849–857. [Google Scholar] [CrossRef]
- Zarzeczny, A.; Caulfield, T. Emerging ethical, legal and social issues associated with stem cell research & and the current role of the moral status of the embryo. Stem Cell Rev. Rep. 2009, 5, 96–101. [Google Scholar]
- Ryu, J.S.; Seo, S.Y.; Jeong, E.J.; Kim, J.Y.; Koh, Y.G.; Kim, Y.I.; Choo, Y.K. Ganglioside GM3 up-regulate chondrogenic differentiation by transform growth factor receptors. Int. J. Mol. Sci. 2020, 21, 1967. [Google Scholar] [CrossRef]
- Kim, Y.I.; Ryu, J.S.; Yeo, J.E.; Choi, Y.J.; Kim, Y.S.; Ko, K.; Koh, Y.G. Overexpression of TGF-beta1 enhances chondrogenic differentiation and proliferation of human synovium-derived stem cells. Biochem. Biophys. Res. Commun. 2014, 450, 1593–1599. [Google Scholar] [CrossRef]
- Yang, H.J.; Jung, K.Y.; Kwak, D.H.; Lee, S.H.; Ryu, J.S.; Kim, J.S.; Chang, K.T.; Lee, J.W.; Choo, Y.K. Inhibition of ganglioside GD1a synthesis suppresses the differentiation of human mesenchymal stem cells into osteoblasts. Dev. Growth Differ. 2011, 53, 323–332. [Google Scholar] [CrossRef]
- Lee, S.H.; Ryu, J.S.; Lee, J.W.; Kwak, D.H.; Ko, K.; Choo, Y.K. Comparison of ganglioside expression between human adipose-and dental pulp-derived stem cell differentiation into osteoblasts. Arch. Pharm. Res. 2010, 33, 585–591. [Google Scholar] [CrossRef]
- Kim, S.M.; Jung, J.U.; Ryu, J.S.; Jin, J.W.; Yang, H.J.; Ko, K.; You, H.K.; Jung, K.Y.; Choo, Y.K. Effects of gangliosides on the differentiation of human mesenchymal stem cells into osteoblasts by modulating epidermal growth factor receptors. Biochem. Biophys. Res. Commun. 2008, 371, 866–871. [Google Scholar] [CrossRef]
- Qu, P.; Wang, L.; Min, Y.; McKennett, L.; Keller, J.R.; Lin, P.C. Vav1 regulates mesenchymal stem cell differentiation decision between adipocyte and chondrocyte via Sirt1. Stem Cells 2016, 34, 1934–1946. [Google Scholar] [CrossRef] [PubMed]
- Kanda, Y.; Hinata, T.; Kang, S.W.; Watanabe, Y. Reactive oxygen species mediate adipocyte differentiation in mesenchymal stem cells. Life Sci. 2011, 89, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef]
- Pezato, R.; de Almeida, D.C.; Bezerra, T.F.; Silva, F.D.S.; Perez-Novo, C.; Gregorio, L.C.; Voegels, R.L.; Camara, N.O.; Bachert, C. Immunoregulatory effects of bone marrow-derived mesenchymal stem cells in the nasal polyp microenvironment. Mediat. Inflamm. 2014, 2014, 583409. [Google Scholar] [CrossRef]
- Mamidi, M.K.; Nathan, K.G.; Singh, G.; Thrichelvam, S.T.; Mohd Yusof, N.A.; Fakharuzi, N.A.; Zakaria, Z.; Bhonde, R.; Das, A.K.; Majumdar, A.S. Comparative cellular and molecular analyses of pooled bone marrow multipotent mesenchymal stromal cells during continuous passaging and after successive cryopreservation. J. Cell. Biochem. 2012, 113, 3153–3164. [Google Scholar] [CrossRef]
- Otsuru, S.; Hofmann, T.J.; Olson, T.S.; Dominici, M.; Horwitz, E.M. Improved isolation and expansion of bone marrow mesenchymal stromal cells using a novel marrow filter device. Cytotherapy 2013, 15, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Graves, S.E.; Ohta, S.; Simmons, P.J. The STRO-1+ fraction of adult human bone marrow contains the osteogenic precursors. Blood 1994, 84, 4164–4173. [Google Scholar] [CrossRef]
- Stewart, K.; Walsh, S.; Screen, J.; Jefferiss, C.M.; Chainey, J.; Jordan, G.R.; Beresford, J.N. Further characterization of cells expressing STRO-1 in cultures of adult human bone marrow stromal cells. J. Bone. Min. Res. 1999, 14, 1345–1356. [Google Scholar] [CrossRef]
- Wagner, W.; Wein, F.; Seckinger, A.; Frankhauser, M.; Wirkner, U.; Krause, U.; Blake, J.; Schwager, C.; Eckstein, V.; Ansorge, W.; et al. Comparative characteristics of mesenchymal stem cells from human bone marrow, adipose tissue, and umbilical cord blood. Exp. Hematol. 2005, 33, 1402–1416. [Google Scholar] [CrossRef]
- Ranera, B.; Remacha, A.R.; Alvarez-Arguedas, S.; Castiella, T.; Vazquez, F.J.; Romero, A.; Zaragoza, P.; Martin-Burriel, I.; Rodellar, C. Expansion under hypoxic conditions enhances the chondrogenic potential of equine bone marrow-derived mesenchymal stem cells. Vet. J. 2013, 195, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hirai, M.; Cantero, S.; Ciubotariu, R.; Dobrila, L.; Hirsh, A.; Igura, K.; Satoh, H.; Yokomi, I.; Nishimura, T.; et al. Isolation and characterization of mesenchymal stem cells from human umbilical cord blood: Reevaluation of critical factors for successful isolation and high ability to proliferate and differentiate to chondrocytes as compared to mesenchymal stem cells from bone marrow and adipose tissue. J. Cell. Biochem. 2011, 112, 1206–1218. [Google Scholar]
- Muruganandan, S.; Roman, A.A.; Sinal, C.J. Adipocyte differentiation of bone marrow-derived mesenchymal stem cells: Cross talk with the osteoblastogenic program. Cell. Mol. Life Sci. 2009, 66, 236–253. [Google Scholar] [CrossRef]
- Stock, P.; Bruckner, S.; Winkler, S.; Dollinger, M.M.; Christ, B. Human bone marrow mesenchymal stem cell-derived hepatocytes improve the mouse liver after acute acetaminophen intoxication by preventing progress of injury. Int. J. Mol. Sci. 2014, 15, 7004–7028. [Google Scholar] [CrossRef]
- Xu, W.; Zhang, X.; Qian, H.; Zhu, W.; Sun, X.; Hu, J.; Zhou, H.; Chen, Y. Mesenchymal stem cells from adult human bone marrow differentiate into a cardiomyocyte phenotype in vitro. Exp. Biol. Med. (Maywood) 2004, 229, 623–631. [Google Scholar] [CrossRef]
- Tang, D.Q.; Wang, Q.; Burkhardt, B.R.; Litherland, S.A.; Atkinson, M.A.; Yang, L.J. In vitro generation of functional insulin-producing cells from human bone marrow-derived stem cells, but long-term culture running risk of malignant transformation. Am. J. Stem Cells 2012, 1, 114–127. [Google Scholar]
- Gabr, M.M.; Zakaria, M.M.; Refaie, A.F.; Ismail, A.M.; Abou-El-Mahasen, M.A.; Ashamallah, S.A.; Khater, S.M.; El-Halawani, S.M.; Ibrahim, R.Y.; Uin, G.S.; et al. Insulin-producing cells from adult human bone marrow mesenchymal stem cells control streptozotocin-induced diabetes in nude mice. Cell Transpl. 2013, 22, 133–145. [Google Scholar] [CrossRef]
- Phadnis, S.M.; Joglekar, M.V.; Dalvi, M.P.; Muthyala, S.; Nair, P.D.; Ghaskadbi, S.M.; Bhonde, R.R.; Hardikar, A.A. Human bone marrow-derived mesenchymal cells differentiate and mature into endocrine pancreatic lineage in vivo. Cytotherapy 2011, 13, 279–293. [Google Scholar] [CrossRef]
- Barzilay, R.; Ben-Zur, T.; Bulvik, S.; Melamed, E.; Offen, D. Lentiviral delivery of LMX1a enhances dopaminergic phenotype in differentiated human bone marrow mesenchymal stem cells. Stem Cells Dev. 2009, 18, 591–601. [Google Scholar] [CrossRef]
- Wilkins, A.; Kemp, K.; Ginty, M.; Hares, K.; Mallam, E.; Scolding, N. Human bone marrow-derived mesenchymal stem cells secrete brain-derived neurotrophic factor which promotes neuronal survival in vitro. Stem Cell Res. 2009, 3, 63–70. [Google Scholar] [CrossRef]
- Bieback, K.; Kern, S.; Kluter, H.; Eichler, H. Critical parameters for the isolation of mesenchymal stem cells from umbilical cord blood. Stem Cells 2004, 22, 625–634. [Google Scholar] [CrossRef]
- Miao, Z.; Jin, J.; Chen, L.; Zhu, J.; Huang, W.; Zhao, J.; Qian, H.; Zhang, X. Isolation of mesenchymal stem cells from human placenta: Comparison with human bone marrow mesenchymal stem cells. Cell Biol. Int. 2006, 30, 681–687. [Google Scholar] [CrossRef]
- La Rocca, G.; Anzalone, R.; Corrao, S.; Magno, F.; Loria, T.; Lo Iacono, M.; Di Stefano, A.; Giannuzzi, P.; Marasa, L.; Cappello, F.; et al. Isolation and characterization of Oct-4+/HLA-G+ mesenchymal stem cells from human umbilical cord matrix: Differentiation potential and detection of new markers. Histochem. Cell Biol. 2009, 131, 267–282. [Google Scholar] [CrossRef]
- Kita, K.; Gauglitz, G.G.; Phan, T.T.; Herndon, D.N.; Jeschke, M.G. Isolation and characterization of mesenchymal stem cells from the sub-amniotic human umbilical cord lining membrane. Stem Cells Dev. 2010, 19, 491–502. [Google Scholar] [CrossRef]
- Moretti, P.; Hatlapatka, T.; Marten, D.; Lavrentieva, A.; Majore, I.; Hass, R.; Kasper, C. Mesenchymal stromal cells derived from human umbilical cord tissues: Primitive cells with potential for clinical and tissue engineering applications. Adv. Biochem. Eng. Biotechnol. 2010, 123, 29–54. [Google Scholar]
- Majore, I.; Moretti, P.; Stahl, F.; Hass, R.; Kasper, C. Growth and differentiation properties of mesenchymal stromal cell populations derived from whole human umbilical cord. Stem Cell Rev. Rep. 2011, 7, 17–31. [Google Scholar] [CrossRef]
- Hang, H.; Yu, Y.; Wu, N.; Huang, Q.; Xia, Q.; Bian, J. Induction of highly functional hepatocytes from human umbilical cord mesenchymal stem cells by HNF4alpha transduction. PLoS ONE 2014, 9, e104133. [Google Scholar] [CrossRef]
- An, S.Y.; Han, J.; Lim, H.J.; Park, S.Y.; Kim, J.H.; Do, B.R.; Kim, J.H. Valproic acid promotes differentiation of hepatocyte-like cells from whole human umbilical cord-derived mesenchymal stem cells. Tissue Cell 2014, 46, 127–135. [Google Scholar] [CrossRef]
- Prabakar, K.R.; Dominguez-Bendala, J.; Molano, R.D.; Pileggi, A.; Villate, S.; Ricordi, C.; Inverardi, L. Generation of glucose-responsive, insulin-producing cells from human umbilical cord blood-derived mesenchymal stem cells. Cell Transpl. 2012, 21, 1321–1339. [Google Scholar] [CrossRef]
- Zhao, F.; Qu, Y.; Liu, H.; Du, B.; Mu, D. Umbilical cord blood mesenchymal stem cells co-modified by TERT and BDNF: A novel neuroprotective therapy for neonatal hypoxic-ischemic brain damage. Int. J. Dev. Neurosci. 2014, 38, 147–154. [Google Scholar] [CrossRef]
- Pendleton, C.; Li, Q.; Chesler, D.A.; Yuan, K.; Guerrero-Cazares, H.; Quinones-Hinojosa, A. Mesenchymal stem cells derived from adipose tissue vs. bone marrow: In vitro comparison of their tropism towards gliomas. PLoS ONE 2013, 8, e58198. [Google Scholar] [CrossRef]
- Baglioni, S.; Francalanci, M.; Squecco, R.; Lombardi, A.; Cantini, G.; Angeli, R.; Gelmini, S.; Guasti, D.; Benvenuti, S.; Annunziato, F.; et al. Characterization of human adult stem-cell populations isolated from visceral and subcutaneous adipose tissue. FASEB J. 2009, 23, 3494–3505. [Google Scholar] [CrossRef]
- Gronthos, S.; Franklin, D.M.; Leddy, H.A.; Robey, P.G.; Storms, R.W.; Gimble, J.M. Surface protein characterization of human adipose tissue-derived stromal cells. J. Cell. Physiol. 2001, 189, 54–63. [Google Scholar] [CrossRef]
- Kim, Y.S.; Choi, Y.J.; Suh, D.S.; Heo, D.B.; Kim, Y.I.; Ryu, J.S.; Koh, Y.G. Mesenchymal stem cell implantation in osteoarthritic knees: Is fibrin glue effective as a scaffold? Am. J. Sports Med. 2015, 43, 176–185. [Google Scholar] [CrossRef]
- Cho, K.S.; Kim, Y.W.; Kang, M.J.; Park, H.Y.; Hong, S.L.; Roh, H.J. Immunomodulatory effect of mesenchymal stem cells on T lymphocyte and cytokine expression in nasal polyps. Otolaryngol. Head Neck Surg. 2014, 150, 1062–1070. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, F.; Zhao, H.; Zhang, X.; Chen, H.; Zhang, K. Human adipose-derived mesenchymal stem cells are resistant to HBV infection during differentiation into hepatocytes in vitro. Int. J. Mol. Sci. 2014, 15, 6096–6110. [Google Scholar] [CrossRef]
- Choi, Y.S.; Dusting, G.J.; Stubbs, S.; Arunothayaraj, S.; Han, X.L.; Collas, P.; Morrison, W.A.; Dilley, R.J. Differentiation of human adipose-derived stem cells into beating cardiomyocytes. J. Cell. Mol. Med. 2010, 14, 878–889. [Google Scholar] [CrossRef]
- Timper, K.; Seboek, D.; Eberhardt, M.; Linscheid, P.; Christ-Crain, M.; Keller, U.; Muller, B.; Zulewski, H. Human adipose tissue-derived mesenchymal stem cells differentiate into insulin, somatostatin, and glucagon expressing cells. Biochem. Biophys. Res. Commun. 2006, 341, 1135–1140. [Google Scholar] [CrossRef]
- Safford, K.M.; Hicok, K.C.; Safford, S.D.; Halvorsen, Y.D.; Wilkison, W.O.; Gimble, J.M.; Rice, H.E. Neurogenic differentiation of murine and human adipose-derived stromal cells. Biochem. Biophys. Res. Commun. 2002, 294, 371–379. [Google Scholar] [CrossRef]
- Kang, S.K.; Lee, D.H.; Bae, Y.C.; Kim, H.K.; Baik, S.Y.; Jung, J.S. Improvement of neurological deficits by intracerebral transplantation of human adipose tissue-derived stromal cells after cerebral ischemia in rats. Exp. Neurol. 2003, 183, 355–366. [Google Scholar] [CrossRef]
- Kuznetsov, S.A.; Krebsbach, P.H.; Satomura, K.; Kerr, J.; Riminucci, M.; Benayahu, D.; Robey, P.G. Single-colony derived strains of human marrow stromal fibroblasts form bone after transplantation in vivo. J. Bone Min. Res. 1997, 12, 1335–1347. [Google Scholar] [CrossRef]
- Hou, T.; Xu, J.; Wu, X.; Xie, Z.; Luo, F.; Zhang, Z.; Zeng, L. Umbilical cord Wharton’s Jelly: A new potential cell source of mesenchymal stromal cells for bone tissue engineering. Tissue Eng. Part A 2009, 15, 2325–2334. [Google Scholar] [CrossRef]
- Salehinejad, P.; Alitheen, N.B.; Ali, A.M.; Omar, A.R.; Mohit, M.; Janzamin, E.; Samani, F.S.; Torshizi, Z.; Nematollahi-Mahani, S.N. Comparison of different methods for the isolation of mesenchymal stem cells from human umbilical cord Wharton’s Jelly. In Vitro Cell. Dev. Biol. Anim. 2012, 48, 75–83. [Google Scholar] [CrossRef]
- Christodoulou, I.; Kolisis, F.N.; Papaevangeliou, D.; Zoumpourlis, V. Comparative evaluation of human mesenchymal stem cells of fetal (Wharton’s Jelly) and adult (Adipose Tissue) origin during prolonged in vitro expansion: Considerations for cytotherapy. Stem Cells Int. 2013, 2013, 246134. [Google Scholar] [CrossRef]
- Yoon, J.H.; Roh, E.Y.; Shin, S.; Jung, N.H.; Song, E.Y.; Chang, J.Y.; Kim, B.J.; Jeon, H.W. Comparison of explant-derived and enzymatic digestion-derived MSCs and the growth factors from Wharton’s Jelly. Biomed. Res. Int. 2013, 2013, 428726. [Google Scholar] [CrossRef]
- Wang, H.S.; Hung, S.C.; Peng, S.T.; Huang, C.C.; Wei, H.M.; Guo, Y.J.; Fu, Y.S.; Lai, M.C.; Chen, C.C. Mesenchymal stem cells in the Wharton’s Jelly of the human umbilical cord. Stem Cells 2004, 22, 1330–1337. [Google Scholar] [CrossRef]
- Anzalone, R.; Lo Iacono, M.; Corrao, S.; Magno, F.; Loria, T.; Cappello, F.; Zummo, G.; Farina, F.; La Rocca, G. New emerging potentials for human Wharton’s Jelly mesenchymal stem cells: Immunological features and hepatocyte-like differentiative capacity. Stem Cells Dev. 2010, 19, 423–438. [Google Scholar] [CrossRef]
- Datta, I.; Mishra, S.; Mohanty, L.; Pulikkot, S.; Joshi, P.G. Neuronal plasticity of human Wharton’s Jelly mesenchymal stromal cells to the dopaminergic cell type compared with human bone marrow mesenchymal stromal cells. Cytotherapy 2011, 13, 918–932. [Google Scholar] [CrossRef]
- Int Anker, P.S.; Scherjon, S.A.; Kleijburg-van der Keur, C.; Noort, W.A.; Claas, F.H.; Willemze, R.; Fibbe, W.E.; Kanhai, H.H. Amniotic fluid as a novel source of mesenchymal stem cells for therapeutic transplantation. Blood 2003, 102, 1548–1549. [Google Scholar] [CrossRef]
- Tsai, M.S.; Lee, J.L.; Chang, Y.J.; Hwang, S.M. Isolation of human multipotent mesenchymal stem cells from second-trimester amniotic fluid using a novel two-stage culture protocol. Hum. Reprod. 2004, 19, 1450–1456. [Google Scholar] [CrossRef]
- Cai, J.; Li, W.; Su, H.; Qin, D.; Yang, J.; Zhu, F.; Xu, J.; He, W.; Guo, X.; Labuda, K.; et al. Generation of human induced pluripotent stem cells from umbilical cord matrix and amniotic membrane mesenchymal cells. J. Biol. Chem. 2010, 285, 11227–11234. [Google Scholar] [CrossRef]
- Huang, G.T.; Gronthos, S.; Shi, S. Mesenchymal stem cells derived from dental tissues vs. those from other sources: Their biology and role in regenerative medicine. J. Dent. Res. 2009, 88, 792–806. [Google Scholar] [CrossRef]
- Hilkens, P.; Gervois, P.; Fanton, Y.; Vanormelingen, J.; Martens, W.; Struys, T.; Politis, C.; Lambrichts, I.; Bronckaers, A. Effect of isolation methodology on stem cell properties and multilineage differentiation potential of human dental pulp stem cells. Cell Tissue Res. 2013, 353, 65–78. [Google Scholar] [CrossRef]
- Seifrtova, M.; Havelek, R.; Cmielova, J.; Jiroutova, A.; Soukup, T.; Bruckova, L.; Mokry, J.; English, D.; Rezacova, M. The response of human ectomesenchymal dental pulp stem cells to cisplatin treatment. Int. Endod. J. 2012, 45, 401–412. [Google Scholar] [CrossRef]
- Kadar, K.; Kiraly, M.; Porcsalmy, B.; Molnar, B.; Racz, G.Z.; Blazsek, J.; Kallo, K.; Szabo, E.L.; Gera, I.; Gerber, G.; et al. Differentiation potential of stem cells from human dental origin—Promise for tissue engineering. J. Physiol. Pharm. 2009, 60 (Suppl. 7), 167–175. [Google Scholar]
- Ryu, J.S.; Ko, K.; Lee, J.W.; Park, S.B.; Byun, S.J.; Jeong, E.J.; Ko, K.; Choo, Y.K. Gangliosides are involved in neural differentiation of human dental pulp-derived stem cells. Biochem. Biophys. Res. Commun. 2009, 387, 266–271. [Google Scholar] [CrossRef]
- Govindasamy, V.; Ronald, V.S.; Abdullah, A.N.; Nathan, K.R.; Ab Aziz, Z.A.; Abdullah, M.; Musa, S.; Kasim, N.H.; Bhonde, R.R. Differentiation of dental pulp stem cells into islet-like aggregates. J. Dent. Res. 2011, 90, 646–652. [Google Scholar] [CrossRef]
- Kanafi, M.M.; Rajeshwari, Y.B.; Gupta, S.; Dadheech, N.; Nair, P.D.; Gupta, P.K.; Bhonde, R.R. Transplantation of islet-like cell clusters derived from human dental pulp stem cells restores normoglycemia in diabetic mice. Cytotherapy 2013, 15, 1228–1236. [Google Scholar] [CrossRef]
- Vollner, F.; Ernst, W.; Driemel, O.; Morsczeck, C. A two-step strategy for neuronal differentiation in vitro of human dental follicle cells. Differentiation 2009, 77, 433–441. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Sun, Z.; Wang, X.; Yang, H.; Shi, S.; Wang, S. Stem cells from human-exfoliated deciduous teeth can differentiate into dopaminergic neuron-like cells. Stem Cells Dev. 2010, 19, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, G.; Yoo, J.J.; De Coppi, P.; Siddiqui, M.M.; Schuch, G.; Pohl, H.G.; Fuhr, J.; Perin, L.; Soker, S.; Atala, A. Propagation, expansion, and multilineage differentiation of human somatic stem cells from dermal progenitors. Stem Cells Dev. 2005, 14, 337–348. [Google Scholar] [CrossRef]
- Riekstina, U.; Muceniece, R.; Cakstina, I.; Muiznieks, I.; Ancans, J. Characterization of human skin-derived mesenchymal stem cell proliferation rate in different growth conditions. Cytotechnology 2008, 58, 153–162. [Google Scholar] [CrossRef]
- Raynaud, C.M.; Maleki, M.; Lis, R.; Ahmed, B.; Al-Azwani, I.; Malek, J.; Safadi, F.F.; Rafii, A. Comprehensive characterization of mesenchymal stem cells from human placenta and fetal membrane and their response to osteoactivin stimulation. Stem Cells Int. 2012, 2012, 658356. [Google Scholar] [CrossRef]
- Rotter, N.; Oder, J.; Schlenke, P.; Lindner, U.; Bohrnsen, F.; Kramer, J.; Rohwedel, J.; Huss, R.; Brandau, S.; Wollenberg, B.; et al. Isolation and characterization of adult stem cells from human salivary glands. Stem Cells Dev. 2008, 17, 509–518. [Google Scholar] [CrossRef]
- Sato, A.; Okumura, K.; Matsumoto, S.; Hattori, K.; Hattori, S.; Shinohara, M.; Endo, F. Isolation, tissue localization, and cellular characterization of progenitors derived from adult human salivary glands. Cloning Stem Cells 2007, 9, 191–205. [Google Scholar] [CrossRef]
- Morito, T.; Muneta, T.; Hara, K.; Ju, Y.J.; Mochizuki, T.; Makino, H.; Umezawa, A.; Sekiya, I. Synovial fluid-derived mesenchymal stem cells increase after intra-articular ligament injury in humans. Rheumatology (Oxf.) 2008, 47, 1137–1143. [Google Scholar] [CrossRef]
- Hatakeyama, A.; Uchida, S.; Utsunomiya, H.; Tsukamoto, M.; Nakashima, H.; Nakamura, E.; Pascual-Garrido, C.; Sekiya, I.; Sakai, A. Isolation and characterization of synovial mesenchymal stem cell derived from hip joints: A comparative analysis with a matched control knee group. Stem Cells Int. 2017, 2017, 9312329. [Google Scholar] [CrossRef]
- Ryu, J.S.; Jung, Y.H.; Cho, M.Y.; Yeo, J.E.; Choi, Y.J.; Kim, Y.I.; Koh, Y.G. Co-culture with human synovium-derived mesenchymal stem cells inhibits inflammatory activity and increases cell proliferation of sodium nitroprusside-stimulated chondrocytes. Biochem. Biophys. Res. Commun. 2014, 447, 715–720. [Google Scholar] [CrossRef]
- Sakaguchi, Y.; Sekiya, I.; Yagishita, K.; Muneta, T. Comparison of human stem cells derived from various mesenchymal tissues: Superiority of synovium as a cell source. Arthritis Rheum. 2005, 52, 2521–2529. [Google Scholar] [CrossRef]
- Schuring, A.N.; Schulte, N.; Kelsch, R.; Ropke, A.; Kiesel, L.; Gotte, M. Characterization of endometrial mesenchymal stem-like cells obtained by endometrial biopsy during routine diagnostics. Fertil. Steril. 2011, 95, 423–426. [Google Scholar] [CrossRef]
- Cheng, Y.; Li, L.; Wang, D.; Guo, Q.; He, Y.; Liang, T.; Sun, L.; Wang, X.; Cheng, Y.; Zhang, G. Characteristics of human endometrium-derived mesenchymal stem cells and their tropism to endometriosis. Stem Cells Int. 2017, 2017, 4794827. [Google Scholar] [CrossRef]
- Jiao, F.; Wang, J.; Dong, Z.L.; Wu, M.J.; Zhao, T.B.; Li, D.D.; Wang, X. Human mesenchymal stem cells derived from limb bud can differentiate into all three embryonic germ layers lineages. Cell. Reprogram. 2012, 14, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Ab Kadir, R.; Zainal Ariffin, S.H.; Megat Abdul Wahab, R.; Kermani, S.; Senafi, S. Characterization of mononucleated human peripheral blood cells. Sci. World J. 2012, 2012, 843843. [Google Scholar] [CrossRef]
- Lin, W.; Xu, L.; Lin, S.; Shi, L.; Wang, B.; Pan, Q.; Lee, W.Y.W.; Li, G. Characterisation of multipotent stem cells from human peripheral blood using an improved protocol. J. Orthop. Transl. 2019, 19, 18–28. [Google Scholar] [CrossRef]
- Ouryazdanpanah, N.; Dabiri, S.; Derakhshani, A.; Vahidi, R.; Farsinejad, A. Peripheral blood-derived mesenchymal stem cells: Growth factor-free isolation, molecular characterization and differentiation. Iran J. Pathol. 2018, 13, 461–466. [Google Scholar] [PubMed]
- Li, S.; Huang, K.J.; Wu, J.C.; Hu, M.S.; Sanyal, M.; Hu, M.; Longaker, M.T.; Lorenz, H.P. Peripheral blood-derived mesenchymal stem cells: Candidate cells responsible for healing critical-sized calvarial bone defects. Stem Cells Transl. Med. 2015, 4, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.S.; Park, J.H.; Kang, J.H.; Kim, S.E.; Park, I.H.; Lee, H.M. Isolation and characterization of multipotent mesenchymal stem cells in nasal polyps. Exp. Biol. Med. (Maywood) 2015, 240, 185–193. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, P.W.; Pezato, R.; Agudelo, J.S.; Perez-Novo, C.A.; Berghe, W.V.; Camara, N.O.; de Almeida, D.C.; Gregorio, L.C. Nasal polyp-derived mesenchymal stromal cells exhibit lack of immune-associated molecules and high levels of stem/progenitor cells markers. Front. Immunol. 2017, 8, 39. [Google Scholar] [CrossRef]
- Shafiee, A.; Kabiri, M.; Ahmadbeigi, N.; Yazdani, S.O.; Mojtahed, M.; Amanpour, S.; Soleimani, M. Nasal septum-derived multipotent progenitors: A potent source for stem cell-based regenerative medicine. Stem Cells Dev. 2011, 20, 2077–2091. [Google Scholar] [CrossRef]
- Koennecke, M.; Boscke, R.; Pfannerstill, A.C.; Reers, S.; Elsner, M.; Fell, B.; Richter, A.; Bruchhage, K.L.; Schumann, S.; Pries, R.; et al. Neuronal differentiation capability of nasal polyps of chronic rhinosinusitis. Arch. Immunol. Exp. (Warsz) 2017, 65, 431–443. [Google Scholar] [CrossRef]
- Okin, D.; Medzhitov, R. Evolution of inflammatory diseases. Curr. Biol. 2012, 22, R733–R740. [Google Scholar] [CrossRef]
- Davidson, A.; Diamond, B. Autoimmune diseases. N. Engl. J. Med. 2001, 345, 340–350. [Google Scholar] [CrossRef]
- Socie, G.; Ritz, J. Current issues in chronic graft-versus-host disease. Blood 2014, 124, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, M.A.; DiPersio, J.F. Mouse models of graft-versus-host disease: Advances and limitations. Dis. Model. Mech. 2011, 4, 318–333. [Google Scholar] [CrossRef]
- Miyashima, S.; Nagata, N.; Nakagawa, T.; Hosaka, N.; Takeuchi, K.; Ogawa, R.; Ikehara, S. Prevention of lpr-graft-versus-host disease and transfer of autoimmune diseases in normal C57BL/6 mice by transplantation of bone marrow cells plus bones (stromal cells) from MRL/lpr mice. J. Immunol. 1996, 156, 79–84. [Google Scholar]
- Chung, N.G.; Jeong, D.C.; Park, S.J.; Choi, B.O.; Cho, B.; Kim, H.K.; Chun, C.S.; Won, J.H.; Han, C.W. Cotransplantation of marrow stromal cells may prevent lethal graft-versus-host disease in major histocompatibility complex mismatched murine hematopoietic stem cell transplantation. Int. J. Hematol. 2004, 80, 370–376. [Google Scholar] [CrossRef]
- Joo, S.Y.; Cho, K.A.; Jung, Y.J.; Kim, H.S.; Park, S.Y.; Choi, Y.B.; Hong, K.M.; Woo, S.Y.; Seoh, J.Y.; Cho, S.J.; et al. Mesenchymal stromal cells inhibit graft-versus-host disease of mice in a dose-dependent manner. Cytotherapy 2010, 12, 361–370. [Google Scholar] [CrossRef]
- Van Belle, T.L.; Coppieters, K.T.; von Herrath, M.G. Type 1 diabetes: Etiology, immunology, and therapeutic strategies. Physiol. Rev. 2011, 91, 79–118. [Google Scholar] [CrossRef] [PubMed]
- Ezquer, F.E.; Ezquer, M.E.; Parrau, D.B.; Carpio, D.; Yanez, A.J.; Conget, P.A. Systemic administration of multipotent mesenchymal stromal cells reverts hyperglycemia and prevents nephropathy in type 1 diabetic mice. Biol. Blood Marrow Transpl. 2008, 14, 631–640. [Google Scholar] [CrossRef]
- Ding, Y.; Xu, D.; Feng, G.; Bushell, A.; Muschel, R.J.; Wood, K.J. Mesenchymal stem cells prevent the rejection of fully allogenic islet grafts by the immunosuppressive activity of matrix metalloproteinase-2 and -9. Diabetes 2009, 58, 1797–1806. [Google Scholar] [CrossRef]
- Berman, D.M.; Willman, M.A.; Han, D.; Kleiner, G.; Kenyon, N.M.; Cabrera, O.; Karl, J.A.; Wiseman, R.W.; O’Connor, D.H.; Bartholomew, A.M.; et al. Mesenchymal stem cells enhance allogeneic islet engraftment in nonhuman primates. Diabetes 2010, 59, 2558–2568. [Google Scholar] [CrossRef]
- Madec, A.M.; Mallone, R.; Afonso, G.; Abou Mrad, E.; Mesnier, A.; Eljaafari, A.; Thivolet, C. Mesenchymal stem cells protect NOD mice from diabetes by inducing regulatory T cells. Diabetologia 2009, 52, 1391–1399. [Google Scholar] [CrossRef]
- Boumaza, I.; Srinivasan, S.; Witt, W.T.; Feghali-Bostwick, C.; Dai, Y.; Garcia-Ocana, A.; Feili-Hariri, M. Autologous bone marrow-derived rat mesenchymal stem cells promote PDX-1 and insulin expression in the islets, alter T cell cytokine pattern and preserve regulatory T cells in the periphery and induce sustained normoglycemia. J. Autoimmun. 2009, 32, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.H.; Seo, M.J.; Reger, R.L.; Spees, J.L.; Pulin, A.A.; Olson, S.D.; Prockop, D.J. Multipotent stromal cells from human marrow home to and promote repair of pancreatic islets and renal glomeruli in diabetic NOD/scid mice. Proc. Natl. Acad. Sci. USA 2006, 103, 17438–17443. [Google Scholar] [CrossRef]
- Solari, M.G.; Srinivasan, S.; Boumaza, I.; Unadkat, J.; Harb, G.; Garcia-Ocana, A.; Feili-Hariri, M. Marginal mass islet transplantation with autologous mesenchymal stem cells promotes long-term islet allograft survival and sustained normoglycemia. J. Autoimmun. 2009, 32, 116–124. [Google Scholar] [CrossRef]
- Sato, M.; Uchida, K.; Nakajima, H.; Miyazaki, T.; Guerrero, A.R.; Watanabe, S.; Roberts, S.; Baba, H. Direct transplantation of mesenchymal stem cells into the knee joints of Hartley strain guinea pigs with spontaneous osteoarthritis. Arthritis Res. Ther. 2012, 14, 1–9. [Google Scholar] [CrossRef]
- Toghraie, F.; Razmkhah, M.; Gholipour, M.A.; Faghih, Z.; Chenari, N.; Torabi Nezhad, S.; Nazhvani Dehghani, S.; Ghaderi, A. Scaffold-free adipose-derived stem cells (ASCs) improve experimentally induced osteoarthritis in rabbits. Arch. Iran. Med. 2012, 15, 495–499. [Google Scholar]
- Soler, R.; Orozco, L.; Munar, A.; Huguet, M.; Lopez, R.; Vives, J.; Coll, R.; Codinach, M.; Garcia-Lopez, J. Final results of a phase I-II trial using ex vivo expanded autologous Mesenchymal Stromal Cells for the treatment of osteoarthritis of the knee confirming safety and suggesting cartilage regeneration. Knee 2016, 23, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Lee, S.M.; Kim, S.H.; Tatman, P.; Gee, A.O.; Kim, D.H.; Lee, K.E.; Jung, Y.; Kim, S.J. Effect of self-assembled peptide-mesenchymal stem cell complex on the progression of osteoarthritis in a rat model. Int. J. Nanomed. 2014, 9 (Suppl. 1), 141–157. [Google Scholar] [CrossRef]
- Li, M.; Luo, X.; Lv, X.; Liu, V.; Zhao, G.; Zhang, X.; Cao, W.; Wang, R.; Wang, W. In vivo human adipose-derived mesenchymal stem cell tracking after intra-articular delivery in a rat osteoarthritis model. Stem Cell Res. Ther. 2016, 7, 160. [Google Scholar] [CrossRef]
- Rao, P.; Segal, B.M. Experimental autoimmune encephalomyelitis. Methods Mol. Biol. 2012, 900, 363–380. [Google Scholar]
- Constantinescu, C.S.; Farooqi, N.; O’Brien, K.; Gran, B. Experimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS). Br. J. Pharm. 2011, 164, 1079–1106. [Google Scholar] [CrossRef]
- Zappia, E.; Casazza, S.; Pedemonte, E.; Benvenuto, F.; Bonanni, I.; Gerdoni, E.; Giunti, D.; Ceravolo, A.; Cazzanti, F.; Frassoni, F.; et al. Mesenchymal stem cells ameliorate experimental autoimmune encephalomyelitis inducing T-cell anergy. Blood 2005, 106, 1755–1761. [Google Scholar] [CrossRef]
- Gerdoni, E.; Gallo, B.; Casazza, S.; Musio, S.; Bonanni, I.; Pedemonte, E.; Mantegazza, R.; Frassoni, F.; Mancardi, G.; Pedotti, R.; et al. Mesenchymal stem cells effectively modulate pathogenic immune response in experimental autoimmune encephalomyelitis. Ann. Neurol. 2007, 61, 219–227. [Google Scholar] [CrossRef]
- Holmdahl, R.; Malmstrom, V.; Burkhardt, H. Autoimmune priming, tissue attack and chronic inflammation—The three stages of rheumatoid arthritis. Eur. J. Immunol. 2014, 44, 1593–1599. [Google Scholar] [CrossRef]
- Abbasi, M.; Mousavi, M.J.; Jamalzehi, S.; Alimohammadi, R.; Bezvan, M.H.; Mohammadi, H.; Aslani, S. Strategies toward rheumatoid arthritis therapy; the old and the new. J. Cell. Physiol. 2019, 234, 10018–10031. [Google Scholar] [CrossRef]
- Brand, D.D.; Latham, K.A.; Rosloniec, E.F. Collagen-induced arthritis. Nat. Protoc. 2007, 2, 1269–1275. [Google Scholar] [CrossRef]
- Augello, A.; Tasso, R.; Negrini, S.M.; Cancedda, R.; Pennesi, G. Cell therapy using allogeneic bone marrow mesenchymal stem cells prevents tissue damage in collagen-induced arthritis. Arthritis Rheum. 2007, 56, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Santalla, M.; Mancheno-Corvo, P.; Menta, R.; Lopez-Belmonte, J.; DelaRosa, O.; Bueren, J.A.; Dalemans, W.; Lombardo, E.; Garin, M.I. Human adipose-derived mesenchymal stem cells modulate experimental autoimmune arthritis by modifying early adaptive T cell responses. Stem Cells 2015, 33, 3493–3503. [Google Scholar] [CrossRef]
- Liu, Y.; Mu, R.; Wang, S.; Long, L.; Liu, X.; Li, R.; Sun, J.; Guo, J.; Zhang, X.; Guo, J.; et al. Therapeutic potential of human umbilical cord mesenchymal stem cells in the treatment of rheumatoid arthritis. Arthritis Res. Ther. 2010, 12, R210. [Google Scholar] [CrossRef] [PubMed]
- Kehoe, O.; Cartwright, A.; Askari, A.; El Haj, A.J.; Middleton, J. Intra-articular injection of mesenchymal stem cells leads to reduced inflammation and cartilage damage in murine antigen-induced arthritis. J. Transl. Med. 2014, 12, 157. [Google Scholar] [CrossRef]
- Gonzalez-Rey, E.; Gonzalez, M.A.; Varela, N.; O’Valle, F.; Hernandez-Cortes, P.; Rico, L.; Buscher, D.; Delgado, M. Human adipose-derived mesenchymal stem cells reduce inflammatory and T cell responses and induce regulatory T cells in vitro in rheumatoid arthritis. Ann. Rheum. Dis. 2010, 69, 241–248. [Google Scholar] [CrossRef]
- Liu, R.; Li, X.; Zhang, Z.; Zhou, M.; Sun, Y.; Su, D.; Feng, X.; Gao, X.; Shi, S.; Chen, W.; et al. Allogeneic mesenchymal stem cells inhibited T follicular helper cell generation in rheumatoid arthritis. Sci. Rep. 2015, 5, 12777. [Google Scholar] [CrossRef]
- Grgic, M.V.; Cupic, H.; Kalogjera, L.; Baudoin, T. Surgical treatment for nasal polyposis: Predictors of outcome. Eur. Arch. Otorhinolaryngol. 2015, 272, 3735–3743. [Google Scholar] [CrossRef]
- Sreeparvathi, A.; Kalyanikuttyamma, L.K.; Kumar, M.; Sreekumar, N.; Veerasigamani, N. Significance of blood eosinophil count in patients with chronic rhinosinusitis with nasal polyposis. J. Clin. Diagn. Res. 2017, 11, MC08–MC11. [Google Scholar] [CrossRef]
- Kim, D.Y.; Lee, S.H.; Carter, R.G.; Kato, A.; Schleimer, R.P.; Cho, S.H. A recently established murine model of nasal polyps demonstrates activation of B cells, as occurs in human nasal polyps. Am. J. Respir. Cell Mol. Biol. 2016, 55, 170–175. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, H.; Xi, Z.; Huang, J.; Nie, J.; Zhou, B.; Deng, Y.; Tao, Z. Establishment of a mouse model of lipopolysaccharide-induced neutrophilic nasal polyps. Exp. Ther. Med. 2017, 14, 5275–5282. [Google Scholar] [CrossRef]
- Takabayashi, T.; Schleimer, R.P. Formation of nasal polyps: The roles of innate type 2 inflammation and deposition of fibrin. J. Allergy Clin. Immunol. 2020, 145, 740–750. [Google Scholar] [CrossRef]
- Polchert, D.; Sobinsky, J.; Douglas, G.; Kidd, M.; Moadsiri, A.; Reina, E.; Genrich, K.; Mehrotra, S.; Setty, S.; Smith, B.; et al. IFN-gamma activation of mesenchymal stem cells for treatment and prevention of graft versus host disease. Eur. J. Immunol. 2008, 38, 1745–1755. [Google Scholar] [CrossRef]
- Tobin, L.M.; Healy, M.E.; English, K.; Mahon, B.P. Human mesenchymal stem cells suppress donor CD4(+) T cell proliferation and reduce pathology in a humanized mouse model of acute graft-versus-host disease. Clin. Exp. Immunol. 2013, 172, 333–348. [Google Scholar] [CrossRef]
- Nylander, A.; Hafler, D.A. Multiple sclerosis. J. Clin. Investig. 2012, 122, 1180–1188. [Google Scholar] [CrossRef]
- Rafei, M.; Birman, E.; Forner, K.; Galipeau, J. Allogeneic mesenchymal stem cells for treatment of experimental autoimmune encephalomyelitis. Mol. Ther. 2009, 17, 1799–1803. [Google Scholar] [CrossRef]
- Bai, L.; Lennon, D.P.; Eaton, V.; Maier, K.; Caplan, A.I.; Miller, S.D.; Miller, R.H. Human bone marrow-derived mesenchymal stem cells induce Th2-polarized immune response and promote endogenous repair in animal models of multiple sclerosis. Glia 2009, 57, 1192–1203. [Google Scholar] [CrossRef]
- Gordon, D.; Pavlovska, G.; Glover, C.P.; Uney, J.B.; Wraith, D.; Scolding, N.J. Human mesenchymal stem cells abrogate experimental allergic encephalomyelitis after intraperitoneal injection, and with sparse CNS infiltration. Neurosci. Lett. 2008, 448, 71–73. [Google Scholar] [CrossRef]
- Rafei, M.; Campeau, P.M.; Aguilar-Mahecha, A.; Buchanan, M.; Williams, P.; Birman, E.; Yuan, S.; Young, Y.K.; Boivin, M.N.; Forner, K.; et al. Mesenchymal stromal cells ameliorate experimental autoimmune encephalomyelitis by inhibiting CD4 Th17 T cells in a CC chemokine ligand 2-dependent manner. J. Immunol. 2009, 182, 5994–6002. [Google Scholar] [CrossRef] [PubMed]
- Rafei, M.; Hsieh, J.; Fortier, S.; Li, M.; Yuan, S.; Birman, E.; Forner, K.; Boivin, M.N.; Doody, K.; Tremblay, M.; et al. Mesenchymal stromal cell-derived CCL2 suppresses plasma cell immunoglobulin production via STAT3 inactivation and PAX5 induction. Blood 2008, 112, 4991–4998. [Google Scholar] [CrossRef]
- Fiorina, P.; Jurewicz, M.; Augello, A.; Vergani, A.; Dada, S.; La Rosa, S.; Selig, M.; Godwin, J.; Law, K.; Placidi, C.; et al. Immunomodulatory function of bone marrow-derived mesenchymal stem cells in experimental autoimmune type 1 diabetes. J. Immunol. 2009, 183, 993–1004. [Google Scholar] [CrossRef]
- Bassi, E.J.; Moraes-Vieira, P.M.; Moreira-Sa, C.S.; Almeida, D.C.; Vieira, L.M.; Cunha, C.S.; Hiyane, M.I.; Basso, A.S.; Pacheco-Silva, A.; Camara, N.O. Immune regulatory properties of allogeneic adipose-derived mesenchymal stem cells in the treatment of experimental autoimmune diabetes. Diabetes 2012, 61, 2534–2545. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B. Articular cartilage degradation in osteoarthritis. HSS J. 2012, 8, 7–9. [Google Scholar] [CrossRef]
- Harrell, C.R.; Markovic, B.S.; Fellabaum, C.; Arsenijevic, A.; Volarevic, V. Mesenchymal stem cell-based therapy of osteoarthritis: Current knowledge and future perspectives. Biomed. Pharm. 2019, 109, 2318–2326. [Google Scholar] [CrossRef]
- Benito, M.J.; Veale, D.J.; FitzGerald, O.; van den Berg, W.B.; Bresnihan, B. Synovial tissue inflammation in early and late osteoarthritis. Ann. Rheum. Dis. 2005, 64, 1263–1267. [Google Scholar] [CrossRef]
- Volarevic, V.; Al-Qahtani, A.; Arsenijevic, N.; Pajovic, S.; Lukic, M.L. Interleukin-1 receptor antagonist (IL-1Ra) and IL-1Ra producing mesenchymal stem cells as modulators of diabetogenesis. Autoimmunity 2010, 43, 255–263. [Google Scholar] [CrossRef]
- Horie, M.; Choi, H.; Lee, R.H.; Reger, R.L.; Ylostalo, J.; Muneta, T.; Sekiya, I.; Prockop, D.J. Intra-articular injection of human mesenchymal stem cells (MSCs) promote rat meniscal regeneration by being activated to express Indian hedgehog that enhances expression of type II collagen. Osteoarthr. Cartil. 2012, 20, 1197–1207. [Google Scholar] [CrossRef]
- Mokbel, A.N.; El Tookhy, O.S.; Shamaa, A.A.; Rashed, L.A.; Sabry, D.; El Sayed, A.M. Homing and reparative effect of intra-articular injection of autologus mesenchymal stem cells in osteoarthritic animal model. BMC Musculoskelet. Disord. 2011, 12, 259. [Google Scholar] [CrossRef] [PubMed]
- Uccelli, A.; de Rosbo, N.K. The immunomodulatory function of mesenchymal stem cells: Mode of action and pathways. Ann. N. Y. Acad. Sci. 2015, 1351, 114–126. [Google Scholar] [CrossRef]
- Cornish, A.L.; Campbell, I.K.; McKenzie, B.S.; Chatfield, S.; Wicks, I.P. G-CSF and GM-CSF as therapeutic targets in rheumatoid arthritis. Nat. Rev. Rheumatol. 2009, 5, 554–559. [Google Scholar] [CrossRef]
- Gonzalez, M.A.; Gonzalez-Rey, E.; Rico, L.; Buscher, D.; Delgado, M. Treatment of experimental arthritis by inducing immune tolerance with human adipose-derived mesenchymal stem cells. Arthritis Rheum. 2009, 60, 1006–1019. [Google Scholar] [CrossRef]
- Mao, F.; Xu, W.R.; Qian, H.; Zhu, W.; Yan, Y.M.; Shao, Q.X.; Xu, H.X. Immunosuppressive effects of mesenchymal stem cells in collagen-induced mouse arthritis. Inflamm. Res. 2010, 59, 219–225. [Google Scholar] [CrossRef]
- Low, D.; Nguyen, D.D.; Mizoguchi, E. Animal models of ulcerative colitis and their application in drug research. Drug Des. Devel. Ther. 2013, 7, 1341–1357. [Google Scholar]
- Gonzalez-Rey, E.; Anderson, P.; Gonzalez, M.A.; Rico, L.; Buscher, D.; Delgado, M. Human adult stem cells derived from adipose tissue protect against experimental colitis and sepsis. Gut 2009, 58, 929–939. [Google Scholar] [CrossRef]
- Gonzalez, M.A.; Gonzalez-Rey, E.; Rico, L.; Buscher, D.; Delgado, M. Adipose-derived mesenchymal stem cells alleviate experimental colitis by inhibiting inflammatory and autoimmune responses. Gastroenterology 2009, 136, 978–989. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, S.; Liu, Y.; Uyanne, J.; Shi, Y.; Shi, S.; Le, A.D. Mesenchymal stem cells derived from human gingiva are capable of immunomodulatory functions and ameliorate inflammation-related tissue destruction in experimental colitis. J. Immunol. 2009, 183, 7787–7798. [Google Scholar] [CrossRef]
- Kim, H.S.; Shin, T.H.; Lee, B.C.; Yu, K.R.; Seo, Y.; Lee, S.; Seo, M.S.; Hong, I.S.; Choi, S.W.; Seo, K.W.; et al. Human umbilical cord blood mesenchymal stem cells reduce colitis in mice by activating NOD2 signaling to COX2. Gastroenterology 2013, 145, 1392–1403. [Google Scholar] [CrossRef]
- Tanaka, F.; Tominaga, K.; Ochi, M.; Tanigawa, T.; Watanabe, T.; Fujiwara, Y.; Ohta, K.; Oshitani, N.; Higuchi, K.; Arakawa, T. Exogenous administration of mesenchymal stem cells ameliorates dextran sulfate sodium-induced colitis via anti-inflammatory action in damaged tissue in rats. Life Sci. 2008, 83, 771–779. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, L.; Jin, Y.; Shi, S. Fas ligand regulates the immunomodulatory properties of dental pulp stem cells. J. Dent. Res. 2012, 91, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, K.; Chen, C.; Wang, D.; Xu, X.; Qu, C.; Yamaza, T.; Cai, T.; Chen, W.; Sun, L.; Shi, S. Mesenchymal-stem-cell-induced immunoregulation involves FAS-ligand-/FAS-mediated T cell apoptosis. Cell Stem Cell 2012, 10, 544–555. [Google Scholar] [CrossRef]
- Tsokos, G.C. Systemic lupus erythematosus. N. Engl. J. Med. 2011, 365, 2110–2121. [Google Scholar] [CrossRef]
- Peng, S.L. Experimental use of mouse models of systemic lupus erythematosus. Methods Mol. Biol. 2012, 900, 135–168. [Google Scholar]
- Chang, J.W.; Hung, S.P.; Wu, H.H.; Wu, W.M.; Yang, A.H.; Tsai, H.L.; Yang, L.Y.; Lee, O.K. Therapeutic effects of umbilical cord blood-derived mesenchymal stem cell transplantation in experimental lupus nephritis. Cell Transpl. 2011, 20, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Akiyama, K.; Zhang, H.; Yamaza, T.; Hou, Y.; Zhao, S.; Xu, T.; Le, A.; Shi, S. Mesenchymal stem cell transplantation reverses multiorgan dysfunction in systemic lupus erythematosus mice and humans. Stem Cells 2009, 27, 1421–1432. [Google Scholar] [CrossRef]
- Zhou, K.; Zhang, H.; Jin, O.; Feng, X.; Yao, G.; Hou, Y.; Sun, L. Transplantation of human bone marrow mesenchymal stem cell ameliorates the autoimmune pathogenesis in MRL/lpr mice. Cell. Mol. Immunol. 2008, 5, 417–424. [Google Scholar] [CrossRef]
- Gu, Z.; Akiyama, K.; Ma, X.; Zhang, H.; Feng, X.; Yao, G.; Hou, Y.; Lu, L.; Gilkeson, G.S.; Silver, R.M.; et al. Transplantation of umbilical cord mesenchymal stem cells alleviates lupus nephritis in MRL/lpr mice. Lupus 2010, 19, 1502–1514. [Google Scholar] [CrossRef] [PubMed]
- Pawankar, R. Nasal polyposis: An update: Editorial review. Curr. Opin. Allergy Clin. Immunol. 2003, 3, 1–6. [Google Scholar] [CrossRef]
- Fokkens, W.J.; Lund, V.J.; Mullol, J.; Bachert, C.; Alobid, I.; Baroody, F.; Cohen, N.; Cervin, A.; Douglas, R.; Gevaert, P.; et al. EPOS 2012: European position paper on rhinosinusitis and nasal polyps 2012. A summary for otorhinolaryngologists. Rhinology 2012, 50, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Meltzer, E.O.; Hamilos, D.L.; Hadley, J.A.; Lanza, D.C.; Marple, B.F.; Nicklas, R.A.; Bachert, C.; Baraniuk, J.; Baroody, F.M.; Benninger, M.S.; et al. Rhinosinusitis: Establishing definitions for clinical research and patient care. J. Allergy Clin. Immunol. 2004, 114, 155–212. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.P.; Li, H.B.; Wang, B.F.; Wang, S.B.; You, X.J.; Cui, Y.H.; Wang, D.Y.; Desrosiers, M.; Liu, Z. Distinct immunopathologic characteristics of various types of chronic rhinosinusitis in adult Chinese. J. Allergy Clin. Immunol. 2009, 124, 478–484. [Google Scholar] [CrossRef]
- Van Crombruggen, K.; Zhang, N.; Gevaert, P.; Tomassen, P.; Bachert, C. Pathogenesis of chronic rhinosinusitis: Inflammation. J. Allergy Clin. Immunol. 2011, 128, 728–732. [Google Scholar] [CrossRef]
- Prota, L.F.; Lassance, R.M.; Maron-Gutierrez, T.; Castiglione, R.C.; Garcia, C.S.; Santana, M.C.; Souza-Menezes, J.; Abreu, S.C.; Samoto, V.; Santiago, M.F.; et al. Bone marrow mononuclear cell therapy led to alveolar-capillary membrane repair, improving lung mechanics in endotoxin-induced acute lung injury. Cell Transpl. 2010, 19, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.G.; He, L.; Shi, X.M.; Wu, S.S.; Zhang, B.; Mei, L.; Xu, Y.J.; Zhang, Z.X.; Zhao, J.P.; Zhang, H.L. Regulation of transplanted mesenchymal stem cells by the lung progenitor niche in rats with chronic obstructive pulmonary disease. Respir. Res. 2014, 15, 33. [Google Scholar] [CrossRef]
- Meng, F.; Meliton, A.; Moldobaeva, N.; Mutlu, G.; Kawasaki, Y.; Akiyama, T.; Birukova, A.A. Asef mediates HGF protective effects against LPS-induced lung injury and endothelial barrier dysfunction. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 308, L452–L463. [Google Scholar] [CrossRef]
- Katsha, A.M.; Ohkouchi, S.; Xin, H.; Kanehira, M.; Sun, R.; Nukiwa, T.; Saijo, Y. Paracrine factors of multipotent stromal cells ameliorate lung injury in an elastase-induced emphysema model. Mol. Ther. 2011, 19, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Fang, X.; Gupta, N.; Serikov, V.; Matthay, M.A. Allogeneic human mesenchymal stem cells for treatment of E. coli endotoxin-induced acute lung injury in the ex vivo perfused human lung. Proc. Natl. Acad. Sci. USA 2009, 106, 16357–16362. [Google Scholar] [CrossRef]
- Thebaud, B.; Ladha, F.; Michelakis, E.D.; Sawicka, M.; Thurston, G.; Eaton, F.; Hashimoto, K.; Harry, G.; Haromy, A.; Korbutt, G.; et al. Vascular endothelial growth factor gene therapy increases survival, promotes lung angiogenesis, and prevents alveolar damage in hyperoxia-induced lung injury: Evidence that angiogenesis participates in alveolarization. Circulation 2005, 112, 2477–2486. [Google Scholar] [CrossRef]
- Mei, S.H.; McCarter, S.D.; Deng, Y.; Parker, C.H.; Liles, W.C.; Stewart, D.J. Prevention of LPS-induced acute lung injury in mice by mesenchymal stem cells overexpressing angiopoietin 1. PLoS Med. 2007, 4, e269. [Google Scholar] [CrossRef]
- McCarter, S.D.; Mei, S.H.; Lai, P.F.; Zhang, Q.W.; Parker, C.H.; Suen, R.S.; Hood, R.D.; Zhao, Y.D.; Deng, Y.; Han, R.N.; et al. Cell-based angiopoietin-1 gene therapy for acute lung injury. Am. J. Respir. Crit. Care Med. 2007, 175, 1014–1026. [Google Scholar] [CrossRef]
- Qin, T.; Liu, C.J.; Zhang, H.W.; Pan, Y.F.; Tang, Q.; Liu, J.K.; Wang, Y.Z.; Hu, M.X.; Xue, F. Effect of the IkBalpha mutant gene delivery to mesenchymal stem cells on rat chronic pancreatitis. Genet. Mol. Res. 2014, 13, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.H.; Li, M.L.; Qin, A.L.; Lv, S.X.; Wen, T.; Zhu, X.Y.; Li, L.Y.; Dong, Y.; Hu, C.Y.; Hu, D.M.; et al. Reduction of fibrosis in dibutyltin dichloride-induced chronic pancreatitis using rat umbilical mesenchymal stem cells from Wharton’s Jelly. Pancreas 2013, 42, 1291–1302. [Google Scholar] [CrossRef]
- Nicoletti, A.; Michel, J.B. Cardiac fibrosis and inflammation: Interaction with hemodynamic and hormonal factors. Cardiovasc. Res. 1999, 41, 532–543. [Google Scholar] [CrossRef]
- Hatzistergos, K.E.; Quevedo, H.; Oskouei, B.N.; Hu, Q.; Feigenbaum, G.S.; Margitich, I.S.; Mazhari, R.; Boyle, A.J.; Zambrano, J.P.; Rodriguez, J.E.; et al. Bone marrow mesenchymal stem cells stimulate cardiac stem cell proliferation and differentiation. Circ. Res. 2010, 107, 913–922. [Google Scholar] [CrossRef]
- Hatzistergos, K.E.; Saur, D.; Seidler, B.; Balkan, W.; Breton, M.; Valasaki, K.; Takeuchi, L.M.; Landin, A.M.; Khan, A.; Hare, J.M. Stimulatory effects of mesenchymal stem cells on cKit+ cardiac stem cells are mediated by SDF1/CXCR4 and SCF/cKit signaling pathways. Circ. Res. 2016, 119, 921–930. [Google Scholar] [CrossRef]
- Quevedo, H.C.; Hatzistergos, K.E.; Oskouei, B.N.; Feigenbaum, G.S.; Rodriguez, J.E.; Valdes, D.; Pattany, P.M.; Zambrano, J.P.; Hu, Q.; McNiece, I.; et al. Allogeneic mesenchymal stem cells restore cardiac function in chronic ischemic cardiomyopathy via trilineage differentiating capacity. Proc. Natl. Acad. Sci. USA 2009, 106, 14022–14027. [Google Scholar] [CrossRef] [PubMed]
- Asmussen, S.; Ito, H.; Traber, D.L.; Lee, J.W.; Cox, R.A.; Hawkins, H.K.; McAuley, D.F.; McKenna, D.H.; Traber, L.D.; Zhuo, H.; et al. Human mesenchymal stem cells reduce the severity of acute lung injury in a sheep model of bacterial pneumonia. Thorax 2014, 69, 819–825. [Google Scholar] [CrossRef]
- Feizpour, A.; Boskabady, M.H.; Ghorbani, A. Adipose-derived stromal cell therapy affects lung inflammation and tracheal responsiveness in guinea pig model of COPD. PLoS ONE 2014, 9, e108974. [Google Scholar] [CrossRef]
- Loi, R.; Beckett, T.; Goncz, K.K.; Suratt, B.T.; Weiss, D.J. Limited restoration of cystic fibrosis lung epithelium in vivo with adult bone marrow-derived cells. Am. J. Respir. Crit. Care Med. 2006, 173, 171–179. [Google Scholar] [CrossRef]
- Tanaka, K.; Fujita, T.; Umezawa, H.; Namiki, K.; Yoshioka, K.; Hagihara, M.; Sudo, T.; Kimura, S.; Tatsumi, K.; Kasuya, Y. Therapeutic effect of lung mixed culture-derived epithelial cells on lung fibrosis. Lab. Investig. 2014, 94, 1247–1259. [Google Scholar] [CrossRef]
- Aslam, M.; Baveja, R.; Liang, O.D.; Fernandez-Gonzalez, A.; Lee, C.; Mitsialis, S.A.; Kourembanas, S. Bone marrow stromal cells attenuate lung injury in a murine model of neonatal chronic lung disease. Am. J. Respir. Crit. Care Med. 2009, 180, 1122–1130. [Google Scholar] [CrossRef]
- Alphonse, R.S.; Vadivel, A.; Fung, M.; Shelley, W.C.; Critser, P.J.; Ionescu, L.; O’Reilly, M.; Ohls, R.K.; McConaghy, S.; Eaton, F.; et al. Existence, functional impairment, and lung repair potential of endothelial colony-forming cells in oxygen-induced arrested alveolar growth. Circulation 2014, 129, 2144–2157. [Google Scholar] [CrossRef] [PubMed]
- Van Haaften, T.; Byrne, R.; Bonnet, S.; Rochefort, G.Y.; Akabutu, J.; Bouchentouf, M.; Rey-Parra, G.J.; Galipeau, J.; Haromy, A.; Eaton, F.; et al. Airway delivery of mesenchymal stem cells prevents arrested alveolar growth in neonatal lung injury in rats. Am. J. Respir. Crit. Care Med. 2009, 180, 1131–1142. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, H.; Shi, Y.; Peng, W.; Zhang, S.; Zhang, W.; Xu, J.; Mei, Y.; Feng, Z. Role of bone marrow-derived mesenchymal stem cells in the prevention of hyperoxia-induced lung injury in newborn mice. Cell Biol. Int. 2012, 36, 589–594. [Google Scholar] [CrossRef]
- Ionescu, L.I.; Alphonse, R.S.; Arizmendi, N.; Morgan, B.; Abel, M.; Eaton, F.; Duszyk, M.; Vliagoftis, H.; Aprahamian, T.R.; Walsh, K.; et al. Airway delivery of soluble factors from plastic-adherent bone marrow cells prevents murine asthma. Am. J. Respir. Cell Mol. Biol. 2012, 46, 207–216. [Google Scholar] [CrossRef]
- De Paepe, M.E.; Mao, Q.; Ghanta, S.; Hovanesian, V.; Padbury, J.F. Alveolar epithelial cell therapy with human cord blood-derived hematopoietic progenitor cells. Am. J. Pathol. 2011, 178, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.O.; Kim, M.Y.; Cho, M.Y.; Baik, S.K.; Cho, Y.Z.; Kwon, S.O. Effect of bone marrow-derived mesenchymal stem cells on hepatic fibrosis in a thioacetamide-induced cirrhotic rat model. BMC Gastroenterol. 2014, 14, 198. [Google Scholar] [CrossRef] [PubMed]
- Schubert, T.; Xhema, D.; Veriter, S.; Schubert, M.; Behets, C.; Delloye, C.; Gianello, P.; Dufrane, D. The enhanced performance of bone allografts using osteogenic-differentiated adipose-derived mesenchymal stem cells. Biomaterials 2011, 32, 8880–8891. [Google Scholar] [CrossRef]
- Baksh, D.; Yao, R.; Tuan, R.S. Comparison of proliferative and multilineage differentiation potential of human mesenchymal stem cells derived from umbilical cord and bone marrow. Stem Cells 2007, 25, 1384–1392. [Google Scholar] [CrossRef]
- Mohammadi Gorji, S.; Karimpor Malekshah, A.A.; Hashemi-Soteh, M.B.; Rafiei, A.; Parivar, K.; Aghdami, N. Effect of mesenchymal stem cells on Doxorubicin-induced fibrosis. Cell J. 2012, 14, 142–151. [Google Scholar]
- Eom, Y.W.; Shim, K.Y.; Baik, S.K. Mesenchymal stem cell therapy for liver fibrosis. Korean J. Intern. Med. 2015, 30, 580–589. [Google Scholar] [CrossRef]
- Zhang, Z.; Lin, H.; Shi, M.; Xu, R.; Fu, J.; Lv, J.; Chen, L.; Lv, S.; Li, Y.; Yu, S.; et al. Human umbilical cord mesenchymal stem cells improve liver function and ascites in decompensated liver cirrhosis patients. J. Gastroenterol. Hepatol. 2012, 27 (Suppl. 2), 112–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, J.; Liu, H.; Li, Y.; Fu, J.; Sun, Y.; Xu, R.; Lin, H.; Wang, S.; Lv, S.; et al. Pilot study of umbilical cord-derived mesenchymal stem cell transfusion in patients with primary biliary cirrhosis. J. Gastroenterol. Hepatol. 2013, 28 (Suppl. 1), 85–92. [Google Scholar] [CrossRef]
- El-Ansary, M.; Abdel-Aziz, I.; Mogawer, S.; Abdel-Hamid, S.; Hammam, O.; Teaema, S.; Wahdan, M. Phase II trial: Undifferentiated versus differentiated autologous mesenchymal stem cells transplantation in Egyptian patients with HCV induced liver cirrhosis. Stem Cell Rev. Rep. 2012, 8, 972–981. [Google Scholar] [CrossRef] [PubMed]
- Salama, H.; Zekri, A.R.; Medhat, E.; Al Alim, S.A.; Ahmed, O.S.; Bahnassy, A.A.; Lotfy, M.M.; Ahmed, R.; Musa, S. Peripheral vein infusion of autologous mesenchymal stem cells in Egyptian HCV-positive patients with end-stage liver disease. Stem Cell Res. Ther. 2014, 5, 70. [Google Scholar] [CrossRef]
- Xu, L.; Gong, Y.; Wang, B.; Shi, K.; Hou, Y.; Wang, L.; Lin, Z.; Han, Y.; Lu, L.; Chen, D.; et al. Randomized trial of autologous bone marrow mesenchymal stem cells transplantation for hepatitis B virus cirrhosis: Regulation of Treg/Th17 cells. J. Gastroenterol. Hepatol. 2014, 29, 1620–1628. [Google Scholar] [CrossRef]
- Kawakubo, K.; Ohnishi, S.; Fujita, H.; Kuwatani, M.; Onishi, R.; Masamune, A.; Takeda, H.; Sakamoto, N. Effect of fetal membrane-derived mesenchymal stem cell transplantation in rats with acute and chronic pancreatitis. Pancreas 2016, 45, 707–713. [Google Scholar] [CrossRef]
- Cao, Y.; Sun, Z.; Liao, L.; Meng, Y.; Han, Q.; Zhao, R.C. Human adipose tissue-derived stem cells differentiate into endothelial cells in vitro and improve postnatal neovascularization in vivo. Biochem. Biophys. Res. Commun. 2005, 332, 370–379. [Google Scholar] [CrossRef]
- Premer, C.; Blum, A.; Bellio, M.A.; Schulman, I.H.; Hurwitz, B.E.; Parker, M.; Dermarkarian, C.R.; DiFede, D.L.; Balkan, W.; Khan, A.; et al. Allogeneic mesenchymal stem cells restore endothelial function in heart failure by stimulating endothelial progenitor cells. EBioMedicine 2015, 2, 467–475. [Google Scholar] [CrossRef]
- Beigi, F.; Schmeckpeper, J.; Pow-Anpongkul, P.; Payne, J.A.; Zhang, L.; Zhang, Z.; Huang, J.; Mirotsou, M.; Dzau, V.J. C3orf58, a novel paracrine protein, stimulates cardiomyocyte cell-cycle progression through the PI3K-AKT-CDK7 pathway. Circ. Res. 2013, 113, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Mirotsou, M.; Zhang, Z.; Deb, A.; Zhang, L.; Gnecchi, M.; Noiseux, N.; Mu, H.; Pachori, A.; Dzau, V. Secreted frizzled related protein 2 (Sfrp2) is the key Akt-mesenchymal stem cell-released paracrine factor mediating myocardial survival and repair. Proc. Natl. Acad. Sci. USA 2007, 104, 1643–1648. [Google Scholar] [CrossRef]
- Suzuki, G.; Iyer, V.; Lee, T.C.; Canty, J.M., Jr. Autologous mesenchymal stem cells mobilize cKit+ and CD133+ bone marrow progenitor cells and improve regional function in hibernating myocardium. Circ. Res. 2011, 109, 1044–1054. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.S.; Miller, L.; Patel, A.N.; Anderson, R.D.; Mendelsohn, F.O.; Traverse, J.; Silver, K.H.; Shin, J.; Ewald, G.; Farr, M.J.; et al. Changes in ventricular remodelling and clinical status during the year following a single administration of stromal cell-derived factor-1 non-viral gene therapy in chronic ischaemic heart failure patients: The STOP-HF randomized Phase II trial. Eur. Heart J. 2015, 36, 2228–2238. [Google Scholar] [CrossRef] [PubMed]
- Mias, C.; Lairez, O.; Trouche, E.; Roncalli, J.; Calise, D.; Seguelas, M.H.; Ordener, C.; Piercecchi-Marti, M.D.; Auge, N.; Salvayre, A.N.; et al. Mesenchymal stem cells promote matrix metalloproteinase secretion by cardiac fibroblasts and reduce cardiac ventricular fibrosis after myocardial infarction. Stem Cells 2009, 27, 2734–2743. [Google Scholar] [CrossRef] [PubMed]
- De Lisio, M.; Jensen, T.; Sukiennik, R.A.; Huntsman, H.D.; Boppart, M.D. Substrate and strain alter the muscle-derived mesenchymal stem cell secretome to promote myogenesis. Stem Cell Res. Ther. 2014, 5, 74. [Google Scholar] [CrossRef]
- Alexakis, C.; Partridge, T.; Bou-Gharios, G. Implication of the satellite cell in dystrophic muscle fibrosis: A self-perpetuating mechanism of collagen overproduction. Am. J. Physiol. Cell Physiol. 2007, 293, C661–C669. [Google Scholar] [CrossRef]
- Ibrahim, A.G.; Cheng, K.; Marban, E. Exosomes as critical agents of cardiac regeneration triggered by cell therapy. Stem Cell Rep. 2014, 2, 606–619. [Google Scholar] [CrossRef]
- Barile, L.; Lionetti, V.; Cervio, E.; Matteucci, M.; Gherghiceanu, M.; Popescu, L.M.; Torre, T.; Siclari, F.; Moccetti, T.; Vassalli, G. Extracellular vesicles from human cardiac progenitor cells inhibit cardiomyocyte apoptosis and improve cardiac function after myocardial infarction. Cardiovasc. Res. 2014, 103, 530–541. [Google Scholar] [CrossRef]
- Gray, W.D.; French, K.M.; Ghosh-Choudhary, S.; Maxwell, J.T.; Brown, M.E.; Platt, M.O.; Searles, C.D.; Davis, M.E. Identification of therapeutic covariant microRNA clusters in hypoxia-treated cardiac progenitor cell exosomes using systems biology. Circ. Res. 2015, 116, 255–263. [Google Scholar] [CrossRef]
- Rao, K.S.; Aronshtam, A.; McElory-Yaggy, K.L.; Bakondi, B.; VanBuren, P.; Sobel, B.E.; Spees, J.L. Human epicardial cell-conditioned medium contains HGF/IgG complexes that phosphorylate RYK and protect against vascular injury. Cardiovasc. Res. 2015, 107, 277–286. [Google Scholar] [CrossRef][Green Version]
- Wei, K.; Serpooshan, V.; Hurtado, C.; Diez-Cunado, M.; Zhao, M.; Maruyama, S.; Zhu, W.; Fajardo, G.; Noseda, M.; Nakamura, K.; et al. Epicardial FSTL1 reconstitution regenerates the adult mammalian heart. Nature 2015, 525, 479–485. [Google Scholar] [CrossRef]
- Chen, L.; Wang, Y.; Pan, Y.; Zhang, L.; Shen, C.; Qin, G.; Ashraf, M.; Weintraub, N.; Ma, G.; Tang, Y. Cardiac progenitor-derived exosomes protect ischemic myocardium from acute ischemia/reperfusion injury. Biochem. Biophys. Res. Commun. 2013, 431, 566–571. [Google Scholar] [CrossRef]
- Nery, A.A.; Nascimento, I.C.; Glaser, T.; Bassaneze, V.; Krieger, J.E.; Ulrich, H. Human mesenchymal stem cells: From immunophenotyping by flow cytometry to clinical applications. Cytom. A 2013, 83, 48–61. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Source | Cell Surface Markers | Lineage Differentiation | References | |
|---|---|---|---|---|
| Positive | Negative | |||
| Bone marrow | SH2, SH3, CD29, CD44, CD49e, CD71, CD73, CD90, CD105, CD106, CD166, CD120a, CD124, STRO-1 | CD14, CD34, CD45, CD19, CD3, CD31, CD11b, HLA-DR | Adipocytes, Chondrocytes, Osteoblasts, Hepatocyte, Cardiomyocytes, Pancreatic cells, Neuronal-like cells | [18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34] |
| Umbilical cord, Umbilical cord blood | CK8, CK18, CK19, CD10, CD13, CD29, CD44, CD73, CD90, CD105, CD106, HLA-I, HLA-II | CD14, CD31, CD33, CD34, CD45, CD38, CD79, CD133, vWF, HLA-DR | Adipocytes, Chondrocytes, Osteoblasts, Hepatocytes, Endothelial-like cells, Neuronal-like cells, Pancreatic cells | [6,24,35,36,37,38,39,40,41,42,43,44] |
| Wharton’s jelly | CD13, CD29, CD44, CD73, CD90, CD105, HLA-I | CD14, CD34, CD45, CD31, CD79, HLA-II, HLA-DR | Adipocytes, Osteoblasts, Chondrocytes, Hepatocytes, Neuronal-like cells | [55,56,57,58,59,60,61,62] |
| Adipose tissue | CD13, CD29, CD44, CD71, CD73, CD90, CD105, CD166, HLA-I, HLA-ABC, STRO-1 | CD10, CD14, CD24, CD31, CD34, CD36, CD38, CD45, CD49, CD117, CD133, SSEA4, CD106, HLA-II, HLA-DR | Adipocytes, Chondrocytes, Osteoblasts, Hepatocyte, Cardiomyocytes, Pancreatic cells, Neuronal-like cells, | [24,26,45,46,47,48,49,50,51,52,53,54] |
| Amniotic fluid | SH2, SH3, SH4, CD, CD29, CD44, CD49, CD54, CD58, CD71, CD73, CD90, CD105, CD123, CD166, HLA-ABC, HLA-DR | CD10, CD11, CD14, CD31, CD34, CD49, CD50, CD117, HLA-DR, DP, DQ, EMA | Adipocytes, Osteoblasts, Neuronal-like cells | [63,64,65] |
| Dental tissues | CD29, CD44, CD90, CD105, SH2, SH3, CDHLA-DR, CD117, CD46, DPSC-EZ, DPSC-OG | CD10, CD14, CD34, CD45, HLA-DR, Stro-1, NGFR | Adipocytes, Chondrocytes, Osteoblasts, Pancreatic cells, Melanocytes, Neuronal-like cells | [12,13,66,67,68,69,70,71,72,73,74] |
| Skin and foreskin | CD44, CD90, CD73, CD105, CD166, SSEA4, Vimentin | CD14, CD45, CD34, c-kit, CD133, SSEA3, OCT-4, TRA 1–60, TRA 1–81, HLA-DR | Adipocytes, Osteoblasts, Chondrocytes, Myoblasts | [75,76] |
| Placenta | CD29, CD44, CD73, CD90, CD105 | CD45, CD34, HLA-DR | Adipocytes, Osteoblasts, Endothelial-like cells, Neuronal-like cells | [36,77] |
| Salivary gland | CD13, CD29, CD44, CD49f, Thy-1, CD90, CD104, p75NGFR, β2-microglobulin, CD130, STRO-1 | CD34, CD38, CD45, CD133 | Adipocytes, Chondrocytes, Osteoblasts, Pancreatic endocrines | [78,79] |
| Synovial fluid | CD10, CD166, CD44, CD54, CD90, CD105, CD147, D7-FIB, STRO-1 | CD31, CD34, CD45, CD106, CD117, CD166, VEGFR2, Flk-1, CXCR4, BMPR-1A, NGFR | Adipocytes, Chondrocytes, Osteoblasts | [80,81] |
| Synovial tissues | CD4, CD34, CD45 | CD44, CD73, CD90, CD105 | Adipocytes, Chondrocytes, Osteoblasts | [10,11,82,83] |
| Nasal polyp tissues | CD105, CD90, CD73, CD54, CD44 | CD34, CD45, CD117, HLA-DR, PDL-1, PDL-2, CTLA-4, CD106, CD146, CD31 | Adipocytes, Osteoblasts, Chondrocytes, Neuronal-like cells | [91,92,93,94] |
| Endometrium | CD73, CD90, CD105, CD146 | CD34, CD45 | Adipocytes, Chondrocytes, Osteoblasts | [84,85] |
| Limb bud | CD13, CD29, CD90, CD105, CD106 | CD3, CD4, CD14, CD15, CD34, CD45, HLA-DR | Osteoblasts, Adipocytes, Hepatocytes, Neuronal-like cells | [86] |
| Peripheral blood | CD44, CD90, CD105, HLA-ABC, CD29, CD73, CD90.1, CD106, CD140α | CD45, CD133, CD34, CD19, CD11b, c-kit | Adipocytes, Osteoblasts, Chondrocytes, Neuronal-like cells | [87,88,89,90] |
| Disease Model (Generation Methods) | Up-Regulation | Down-Regulation | References |
|---|---|---|---|
| Graft-vs-host disease; Depleting endogenous hematopoietic cells by radiation or chemotherapy | Regulatory T cells | Auto-antibodies Inflammatory cytokines T cell proliferation TH1 cells | [97,98,99,100,101] |
| Type 1 diabetes (T1D); Treatment of streptozotocin | Regulatory T cells Tissue repair TH2 cells | Inflammatory T cells TH1 cells | [102,103,104,105,106] |
| Pancreatic islet transplantation Treatment of streptozotocin | Islet survival Regulatory T cells | TH1 cytokines T cell responsiveness | [107,108,109] |
| Experimental autoimmune arthritis For rheumatoid arthritis, collagen-induced arthritis For osteoarthritis, meniscectomy; ovariectomy; treatment of sodium monoiodoacetate | Regulatory T cells IL-10 TH2 cells | Inflammatory cytokines T cell responsiveness | [110,111,112,113,114] |
| Experimental autoimmune encephalomyelitis (EAE); Induced CNS inflammation by treatment of complete Freund’s adjuvant | TH2 | T cell responsiveness CNS infiltration Auto-antibodies TH1/TH17 cells | [115,116,117,118] |
| Inflammatory bowel disease (IBD); 1, Treatment with dextran sulfate sodium added to drinking water 2, Intrarectal administration of trinitrobenzene sulfonic acid | Anti-inflammatory cytokines Regulatory T cells FasL-mediated T cell apoptosis | Inflammatory T cells Inflammatory cytokines Intestinal CD4+ T cell infiltration Growth factor expression T cell responsiveness | [111,119,120,121,122,123] |
| Systemic lupus erythematosus (SLE); 1, Progeny of a breeding pair consisting of a New Zealand Black mouse and New Zealand White mouse. 2, Mutation in the gene encoding Fas on the MRL strain background | Regulatory T cells Anti-inflammatory cytokines | Anti-DNA antibodies T cell frequency TH17 cells Plasma cells Inflammatory cytokines | [124,125,126,127] |
| Chronic rhinosinusitis with nasal polyps (CRSwNP); For eosinophilic CRSwNP, Ovalbumin and Staphylococcus aureus enterotoxin B For non-eosinophilic CRSwNP, Lipopolysaccharide | Regulatory T cells IL-10 | CD4+ and CD8+ T cell proliferation IL-2, TNF-α, IFN-γ | [49,128,129,130,131,132] |
| Disease Model | Route of Delivery | Therapeutic Effect | References |
|---|---|---|---|
| Lung | |||
| Bronchopulmoary dysplasia | |||
| Hyperoxia neonatal lung injury | Intravenous, Intratracheal, intraperitoneal | Protection of alveoli, Reduce and decrease inflammation, pulmonary injury, hypertension and fibrosis Vascular growth, Increase survival | [166,167,168,169] |
| Acute respiratory distress syndrome | |||
| Bacterial pneumonia | Intravenous | Improve oxygenation (PaO2/FiO2w) Decrease pulmonary edema | [162] |
| LPS-induced inflammation | Intravenous | Reduce histopathological changes, Increased survival, Protection of alveoli, Lung mechanics improve | [170] |
| Chronic lower respiratory disease | |||
| Cigarette smoke exposure | Intratracheal /Intravenous | Decrease tracheal responsiveness, inflammatory cytokines, and inflammatory cell infiltration | [163] |
| LPS, cigarette smoke, and 17% oxygen exposure | Intratracheal | Decrease in inflammatory cytokines, Increase in ECM production | [171] |
| Cycstic fibrosis | |||
| Naphthalene-induced lung injury | Intravenous | Little to no level of CFTR dependent chloride secretion | [164] |
| Idiopathic pulmonary fibrosis | |||
| Bleomycin-induced lung injury | Intratracheal | Decrease fibrosis and airway inflammation | [165] |
| Liver | |||
| Chronic hepatitis B | Intravenous | Improvement of liver function and MELD score Reduce ascites | [172] |
| Primary biliary cirrhosis | Intravenous | Decrease in serum ALP and γ-GGT | [173] |
| Hepatitis C virus cirrhosis | Intravenous infusion, Peripheral vein | Improvement in liver function; Frequency of encephalopathy, jaundice, ascites, bleeding tendency, and lower limb edema | [174,175] |
| Hepatitis B virus cirrhosis | Hepatic artery | Improvement in liver function | [176] |
| Pancreas | |||
| Dibutyltin dichloride | Penile vein, Jugular vein | Immunomodulatory effect Inhibition of activation of pancreatic satellite cells Anti-apoptotic effect | [177,178,179] |
| Heart | |||
| Ischemic heart failure | Intramyocardial | Reduction of infarct scar, inflammation, vascular permeability, fibrosis in scarred tissues Improve LVEF and endothelial function Increase cardiac function, survival and angiogenesis | [180,181,182,183,184,185] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryu, J.-S.; Jeong, E.-J.; Kim, J.-Y.; Park, S.J.; Ju, W.S.; Kim, C.-H.; Kim, J.-S.; Choo, Y.-K. Application of Mesenchymal Stem Cells in Inflammatory and Fibrotic Diseases. Int. J. Mol. Sci. 2020, 21, 8366. https://doi.org/10.3390/ijms21218366
Ryu J-S, Jeong E-J, Kim J-Y, Park SJ, Ju WS, Kim C-H, Kim J-S, Choo Y-K. Application of Mesenchymal Stem Cells in Inflammatory and Fibrotic Diseases. International Journal of Molecular Sciences. 2020; 21(21):8366. https://doi.org/10.3390/ijms21218366
Chicago/Turabian StyleRyu, Jae-Sung, Eun-Jeong Jeong, Jong-Yeup Kim, Soon Ju Park, Won Seok Ju, Chang-Hyun Kim, Jang-Seong Kim, and Young-Kug Choo. 2020. "Application of Mesenchymal Stem Cells in Inflammatory and Fibrotic Diseases" International Journal of Molecular Sciences 21, no. 21: 8366. https://doi.org/10.3390/ijms21218366
APA StyleRyu, J.-S., Jeong, E.-J., Kim, J.-Y., Park, S. J., Ju, W. S., Kim, C.-H., Kim, J.-S., & Choo, Y.-K. (2020). Application of Mesenchymal Stem Cells in Inflammatory and Fibrotic Diseases. International Journal of Molecular Sciences, 21(21), 8366. https://doi.org/10.3390/ijms21218366