E3 Ubiquitin Ligase APC/CCdh1 Regulation of Phenylalanine Hydroxylase Stability and Function

 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. PAH Is Degraded by the 26S Proteasomal Pathway
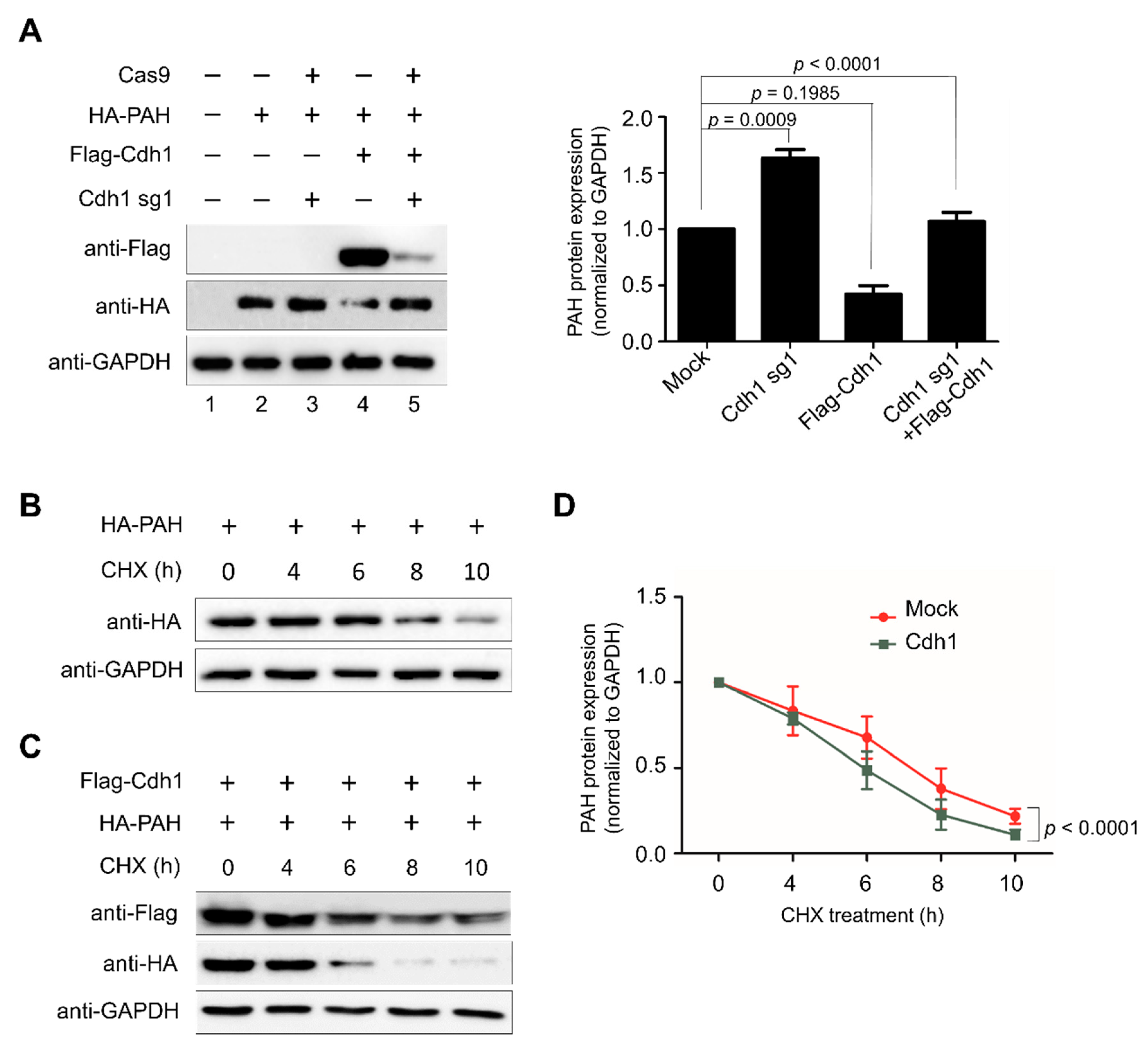
2.2. APC/CCdh1 Is the E3 Ligase Responsible for PAH Protein Degradation
2.3. APC/CCdh1 Is a Negative Regulator of PAH Protein Stability
2.4. APC/CCdh1 Interacts and Co-Localizes with PAH
2.5. APC/CCdh1 Declines PAH Protein Half-Life
2.6. APC/CCdh1 Promotes PAH Polyubiquitination
2.7. Depletion of APC/CCdh1 Promotes Phenylalanine Metabolism
2.8. Association of APC/CCdh1 and PAH in HCC
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plasmids
5.2. Antibodies and Reagents
5.3. Cas9 and sgRNA Constructs
5.4. Cell Culture and Transfections
5.5. T7 Endonuclease I (T7E1) Assay
5.6. Immunoprecipitation and Immunoblotting
5.7. Immunofluorescence Microscopy
5.8. PAH Activity Assay
5.9. Expression and Survival Analysis Based on TCGA Data
5.10. Immunohistochemistry
5.11. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AAAHs | aromatic amino acid hydroxylases; |
| PAH | phenylalanine hydroxylase; |
| TH | tyrosine hydroxylase; |
| TPH1 | tryptophan hydroxylase 1; |
| TPH2 | tryptophan hydroxylase 2; |
| l-Phe | l-phenylalanine; |
| l-Tyr | l-tyrosine; |
| PKU | phenylketonuria; |
| UPS | ubiquitin-proteasome system; |
| ERAD | endoplasmic reticulum-associated degradation; |
| APC/C | anaphase-promoting complex/cyclosome; |
| BH4 | tetrahydrobiopterin; |
| Cdh1 | cell division cycle 20 related protein 1; |
| CDC20 | cell division cycle 20; |
| sgRNA | single-guide RNA; |
| T7E1 | T7 endonuclease I; |
| HEK293 | human embryonic kidney 293; |
| CHX | cycloheximide; |
| PEI | polyethyleneimine |
References
- Arturo, E.C.; Gupta, K.; Héroux, A.; Stith, L.; Cross, P.J.; Parker, E.J.; Loll, P.J.; Jaffe, E.K. First structure of full-length mammalian phenylalanine hydroxylase reveals the architecture of an autoinhibited tetramer. Proc. Natl. Acad. Sci. USA 2016, 113, 2394–2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arturo, E.C.; Gupta, K.; Hansen, M.R.; Borne, E.; Jaffe, E.K. Biophysical characterization of full-length human phenylalanine hydroxylase provides a deeper understanding of its quaternary structure equilibrium. J. Biol. Chem. 2019, 294, 10131–10145. [Google Scholar] [CrossRef] [PubMed]
- Erlandsen, H.; Stevens, R.C. The structural basis of phenylketonuria. Mol. Genet. Metab. 1999, 68, 103–125. [Google Scholar] [CrossRef] [PubMed]
- Heintz, C.; Troxler, H.; Martinez, A.; Thöny, B.; Blau, N. Quantification of phenylalanine hydroxylase activity by isotope-dilution liquid chromatography-electrospray ionization tandem mass spectrometry. Mol. Genet. Metab. 2012, 105, 559–565. [Google Scholar] [CrossRef] [Green Version]
- Tourian, A.; Goddard, J.; Puck, T.T. Phenylalanine hydroxylase activity in mammalian cells. J. Cell. Physiol. 1969, 73, 159–170. [Google Scholar] [CrossRef]
- van Wegberg, A.M.J.; MacDonald, A.; Ahring, K.; Bélanger-Quintana, A.; Blau, N.; Bosch, A.M.; Burlina, A.; Campistol, J.; Feillet, F.; Giżewska, M.; et al. The complete European guidelines on phenylketonuria: Diagnosis and treatment. Orphanet J. Rare Dis. 2017, 12, 162. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.A.; Mamotte, C.D.S.; Burnett, J.R. Phenylketonuria: An inborn error of phenylalanine metabolism. Clin. Biochem. Rev. 2008, 29, 31–41. [Google Scholar]
- Centerwall, S.A.; Centerwall, W.R. The discovery of phenylketonuria: The story of a young couple, two retarded children, and a scientist. Pediatrics 2000, 105, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Xu, T.; Liu, C.; Meng, G.; Sun, Y.; Qian, L.; Wang, S.; Wei, J.; Yu, D.; Ding, Y. Liver-enriched Genes are Associated with the Prognosis of Patients with Hepatocellular Carcinoma. Sci. Rep. 2018, 8, 11197. [Google Scholar] [CrossRef]
- Døskeland, A.P.; Flatmark, T. Recombinant human phenylalanine hydroxylase is a substrate for the ubiquitin-conjugating enzyme system. Biochem. J. 1996, 319, 941–945. [Google Scholar] [CrossRef] [Green Version]
- Antao, A.M.; Tyagi, A.; Kim, K.S.; Ramakrishna, S. Advances in Deubiquitinating Enzyme Inhibition and Applications in Cancer Therapeutics. Cancers 2020, 12, 1579. [Google Scholar] [CrossRef] [PubMed]
- Hershko, A.; Ciechanover, A. The ubiquitin system. Annu. Rev. Biochem. 1998, 67, 425–479. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.-M. The anaphase promoting complex/cyclosome: A machine designed to destroy. Nat. Rev. Mol. Cell Biol. 2006, 7, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; Jiang, S.; Zheng, H.; Yin, Q.; Xie, M.; Little, M.R.; Yin, X.; Chen, M.; Song, S.J.; Beg, A.A.; et al. Interplay between c-Src and the APC/C co-activator Cdh1 regulates mammary tumorigenesis. Nat. Commun. 2019, 10, 3716. [Google Scholar] [CrossRef] [PubMed]
- Meghini, F.; Martins, T.; Tait, X.; Fujimitsu, K.; Yamano, H.; Glover, D.M.; Kimata, Y. Targeting of Fzr/Cdh1 for timely activation of the APC/C at the centrosome during mitotic exit. Nat. Commun. 2016, 7, 12607. [Google Scholar] [CrossRef] [Green Version]
- Baker, R.E.; Shiman, R. Measurement of phenylalanine hydroxylase turnover in cultured hepatoma cells. J. Biol. Chem. 1979, 254, 9633–9639. [Google Scholar]
- Chang, N.; Kaufman, S.; Milstien, S. The mechanism of the irreversible inhibition ofrat liver phenylalanine hydroxylase due to treatment with p-chlorophenylalanine. The lack of effect on turnover of phenylalanine hydroxylase. J. Biol. Chem. 1979, 254, 2665–2668. [Google Scholar]
- Dawson, S.P. Hepatocellular carcinoma and the ubiquitin-proteasome system. Biochim. Biophys. Acta 2008, 1782, 775–784. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Wu, J.; Cai, H.; Wang, D.; Yu, L.; Zhang, W.H. E3 Ubiquitin Ligase Siah-1 is Down-regulated and Fails to Target Natural HBx Truncates for Degradation in Hepatocellular Carcinoma. J. Cancer 2016, 7, 418–426. [Google Scholar] [CrossRef] [Green Version]
- Cesari, R.; Martin, E.S.; Calin, G.A.; Pentimalli, F.; Bichi, R.; McAdams, H.; Trapasso, F.; Drusco, A.; Shimizu, M.; Masciullo, V.; et al. Parkin, a gene implicated in autosomal recessive juvenile parkinsonism, is a candidate tumor suppressor gene on chromosome 6q25-q27. Proc. Natl. Acad. Sci. USA 2003, 100, 5956–5961. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, M.; Marusawa, H.; Wang, H.Q.; Iwai, A.; Ikeuchi, K.; Imai, Y.; Kataoka, A.; Nukina, N.; Takahashi, R.; Chiba, T. Parkin as a tumor suppressor gene for hepatocellular carcinoma. Oncogene 2008, 27, 6002–6011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Denison, S.; Lai, J.P.; Philips, L.A.; Montoya, D.; Kock, N.; Schüle, B.; Klein, C.; Shridhar, V.; Roberts, L.R.; et al. Parkin gene alterations in hepatocellular carcinoma. Genes Chromosomes Cancer 2004, 40, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Ishizawa, J.; Sugihara, E.; Kuninaka, S.; Mogushi, K.; Kojima, K.; Benton, C.B.; Zhao, R.; Chachad, D.; Hashimoto, N.; Jacamo, R.O.; et al. FZR1 loss increases sensitivity to DNA damage and consequently promotes murine and human B-cell acute leukemia. Blood 2017, 129, 1958–1968. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, H.; Li, J.; Zhang, J.; Wu, J.; Li, Y.; Piao, Y.; Pan, L.; Xiang, R.; Yue, S. FZR1 as a novel biomarker for breast cancer neoadjuvant chemotherapy prediction. Cell Death Dis. 2020, 11, 804. [Google Scholar] [CrossRef]
- García-Higuera, I.; Manchado, E.; Dubus, P.; Cañamero, M.; Méndez, J.; Moreno, S.; Malumbres, M. Genomic stability and tumour suppression by the APC/C cofactor Cdh1. Nat. Cell Biol. 2008, 10, 802–811. [Google Scholar] [CrossRef]
- Greil, C.; Krohs, J.; Schnerch, D.; Follo, M.; Felthaus, J.; Engelhardt, M.; Wäsch, R. The role of APC/C(Cdh1) in replication stress and origin of genomic instability. Oncogene 2016, 35, 3062–3070. [Google Scholar] [CrossRef]
- Brito, D.A.; Rieder, C.L. Mitotic checkpoint slippage in humans occurs via cyclin B destruction in the presence of an active checkpoint. Curr. Biol. 2006, 16, 1194–1200. [Google Scholar] [CrossRef] [Green Version]
- Flydal, M.I.; Martinez, A. Phenylalanine hydroxylase: Function, structure, and regulation. IUBMB Life 2013, 65, 341–349. [Google Scholar] [CrossRef]
- Lee, Y.W.; Lee, D.H.; Kim, N.D.; Lee, S.T.; Ahn, J.Y.; Choi, T.Y.; Lee, Y.K.; Kim, S.H.; Kim, J.W.; Ki, C.S. Mutation analysis of PAH gene and characterization of a recurrent deletion mutation in Korean patients with phenylketonuria. Exp. Mol. Med. 2008, 40, 533–540. [Google Scholar] [CrossRef]
- Wang, Z.-W.; Jiang, S.-W.; Zhou, B.-C. PAH mutation spectrum and correlation with PKU manifestation in north Jiangsu province population. Kaohsiung J. Med Sci. 2018, 34, 89–94. [Google Scholar] [CrossRef]
- Zschocke, J. Phenylketonuria mutations in Europe. Hum. Mutat. 2003, 21, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, S. The phenylalanine hydroxylating system. Adv. Enzymol. Relat. Areas Mol. Biol. 1993, 67, 77–264. [Google Scholar] [CrossRef] [PubMed]
- Scheller, R.; Stein, A.; Nielsen, S.V.; Marin, F.I.; Gerdes, A.M.; Di Marco, M.; Papaleo, E.; Lindorff-Larsen, K.; Hartmann-Petersen, R. Toward mechanistic models for genotype-phenotype correlations in phenylketonuria using protein stability calculations. Hum. Mutat. 2019, 40, 444–457. [Google Scholar] [CrossRef] [PubMed]
- Gersting, S.W.; Kemter, K.F.; Staudigl, M.; Messing, D.D.; Danecka, M.K.; Lagler, F.B.; Sommerhoff, C.P.; Roscher, A.A.; Muntau, A.C. Loss of function in phenylketonuria is caused by impaired molecular motions and conformational instability. Am. J. Hum. Genet. 2008, 83, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Gallego, D.; Leal, F.; Gámez, A.; Castro, M.; Navarrete, R.; Sanchez-Lijarcio, O.; Vitoria, I.; Bueno-Delgado, M.; Belanger-Quintana, A.; Morais, A.; et al. Pathogenic variants of DNAJC12 and evaluation of the encoded cochaperone as a genetic modifier of hyperphenylalaninemia. Hum. Mutat. 2020, 41, 1329–1338. [Google Scholar] [CrossRef]
- Liu, X.; Yamashita, T.; Shang, J.; Shi, X.; Morihara, R.; Huang, Y.; Sato, K.; Takemoto, M.; Hishikawa, N.; Ohta, Y.; et al. Molecular switching from ubiquitin-proteasome to autophagy pathways in mice stroke model. J. Cereb. Blood Flow Metab. 2020, 40, 214–224. [Google Scholar] [CrossRef]
- Kevei, É.; Pokrzywa, W.; Hoppe, T. Repair or destruction-an intimate liaison between ubiquitin ligases and molecular chaperones in proteostasis. FEBS Lett. 2017, 591, 2616–2635. [Google Scholar] [CrossRef]
- Rahimi, H.; Negahdari, B.; Shokrgozar, M.A.; Madadkar-Sobhani, A.; Mahdian, R.; Foroumadi, A.; Amin, M.K.; Karimipoor, M. A structural model of the anaphase promoting complex co-activator (Cdh1) and in silico design of inhibitory compounds. Res. Pharm. Sci. 2015, 10, 59–67. [Google Scholar]
- Qiao, X.; Zhang, L.; Gamper, A.M.; Fujita, T.; Wan, Y. APC/C-Cdh1: From cell cycle to cellular differentiation and genomic integrity. Cell Cycle 2010, 9, 3904–3912. [Google Scholar] [CrossRef]
- Maestre, C.; Delgado-Esteban, M.; Gomez-Sanchez, J.C.; Bolanos, J.P.; Almeida, A. Cdk5 phosphorylates Cdh1 and modulates cyclin B1 stability in excitotoxicity. EMBO J. 2008, 27, 2736–2745. [Google Scholar] [CrossRef] [Green Version]
- Komander, D.; Rape, M. The ubiquitin code. Annu. Rev. Biochem. 2012, 81, 203–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chau, V.; Tobias, J.W.; Bachmair, A.; Marriott, D.; Ecker, D.J.; Gonda, D.K.; Varshavsky, A. A multiubiquitin chain is confined to specific lysine in a targeted short-lived protein. Science 1989, 243, 1576. [Google Scholar] [CrossRef] [PubMed]
- Yau, R.; Rape, M. The increasing complexity of the ubiquitin code. Nat. Cell Biol. 2016, 18, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Ohtake, F.; Tsuchiya, H.; Saeki, Y.; Tanaka, K. K63 ubiquitylation triggers proteasomal degradation by seeding branched ubiquitin chains. Proc. Natl. Acad. Sci. USA 2018, 115, E1401. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.J.; Sun, L.J. Nonproteolytic Functions of Ubiquitin in Cell Signaling. Mol. Cell 2009, 33, 275–286. [Google Scholar] [CrossRef]
- Senft, D.; Qi, J.; Ronai, Z.A. Ubiquitin ligases in oncogenic transformation and cancer therapy. Nat. Rev. Cancer 2018, 18, 69–88. [Google Scholar] [CrossRef]
- Geng, C.; He, B.; Xu, L.; Barbieri, C.E.; Eedunuri, V.K.; Chew, S.A.; Zimmermann, M.; Bond, R.; Shou, J.; Li, C.; et al. Prostate cancer-associated mutations in speckle-type POZ protein (SPOP) regulate steroid receptor coactivator 3 protein turnover. Proc. Natl. Acad. Sci. USA 2019, 116, 14386–14387. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Ci, W.; Karmakar, S.; Chen, K.; Dhar, R.; Fan, Z.; Guo, Z.; Zhang, J.; Ke, Y.; Wang, L.; et al. SPOP promotes tumorigenesis by acting as a key regulatory hub in kidney cancer. Cancer Cell 2014, 25, 455–468. [Google Scholar] [CrossRef] [Green Version]
- Emanuelli, A.; Manikoth Ayyathan, D.; Koganti, P.; Shah, P.A.; Apel-Sarid, L.; Paolini, B.; Detroja, R.; Frenkel-Morgenstern, M.; Blank, M. Altered Expression and Localization of Tumor Suppressive E3 Ubiquitin Ligase SMURF2 in Human Prostate and Breast Cancer. Cancers 2019, 11, 556. [Google Scholar] [CrossRef] [Green Version]
- Manfredi, J.J. The Mdm2-p53 relationship evolves: Mdm2 swings both ways as an oncogene and a tumor suppressor. Genes Dev. 2010, 24, 1580–1589. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.J.; Wu, H.; Shen, X.Z. The ubiquitin-proteasome system and its potential application in hepatocellular carcinoma therapy. Cancer Lett. 2016, 379, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Ma, X.; Zhao, W.; Huai, W.; Li, T.; Qiu, Y.; Zhang, Y.; Han, L. TRIM31 is upregulated in hepatocellular carcinoma and promotes disease progression by inducing ubiquitination of TSC1-TSC2 complex. Oncogene 2018, 37, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Higashitsuji, H.; Itoh, K.; Nagao, T.; Dawson, S.; Nonoguchi, K.; Kido, T.; Mayer, R.J.; Arii, S.; Fujita, J. Reduced stability of retinoblastoma protein by gankyrin, an oncogenic ankyrin-repeat protein overexpressed in hepatomas. Nat. Med. 2000, 6, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Ally, A.; Balasundaram, M.; Carlsen, R.; Chuah, E.; Clarke, A.; Dhalla, N.; Holt, R.A.; Jones, S.J.M.; Lee, D.; Ma, Y.; et al. Comprehensive and Integrative Genomic Characterization of Hepatocellular Carcinoma. Cell 2017, 169, 1327–1341.e1323. [Google Scholar] [CrossRef]
- Yang, J.D.; Hainaut, P.; Gores, G.J.; Amadou, A.; Plymoth, A.; Roberts, L.R. A global view of hepatocellular carcinoma: Trends, risk, prevention and management. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 589–604. [Google Scholar] [CrossRef]
- Fitzmaurice, C.; Allen, C.; Barber, R.M.; Barregard, L.; Bhutta, Z.A.; Brenner, H.; Dicker, D.J.; Chimed-Orchir, O.; Dandona, R.; Dandona, L.; et al. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life-years for 32 Cancer Groups, 1990 to 2015: A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncol. 2017, 3, 524–548. [Google Scholar] [CrossRef]
- Ramakrishna, S.; Kwaku Dad, A.-B.; Beloor, J.; Gopalappa, R.; Lee, S.-K.; Kim, H. Gene disruption by cell-penetrating peptide-mediated delivery of Cas9 protein and guide RNA. Genome Res. 2014, 24, 1020–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Kim, D.; Cho, S.W.; Kim, J.; Kim, J.S. Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins. Genome Res. 2014, 24, 1012–1019. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Lee, H.J.; Kim, H.; Cho, S.W.; Kim, J.S. Targeted genome editing in human cells with zinc finger nucleases constructed via modular assembly. Genome Res. 2009, 19, 1279–1288. [Google Scholar] [CrossRef] [Green Version]
- Staudigl, M.; Gersting, S.W.; Danecka, M.K.; Messing, D.D.; Woidy, M.; Pinkas, D.; Kemter, K.F.; Blau, N.; Muntau, A.C. The interplay between genotype, metabolic state and cofactor treatment governs phenylalanine hydroxylase function and drug response. Hum. Mol. Genet. 2011, 20, 2628–2641. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Mishra, M.K.; Eltoum, I.A.; Bae, S.; Lillard, J.W., Jr.; Singh, R. CCR5/CCL5 axis interaction promotes migratory and invasiveness of pancreatic cancer cells. Sci. Rep. 2018, 8, 1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyagi, A.; Sarodaya, N.; Kaushal, K.; Chandrasekaran, A.P.; Antao, A.M.; Suresh, B.; Rhie, B.H.; Kim, K.S.; Ramakrishna, S. E3 Ubiquitin Ligase APC/CCdh1 Regulation of Phenylalanine Hydroxylase Stability and Function. Int. J. Mol. Sci. 2020, 21, 9076. https://doi.org/10.3390/ijms21239076
Tyagi A, Sarodaya N, Kaushal K, Chandrasekaran AP, Antao AM, Suresh B, Rhie BH, Kim KS, Ramakrishna S. E3 Ubiquitin Ligase APC/CCdh1 Regulation of Phenylalanine Hydroxylase Stability and Function. International Journal of Molecular Sciences. 2020; 21(23):9076. https://doi.org/10.3390/ijms21239076
Chicago/Turabian StyleTyagi, Apoorvi, Neha Sarodaya, Kamini Kaushal, Arun Pandian Chandrasekaran, Ainsley Mike Antao, Bharathi Suresh, Byung Ho Rhie, Kye Seong Kim, and Suresh Ramakrishna. 2020. "E3 Ubiquitin Ligase APC/CCdh1 Regulation of Phenylalanine Hydroxylase Stability and Function" International Journal of Molecular Sciences 21, no. 23: 9076. https://doi.org/10.3390/ijms21239076
APA StyleTyagi, A., Sarodaya, N., Kaushal, K., Chandrasekaran, A. P., Antao, A. M., Suresh, B., Rhie, B. H., Kim, K. S., & Ramakrishna, S. (2020). E3 Ubiquitin Ligase APC/CCdh1 Regulation of Phenylalanine Hydroxylase Stability and Function. International Journal of Molecular Sciences, 21(23), 9076. https://doi.org/10.3390/ijms21239076